北部湾鱼类碳、氮、磷生态化学计量特征
2023-06-14王才广黄亮亮RadSaeed吴志强
王才广,朱 亮,黄亮亮,2,*,刘 昊,3,胥 鹏,Rad Saeed,吴志强
1 桂林理工大学环境科学与工程学院,桂林 541004
2 广西岩溶地区水污染控制与用水安全保障协同创新中心,桂林 541004
3 广西环境污染控制理论与技术重点实验室,桂林 541004
4 广西北部湾海洋生物多样性养护重点实验室(北部湾大学),钦州 535011
生态化学计量学是研究碳(C)、氮(N)和磷(P)平衡以及元素平衡理论对生态交互作用的影响,它将生物体的生态功能与其元素组成或化学计量比联系起来[1—3]。目前,国内研究主要集中在C、N、P元素循环[4—5]、生物养分限制判断[6]及C、N、P生态化学计量学特征对植物生长的指示性作用[7—10]等方面,有关鱼类生态化学计量学研究偏少。近几年国内研究人员在水生生物如浮游动物、底栖动物及鱼类也有相关研究,但相对较少。如陈蕾[11]、苏强[12]等对浮游动物化学计量学稳态性特征开展研究并进行总结;蔡永久等[13]研究了长江中下游浅水湖泊常见底栖动物碳、氮、磷化学计量特征;Huang等[14]、李培培[15]、孙培英[16]和朱利明等[17]分别对黄海鳀鱼、千岛湖鲢、鳙、长江口中国花鲈和斑尾刺虾虎鱼及三峡水库小江流域鱼类的生态化学计量特征开展研究。另外,王芳等[18]综述了水域生态系统生态化学计量学研究进展。与此同时,国外学者如Saara Mäkelin等[19]、Priscilla Le Mézo等[20]、Jacob E.Allgeier[21]等对不同海域鱼类及无脊椎动物的生态化学计量学开展系列研究,并提出构建全球海洋鱼类及无脊椎动物生态化学计量学数据库倡议。截至目前,我国尚无系统的近岸海域生态系统生态化学计量学研究数据,而全球生物地球化学循环的计量需要世界各海区(包括中国海区)的数据积累,为生态化学计量学理论提供海洋案例的补充。鱼类作为海域生态系统的主要消费者,在生态系统的物质循环中起重要作用,消费者自身的 C、N、P 含量反映其对不同元素的需求比例,消费者和食物的营养比例不平衡会影响生物体新陈代谢、生长、繁殖乃至整个种群的生态过程[22—23]。鱼类生态化学计量学研究大多基于动态平衡理论和生长速率假说[23—24]。动态平衡理论认为面对多变的环境,生物可以保持相对恒定的营养成分[3],超过生物需要的元素含量则被生物过程排出体外,而那些限制供应量的元素被保留[25];生长速率假说认为环境(包括饵料)C∶N∶P 的变化能影响有机体的生长速率,进而影响其种群分布特征。反之,生物为适应生长速率的改变会调整其C∶N∶P比值[26]。迄今为止,鱼类生态化学计量学研究多聚焦阐明元素组成与大小或年龄的关系。此外,食性也可能对生物体的营养成分有重要影响[19,27—31]。
北部湾(105°40′—110°10′ E、17°00′—21°45′ N)作为南海北部重要水域,属于热带和亚热带半封闭海湾,其三面环陆且海底呈扇形向南部陆架海洋敞开,平均水深38 m,最大水深不超过100 m,水深由北向南、自岸向中逐渐加深,沿岸有大型河流红河等注入,区域降水丰沛。湾内地形地貌及底质特征复杂,鱼类资源丰富[32]。北部湾鱼类研究主要集中在经济鱼类生物学特征[33—35]、渔业资源调查及动态分析[36]等方面,缺乏系统的海域生态系统生态化学计量学研究数据。因此,本研究以北部湾海域作为研究区域,系统研究该海域鱼类生态化学计量特征及其影响因素,丰富我国沿海鱼类生态化学计量学数据,为海洋生物地球化学循环及渔业资源的保护和管理提供科学依据。
1 材料与方法
1.1 采样点设置和采样时间
本研究在广西、广东、海南三个省(区)共布置了9个采样站点,其分别为北部湾海域近岸城市(即企沙、北海、江洪、企水、徐闻、海口、白马井、八所和崖州)的大型渔港(图1)。鱼类样品于2021年3—4月在各个渔港购得,经现场分类鉴定后冰冻运往实验室进行后续处理。

图1 北部湾样点分布图Fig.1 Distribution of sampling sites in the Beibu Gulf
1.2 样品处理和元素测定
物种鉴定参考《中国海洋鱼类》[37]和《北部湾鱼类图鉴》[38]。所有鱼类个体解冻,并用去离子水清洗后,测量全长、体长(±1 mm)及体重(±0.1 g),鱼体内脏含有大量富含C的脂质和富含N的蛋白质,且不同物种差异较大[39—40],为了标准化鱼类的潜在偏差,本研究鱼体内脏均被去除[23,41—42]。用研磨机捣碎鱼体,经冷冻干燥后磨碎混匀过筛(60目),待上机测定。称取2.5—4.0 mg干燥样品,采用元素分析仪(型号EA2400Ⅱ)进行C、N测定;称取0.2—0.5 g干燥样品,加入混合酸7 mL(HNO3∶H2O2=5∶2%)微波消解,消解后于加热板(100 ℃)上赶酸30 min,冷却后定容至50 ml,采用电感耦合等离子体发射光谱(型号Opetima 7000DV)进行Ca、P测定,Ca和P的回收率是90%—110%。C、N、P 、Ca含量的结果为干质量百分比(%),C∶N、C∶P、N∶P 比为元素的摩尔比。
1.3 数据分析
运用Excel 2016对数据进行预处理,SPSS 23.0软件进行数据统计分析,OriginPro 2021作图。首先对所有鱼类样品的C、N和P含量及其比值进行基本统计分析,正态分布检验采用K-S检验(Kolmogorov-Smirnov test)并作正态直方图观察其正态分布趋势,不同种类的 C、N、P 含量用单因素方差分析(One-way ANOVA)比较;通过多元方差分析(MANOVA)不同种类元素比值的差异显著性,鉴于C∶N、C∶P、N∶P 相互之间并不独立,当 MANOVA 分析显示有显著性差异时,再用 One-way ANOVA分别对元素比值进行比较。为分析元素含量及比值之间的关系,用Pearson相关分析和一元线性回归分析其相关性及其数据变异的解释量。统计分析前,元素含量和比值分别进行对数(lg)转换。
2 结果与分析
2.1 鱼类不同种类元素组成
本研究共采集鱼类79种382尾,隶属14目39科61属,根据食性分为草食性、滤食性、杂食性和肉食性四大类(附表1)。鱼类元素含量变化较大(表1),C、N、P含量的变化范围分别为33.87%—58.34%、6.31%—14.92%和0.77%—4.67%,变异系数分别为8.71%、13.60%和36.10%,三种元素含量的变异系数由大到小排列均为P >N >C(图2)。P含量变化较大也导致了C∶P和N∶P变幅较大(变异系数分别为40.30%和39.23%),其最大值可达最小值的9倍和6.7倍。相反,C∶N的变幅较小(变异系数为20.19),其最大值仅为最小值的2.8倍。

附表1 北部湾鱼类基本情况Supplementary Table 1 Basic situation of fish in the Beibu Gulf
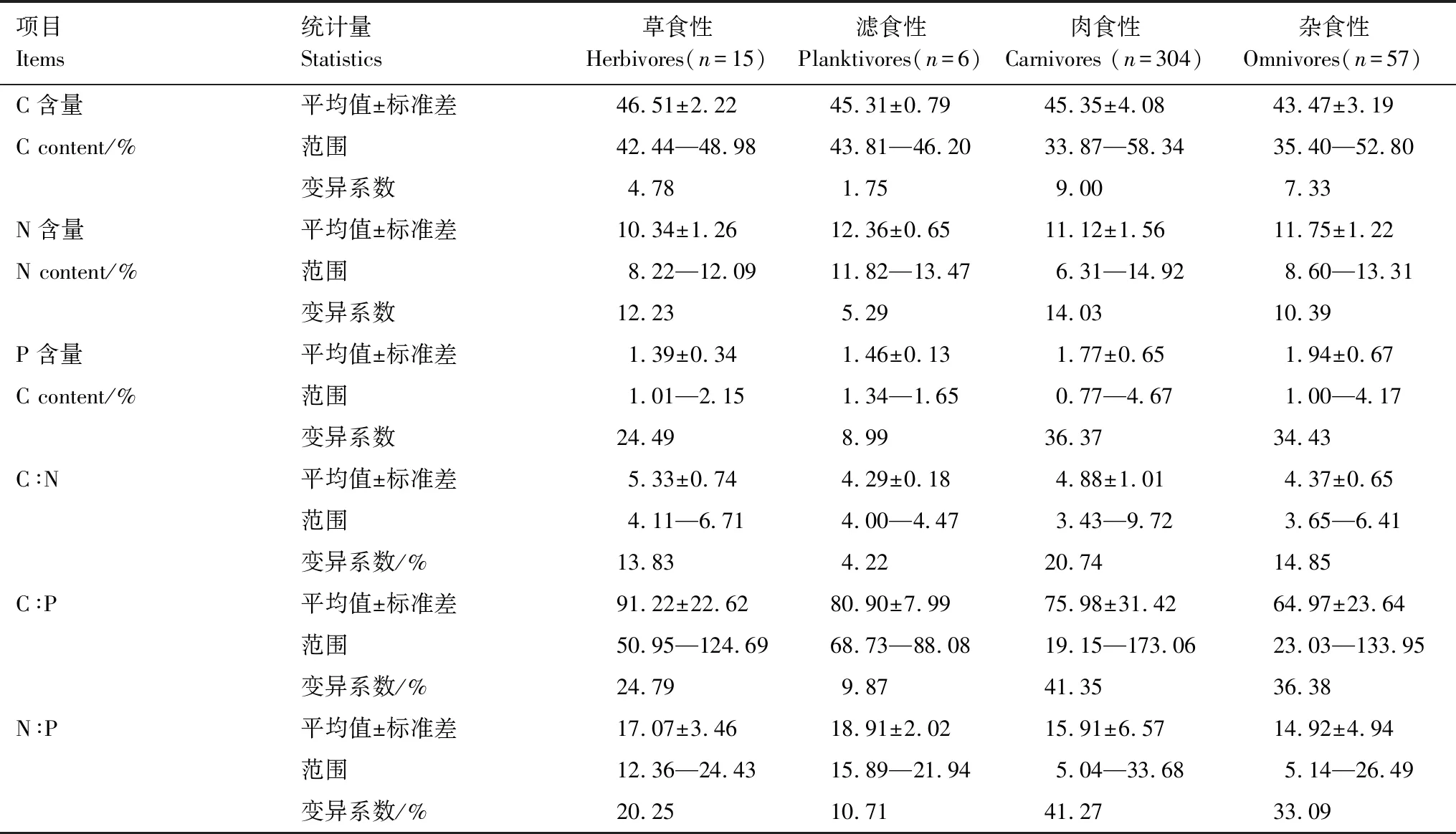
表1 不同食性鱼类碳、氮、磷元素含量和比值Table 1 C,N,P contents and ratios of fishes in different feeding habits
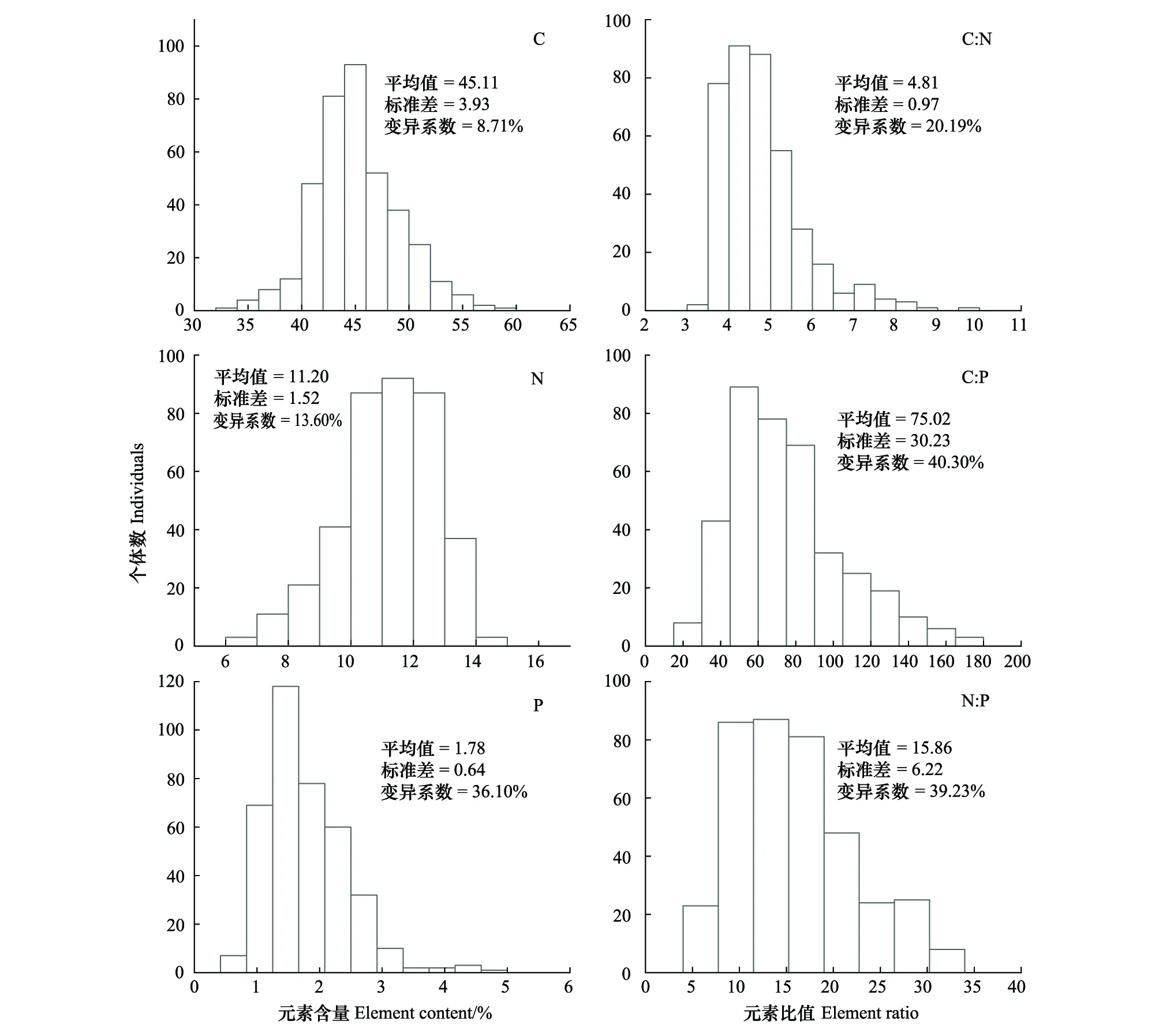
图2 鱼类碳、氮、磷元素含量和比值频数直方图(n=382)Fig.2 Frequency histograms of C,N,P,C∶N,C∶P and N∶P in fishes (n=382)
主成分分析法(PCA)分析了北部湾79种鱼类的C、N、P含量及其比值之间的关系(图3),主成分的解释率为81.1%,物种之间存在显著差异,尤其在科间的差异较大如虾虎鱼科、带鱼科、鲾科、蛇鳗科和石首鱼科。物种在科内的距离较近,差异性较小。
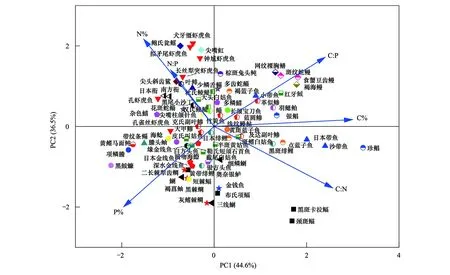
图3 北部湾鱼类与碳、氮、磷元素含量和比值之间的主成分分析(PCA)图Fig.3 Principal Component Analysis (PCA) diagram of the contents and ratios of C,N and P among fishes from the Beibu Gulf
2.2 不同体型鱼类生态化学计量特征
将鱼类体型分为侧扁型、带型、纺锤型、鳗型、平扁型和鲀型6类(附表1),方差分析结果表明不同体型鱼类的C、N、P含量及比值具有显著差异(P<0.05)(图4)。带型鱼类C含量显著高于其他体型鱼类,平扁型鱼类C含量显著低于其他体型鱼类;鲀型和平扁型鱼类N含量显著高于其他鱼类;侧扁型、纺锤型和平扁型鱼类P含量高于带型、鳗型和鲀型鱼类;侧扁型和带型鱼类C∶N最高,纺锤型和鳗型次之,平扁型和鲀型最低;带型、鳗型和鲀型鱼类C∶P显著高于侧扁型、纺锤型和平扁型;鲀型和鳗型鱼类N∶P最高,侧扁型和纺锤型最低。

图4 不同体型鱼类碳、氮、磷元素含量和比值箱线图Fig.4 Box diagram of the contents and ratios of C,N and P in fishes with different body shapes
2.3 不同食性鱼类生态化学计量特征
方差分析结果表明不同食性鱼类的C含量、N含量、P含量、C∶N和C∶P具有显著差异(P<0.05)(图5)。杂食性鱼类的C含量和C∶P显著低于草食性、滤食性和肉食性鱼类;滤食性和杂食性鱼类N含量最高,肉食性鱼类次之,草食性鱼类最低;草食性和肉食性鱼类C∶N显著高于滤食性和杂食性鱼类;杂食性和肉食性鱼类P含量显著高于草食性和滤食性鱼类;N∶P在各食性鱼类之间无显著差异(P>0.05)。

图5 不同食性鱼类碳、氮、磷元素含量和比值箱线图Fig.5 Box diagram of the contents and ratios of C,N and P in fishes with different feeding habits
2.4 鱼类生态化学计量特征相关性
北部湾鱼类P含量和C含量呈显著负相关关系(R2=0.296,P<0.01);C∶N和N含量、C∶P和P含量、N∶P和P含量均呈显著负相关关系(R2=0.782,P<0.01;R2=0.966,P<0.01;R2=0.877,P<0.01);C∶N和C含量、C∶P和C含量、N∶P和C含量、N∶P和N含量呈显著正相关关系(R2=0.388,P<0.01;R2=0.472,P<0.01;R2=0.160,P<0.01;R2=0.286,P<0.01)(图6);C∶P和P含量、N∶P和P含量的相关性最强(R2>0.8),说明北部湾鱼类C∶P比和N∶P比变化主要由P含量变化决定。
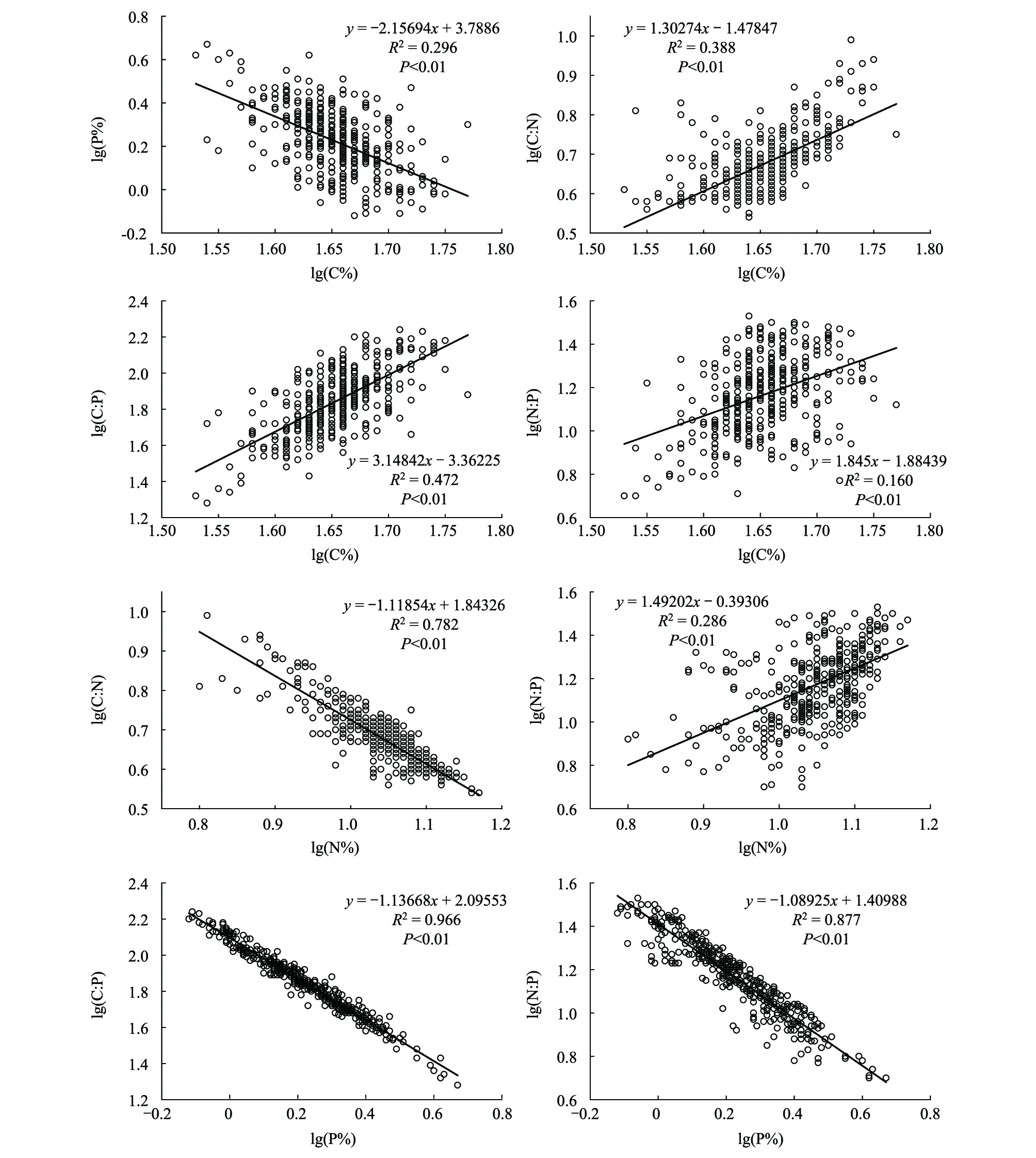
图6 北部湾海域鱼类碳、氮、磷含量及其比值的相关性分析Fig.6 Correlation analysis results of relationships between C,N,P contents and their ratios of fishes from the Beibu Gulf
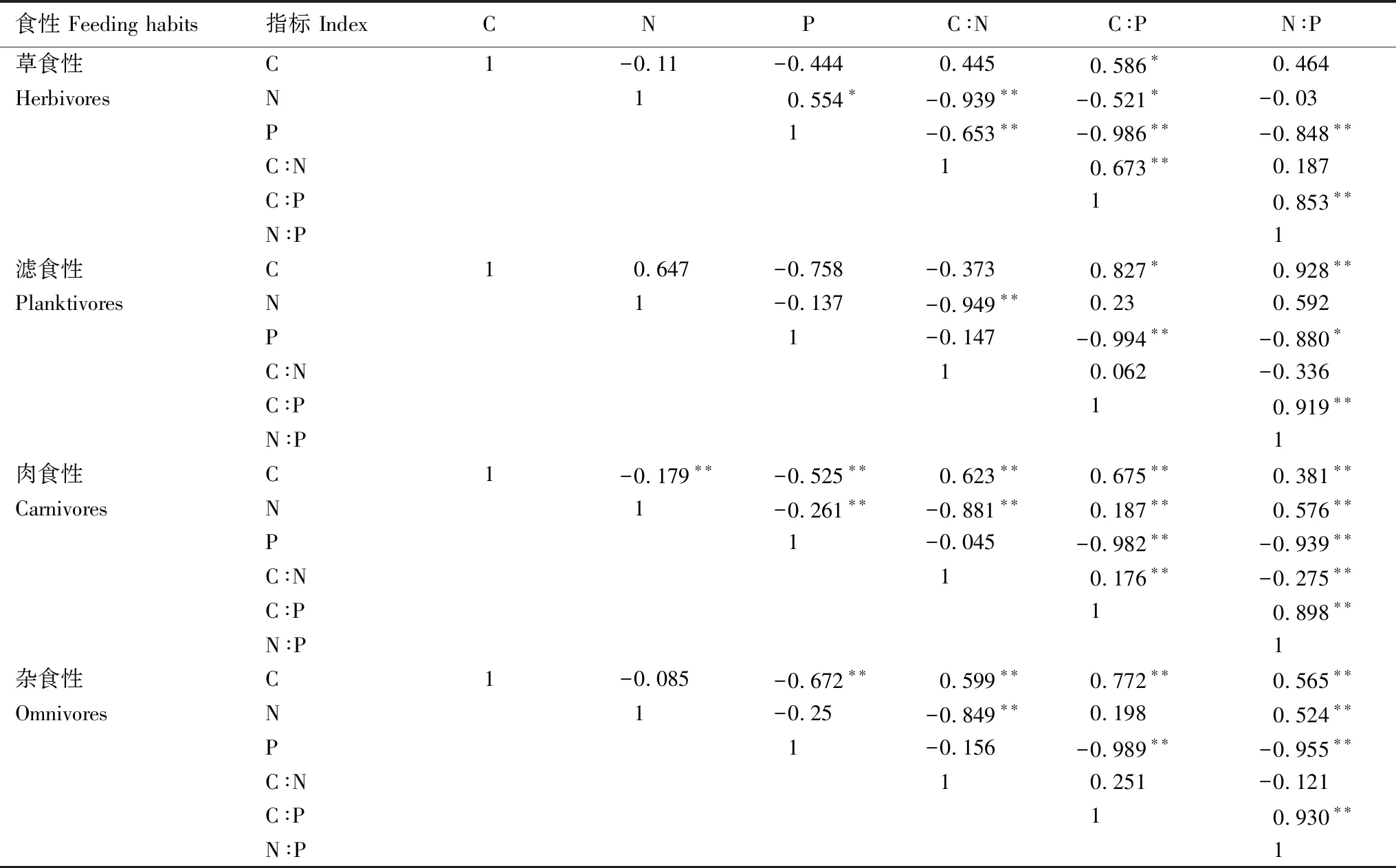
食性 Feeding habits指标 IndexCNPC∶NC∶PN∶P草食性C1-0.11-0.4440.4450.586∗0.464HerbivoresN10.554∗-0.939∗∗-0.521∗-0.03P1-0.653∗∗-0.986∗∗-0.848∗∗C∶N10.673∗∗0.187C∶P10.853∗∗N∶P1滤食性C10.647-0.758-0.3730.827∗0.928∗∗PlanktivoresN1-0.137-0.949∗∗0.230.592P1-0.147-0.994∗∗-0.880∗C∶N10.062-0.336C∶P10.919∗∗N∶P1肉食性C1-0.179∗∗-0.525∗∗0.623∗∗0.675∗∗0.381∗∗CarnivoresN1-0.261∗∗-0.881∗∗0.187∗∗0.576∗∗P1-0.045-0.982∗∗-0.939∗∗C∶N10.176∗∗-0.275∗∗C∶P10.898∗∗N∶P1杂食性C1-0.085-0.672∗∗0.599∗∗0.772∗∗0.565∗∗OmnivoresN1-0.25-0.849∗∗0.1980.524∗∗P1-0.156-0.989∗∗-0.955∗∗C∶N10.251-0.121C∶P10.930∗∗N∶P1
不同食性鱼类C∶P和C含量均呈显著正相关关系(P<0.05);不同食性鱼类C∶N和N含量、C∶P和P含量、N∶P和P含量呈显著负相关关系(P<0.01)(表2)。
2.5 鱼类Ca和P的化学计量关系
相关性分析发现,北部湾鱼类Ca含量和P含量呈显著正相关关系(R2=0.919,P<0.01),P含量随着Ca含量的增加而增加(图7)。
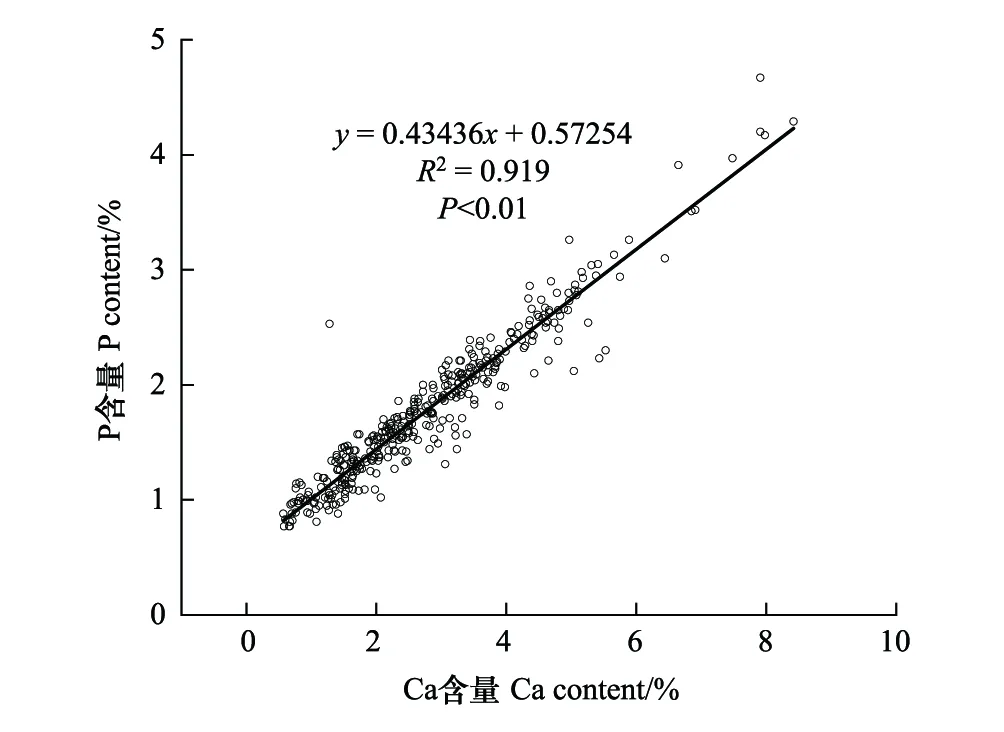
图7 鱼类钙磷含量相关性分析Fig.7 Correlation analysis results of between Ca and P in fishes
3 讨论
3.1 北部湾鱼类元素组成与变化特征
北部湾鱼类的C、N、P元素含量变化较大(图2),而与C含量和N含量相比,P含量变幅更大(变异系数P >N >C)。不同鱼类骨骼和鳞片的种间分布差异和富含C和N的内脏被去除导致P含量变化较大,C、N含量变化较小。李培培[15]、孙培英[16]分别对千岛湖和长江口鱼类的生态化学计量特征研究发现,鱼类P含量的变化均高于C和N含量的变化。国外学者在委内瑞拉热带溪流的研究中也发现鱼类体内的 C、N 含量变化较小,而P含量变化较大[43]。一般认为,鱼类化学计量在物种水平上差异很大,尤其是P含量水平上[31]。Hendrixson等人[44]提出鱼体P含量与系统发育存在紧密联系,P含量是物种间重要的特征,是导致鱼类元素组成多样化的重要因子。
研究表明鱼类科级分类水平是生态化学计量学最佳的预测因子,鱼类生态化学计量特征在科级水平上差异较大[45—47]。主成分分析(PCA)发现北部湾鱼类物种在科间差异较大,科内差异较小(图3)。鱼类生态化学计量上存在系统发育印记,不同物种间营养成分的模式为生态化学计量学模式增加了额外的、潜在的基于系统发育的变异,如热带鱼类生态化学计量变化(特别是磷含量)在科之间比在科内部变化更大[42,44]。营养物质的浓度(特别是磷)在不同物种间大不相同[44,48—49],可能与不同物种在向骨骼和鳞片分配营养物质方面的差异有关。
3.2 北部湾不同体型鱼类生态化学计量特征差异
生物体元素组成差异很大程度上取决于其对关键部位的富集,如植物富含C的纤维素和木质素,节肢动物富含C和N的甲壳素和肌肉及脊椎动物富含P的骨骼等,这些部位在不同生物体之间分布和占比不同,结构和形态差异显著[26,50]。鱼类体型较其他脊椎动物更为多样化,其化学计量特征可能会随体型变化较大,因为其肌肉、骨骼和鳞片等组织在不同体型鱼类之间分配不同[24],且组织结构特征复杂多样。本研究不同体型鱼类的C、N、P含量及比值均存在显著差异(图4),尤其是鳗型鱼类N∶P显著高于其它体型鱼类,主要因鳗型鱼类有较小的表面积/体积、头部/全长比例,头部骨骼含P量较大,身体肌肉含量丰富,且鳗型鱼类体表无鳞。这与Tanner等[2]的研究结果类似,即细长的鱼类具有较高的N∶P。本研究发现鲀型鱼类有较高的N含量和较低的C含量,与徐羊羊等[51]研究发现兔头鲀属于高蛋白、低脂肪鱼类结果相吻合。另外,本研究还发现C含量较高的体型鱼类P含量较低(高C∶P),再次验证了鱼类C含量与P含量呈负相关(图6)。
3.3 北部湾不同食性鱼类生态化学计量特征差异
自然界食物资源的可获得性是高度可变的。海洋鱼类食物来源广泛、种类繁多,食物化学计量差异很大。因此,鱼类生态化学计量的种间差异可能归咎于食物来源的差异[19]。消费者利用不同的食物来源来满足其能量和营养需求[30]。例如,以高质量肉食动物为食的动物比以低质量草食或腐食为食的动物具有更高的N%或P%和更低的C%[30—31];Hendrixson发现淡水鱼类P含量与食性相关,肉食性鱼类P含量显著高于杂食性鱼类和草食性鱼类[44];在许多鱼类中,以C∶N为代表的脂质含量随着食物浓度的增加而增加[52],并且当食物现存量高时,消费者能获得更多的脂质。另外,C∶N∶P含量在不同生物类群体内积累存在差异,研究发现从藻类到浮游动物再到鱼类,C∶P值的平均值随营养级降低[53]。
北部湾水域水质特征复杂多样,饵料来源丰富。水体营养状态会影响基础资源和饵料的组成,进而直接或间接影响消费者的元素含量。研究表明基础资源质量的变化限制了较高营养级的元素含量,其变化不仅影响初级消费者,还影响多个营养水平[53],如水体富营养化可影响杂食性鱼类的化学计量特征[19]。杂食性鱼类的化学计量比随着浮游植物数量和质量的变化而变化,而浮游植物的数量和质量受到营养物质和光线的调节[54]。因此,生物生态化学计量学的变化比生物特征的变化更有可能反映环境条件的变化,而环境条件的变化影响了消费者食物或基础资源中元素的可获得性。因此动物的元素组成比最初认为的更灵活,动态平衡更少[55]。尽管单个鱼类物种可能以稳态的方式调节其自身化学计量,但并非所有鱼类都具有相同的元素组成。相反,不同物种的各种元素含量差异很大,尤其是磷[31]。
3.4 北部湾鱼类C、N、P含量及其比值关系
北部湾鱼类C∶P和P含量、N∶P和P含量均呈显著负相关关系(R2=0.966,P<0.01;R2=0.877,P<0.01);各食性鱼类的C∶P和P含量、N∶P和P含量也均呈显著负相关关系(P<0.01)(图6,表3)。研究发现底栖动物体内N∶P和P含量呈显著负相关关系[13](r=-0.795,P<0.001);三峡水库小江流域鱼类肌肉的C∶P和P含量、N∶P和P含量都呈显著负相关关系[36](r=-0.971,P<0.01;r=-0.911,P<0.01),本研究结果与之相似。在脊椎动物的成长过程中更多的磷被用于骨骼的形成,因此,随着个体大小的增加,全身的磷含量可能会增加,导致C∶P和N∶P下降[56]。
3.5 北部湾鱼类Ca和P的化学计量关系
北部湾鱼类P含量与Ca含量呈显著正相关性(图7)。P含量的变化与鱼类骨骼和鳞片的形成有关,脊椎动物骨骼中的主要矿质元素(Ca,P)在其营养生理中起着重要作用,说明脊椎动物对矿质元素的需求也会影响生态系统的养分循环。鱼类骨骼主要是由羟基磷灰石构成,其钙磷摩尔比为2.16:1[57],从海洋鱼类到淡水鱼类,其骨骼中Ca含量占鱼体总Ca含量的79%—95%[23]。鱼类体型和鳞片类型可以部分解释鱼类的磷含量较低,如鲤科和鲑科鱼类身体呈纺锤型,具有软鳍条和体被圆鳞;而磷含量较高的太阳鱼科和鲈科鱼类,身体常呈侧扁型,具有硬棘和体被栉鳞,或者具有高度骨化的头骨[2,47]。因此,骨骼和鳞片的差异可导致鱼体磷浓度的显著变化。另外,鱼类个体大小可以通过骨骼异速生长来改变其元素组成,因为骨骼占体重的比例随着个体增大而增加[46,56]。脂质含量、肌肉占比和个体大小等特征也会影响元素组成[56—58]。富含碳的脂质和富含氮的肌肉含磷量少,因此脂质和肌肉的增加会稀释全身磷的比例。Vrede等[24]研究发现全鱼的P含量为3.06%,肌肉的P含量为0.94%,全鱼的P含量高于肌肉的3倍多;李培培[15]研究千岛湖鲢鳙发现,P、Ca含量均为头骨>鳍条>鳃>鱼骨>鱼皮>肌肉,而C含量为肌肉>鱼骨>鱼皮>鳃>鳍条>头骨。因此,鱼类肌肉、骨骼、鳞片和其他组织的占比不同可导致鱼类生态化学计量特征的种间差异。
4 结论
本研究结果表明北部湾鱼类的生态化学计量特征存在显著的种间差异,P含量和Ca含量呈显著正相关;鱼类体型和食性不同导致鱼类生态化学计量特征存在显著差异,且鱼类并非保持严格的生态化学计量平衡。北部湾水域面积宽阔,环境特征复杂多样,鱼类群落结构具有显著的时空差异特征[59—61]。因此,后续研究将深入探讨鱼类生态化学计量特征的时空差异,并结合水生植物、浮游生物、无脊椎动物和水体等生态化学计量特征,探究不同生物体内的生态化学计量特征差异及其与生态因子的关联,为北部湾海域生态环境的保护和治理提供理论支持。
