基于系统发育及功能性状的不同坡向荒漠植物群落构建研究
——以博湖县沙化封禁保护区为例
2023-06-14马紫荆王军强
马紫荆,刘 彬,*,王军强,余 立
1 新疆特殊环境物种保护与调控生物学实验室,乌鲁木齐 830054
2 新疆特殊环境物种多样性应用与调控重点实验室,乌鲁木齐 830054
3 新疆师范大学生命科学学院,乌鲁木齐 830054
揭示和阐明植物群落物种共存及生物多样性维持机制的群落构建是生态学研究的核心论题之一[1]。目前对群落构建机制的解释主要基于确定性过程的生态位理论和随机性过程的中性理论[1],前者强调生境过滤及竞争排斥两种相反的作用力促使生态位分化,以达到物种间共存[2];而后者则认为扩散和随机作用是主要决定因子[3]。随着理论和方法的更新,植物功能性状作为能够反映植物生态适应策略的核心属性[4],为群落构建的研究提供了新的途径。近年来,生态学家们通过构建随机零模型[3],检测系统发育结构和功能结构发散、聚集或是随机的基础上,使生态位理论和中性理论得以不断完善与相互补充,同时检测功能性状的系统发育信号,以此考虑物种进化与外界环境对功能性状的影响[5]。由此可见,系统发育、植物功能性状和生态过程三者紧密相关,功能性状不同,表现出的群落构建机制不同,基于系统发育信号,综合考虑系统发育结构和功能性状结构,可更加准确地探讨驱动物种共存的主要进化和生态过程。
森林生态系统与草原生态系统作为验证物种共存机制的理想场所在以往的研究中倍受关注,而生物多样性较低的干旱荒漠生态系统由于其本身的脆弱性与全球化环境问题使得生物多样性保护与物种共存的研究更加迫切[6]。Qian等[7]发现加利福尼亚干旱地区的群落受生境过滤主导谱系大多呈现聚集的趋势;王诗韵等[8]则发现艾比湖典型植物群落随空间尺度的改变,系统发育结构和功能结构发生改变;而旧金山山峰的植物群落则随干旱程度的增加,表现出物种多样性受生境过滤与竞争排斥的共同作用[9]。可见,荒漠生态系统植物群落物种共存的维持机制尚未取得一致结论,可能随生境、地域、尺度和梯度等因素的不同而结果不尽相同。坡向这样的微地形在降水量极少的干旱荒漠地区,极大程度地改变了生境的异质性,影响着植被对水热、养分和盐分的利用效率,使植物群落的物种组成及维持机制有所差异[10]。目前,国内基于坡向梯度群落构建的研究更多的依据与理论来自于高海拔的青藏高原和华北山地、高原等森林生态系统和草原生态系统[1,11],对于荒漠生态系统的关注相对较少,仅有少数研究学者对喀斯特石漠化区和毛乌素干旱荒漠区基于水分梯度和不同恢复阶段的植物群落展开了研究[12—13]。因此,基于固定、半固定沙丘的不同坡向来探究植物物种共存的驱动因素,有助于理解沙地植物群落对微生境适应的生态策略,可为脆弱的荒漠生态系统的植被群落构建研究提供重要借鉴。
以往对于沙化封禁保护区的研究主要集中在土壤特性、植被盖度及生态效益等方面[14],而对植物群落构建机制研究较少。博湖县国家沙化封禁保护区覆盖面积大,由于其特殊的地理位置,严酷的自然环境,加之以前的人类活动影响,严重威胁到该区荒漠生态系统的稳定性及邻近大湖区的生态环境,而稀疏单一的植被在生态与经济的恢复发展中至关重要,探明沙化封禁保护区的植物物种共存机制是防沙治沙和生物多样性保护的关键。因此,本文以博湖县沙化封禁保护区荒漠植物群落为研究对象,结合环境因子,基于系统谱系和功能性状的方法,探究该地区荒漠生态系统上不同坡向植物群落物种共存的主要进化和生态过程,以期解决以下科学问题:(1)植物系统发育多样性和功能多样性在不同坡向上变化特征如何?(2)影响该地区群落构建的主要环境因子有哪些?期望通过解决这些科学问题为该地区植被恢复与生态重建等措施提供多元的数据,同时丰富荒漠生态系统植物群落构建的理论研究。
1 材料与方法
1.1 研究区概况
博斯腾湖县国家沙化封禁保护区,即博湖南岸阿克别勒库姆沙漠封禁保护区,地点位于博湖县扬水站至白鹭洲以南,封禁范围为86.78°—87.04°E,41.80°—41.90°N,封禁面积达10723 hm2(图1)[15]。阿克别勒库姆沙漠是新疆十大沙漠之一,位于博湖县博斯腾湖南岸,库鲁克塔格山北麓的南山地区,从阿訇口起至湖东南为止,地跨博湖与和硕两县,长约75 km,宽度一般为 10 km 左右,最宽 23 km,最窄3 km,沙丘主要由复合式沙丘链组成,高出附近地面一般达20—100 m,沙行走向近乎南北(https://www.zsbeike.com/),参照沙丘有关研究[16],经过初步踏查,发现该地区沙丘由丘间、阴坡、阳坡3个不同地形组成,本研究坡向采用0°—360°的方位角转换为TRASP(Transformation of aspect)指数来划分[17]。据博湖县气象资料显示,该地区年平均气温9.1℃,年均降水量93.3 mm,蒸发量为1853 mm,年日照2793.6—3136.5 h,无霜期219 d左右[18]。

图1 研究区示意图Fig.1 Schematic diagram of the study area
1.2 样方调查与指标测定
1.2.1植物功能性状调查及测定
2021年7月,沿东北走向对博湖县国家沙化封禁保护区所处的沙漠区边缘进行野外植物调查,每500 m左右为间距选择主要沙丘形态——南北走向的纵向沙垄10个,使用 360°罗盘测定坡向,分别在阳坡、阴坡、丘间低地3种立地条件下各布设3个10 m×10 m的木本样方共90个,沿对角线设置3个1 m×1 m的草本样方共270个,记录坡度坡位以及样方中植物种类、数量、高度、冠幅。
在10个沙垄的3个坡向上每种植物选取3株长势良好、大小基本一致的植物个体作为取样植株,每株随机采集东南西北4个方向健康完整且大小一致的叶片各20 g,混合作为同一样叶装入保鲜箱带回实验室[19]。取不同沙垄3个坡向的每个物种3—5片成熟展开的叶片进行相关功能性状指标测定并计算其均值,采用便携式叶绿素测定仪(SPAD-502,Minolta Co,Japan),避开叶脉,测定叶片叶绿素含量(SPAD)值;选用精度为0.001 mm的电子游标卡尺测量叶中部分的叶厚度(Leaf thickness,LT);选用精度为0.0001g的电子天平称量叶片鲜重后,置于60℃的烘箱中烘至恒重称取叶片干重(Leaf dry weight,LDW);采用CI-203(CID,Inc,USA)手持式激光叶面积仪测定叶面积;比叶面积(Specific leaf area,SLA)=叶片面积/叶片干重;叶片含水量(Leaf water content,LWC)=(叶片鲜重-叶干重)/叶片干重,叶干物质含量(Leaf dry matter content,LDMC)=叶片干重/叶片鲜重。其余叶片烘干磨成粉,用高氯酸—硫酸消化法测量叶片N含量;用酸溶—钼锑抗比色法测定叶片P含量;用重铬酸钾容量—外加热法测定叶片C含量[13]。
1.2.2土壤理化指标的测定
在每个木本样方内随机挖取0—30 cm土样200 g,带回实验室在室温条件下自然风干,过80目筛,参照土样指标常规方法[20],用重铬酸钾滴定法测定土壤有机质(Soil organic matter,SOM)、高氯酸—硫酸消化法测定全氮(Total nitrogen,TN)、酸溶—钼锑抗比色法测定全磷(Total phosphorous,TP)、酸溶—原子吸收法测定全钾(Total potassium,TK)、碱解蒸馏法测定有效氮(碱解氮)(Available nitrogen,AN)、碳酸氢钠浸提—钼锑抗比色法测定土壤中有效磷(Available phosphorous,AP)、乙酸铵浸提—火焰光度计法来测定有效钾(Available potassium,AK)、干渣法测定全盐(Total salt,TS)、烘干称重法测定土壤含水量(Soil water,SW)、雷磁pH计测定pH。
1.2.3系统发育树的构建
样方中共记录到13种主要物种,隶属7科,13属,其中灌木植物有6科7属7种,草本植物有2科6属6种。对其中12种被子植物物种名录进行核对,按照科/属/种的格式进行统计整理,通过在线phylomatic软件(http://www.phylodiversity.net/phylomatic),并选用Zanne等[21]维管植物进化树计算物种间进化枝长,其分支长度代表了演化时间(利用化石数据推定)[22],用MEGA7.0软件构建系统发育树(图2)。本研究中使用的谱系树未包括裸子植物膜果麻黄(Ephedraprzewalskii)。

图2 调查样方中植物物种的系统发育树 Fig.2 Phylogenetic tree of plant species in the investigation quadrat
1.2.4功能性状的系统发育信号检测
采用Blomberg等[23]提出的K值系统发育信号检测法检验研究区植物功能性状的系统发育信号强度。通过R 4.1.3中ape和picante包对功能性状的系统发育信号K值进行计算,并随即置换999次以检验其显著性(P<0.05)。K=1 对应布朗运动的随机进化模型,表示物种随机进化,功能性状结构和系统发育结构无关;K>1 为检测到比随机进化模型更强的系统发育信号,功能性状保守,功能性状结构和系统发育结构具有一致性;K<1 表示检测出比随机进化模型更弱的系统发育信号,物种趋同性进化,即群落系统发育和性状格局并不完全一致[8]。
1.2.5群落构建中的系统发育多样性和功能多样性
系统发育多样性采用Faith[24]提出的量化谱系树进化总枝长的群落谱系多样性指数(phylogenetic diversity,PD)和Webb[2]提出的系统发育结构指数,即净谱系亲缘关系指数(net relatedness index,NRI)与最近亲缘关系指数(nearest taxon index,NTI)来表达。NRI和NTI分别由群落中物种对的平均谱系距离(MPD)和最近相邻谱系距离的平均值(MNTD)与零模型随机化相比较所得到的。若NRI/NTI值大于零模型,则该样方群落的系统发育结构聚集,环境过滤起主导作用;若NRI/NTI值小于零模型,则该样方群落的系统发育结构发散,限制相似性起主导作用;若NRI/NTI值等于零模型,则该样方群落的系统发育结构随机[8]。群落系统发育结构指数的计算公式如下:
式中,MPD表示种间平均系统发育距离,MPDobs为MPD观测值,MPDrand表示999次随机模拟零模型的群落MPD值,mean(MPDrand)为零模型模拟下随机群落MPD的平均值,sd(MPDrand)为零模型模拟下随机群落MPD的标准差。同理可知NTI中各值含义。
功能多样性可用功能丰富度指数(Functional richness,FRic)和功能结构指数来表示。功能结构与群落系统发育结构相类似,以群落平均配对性状距离的标准效应值trait SES(MPD)和最近性状距离的标准效应值trait SES(MNTD)来表示,分别是由平均配对性状距离(trait(MPD))和最近性状距离(trait(MNTD))与零模型生成的格局进行比较所得到。当trait SES(MPD) /trait SES(MNTD)>0,则样方物种的功能性状结构聚集,环境过滤作用为主要驱动因素;若trait SES(MPD) /trait SES(MNTD)<0,则样方物种的功能性状结构发散,限制相似性起主导作用;若trait SES(MPD) /trait SES(MNTD)=0,则样方物种的功能性状结构随机,中性理论占主导。具体公式如下:
式中,MPD 表示种间平均性状距离,MPDobs为MPD观测值,MPDrand表示999次随机模拟零模型的群落MPD值,mean(MPDrand)为零模型模拟下随机群落MPD的平均值,sd(MPDrand)为零模型模拟下随机群落 MPD 的标准差。同理可知trait SES(MNTD)中各值含义[8]。
通过R 4.1.3中PD、vegan、picante、FD 包完成系统发育多样性指数 NRI、NTI、PD和功能性状多样性指数 trait SES(MPD)、trait SES(MNTD)、FRic的计算;使用vegan包对环境因子进行主成分分析。通过SPSS 20.0完成环境因子间的相关性及线性回归分析;所有作图均在Origin中实现。
2 结果与分析
2.1 不同坡向土壤环境因子分布特征及植物群落的优势种组成
由表1可知,不同坡向土壤环境因子具有异质性,阴坡土壤含水量显著高于丘间和阳坡(P<0.05);丘间土壤全磷、有效钾含量显著高于阴坡和阳坡(P<0.05),可能是由于丘间较缓的坡度积累了更多的土壤养分[25]。因此,除酸碱度、全钾外,丘间和阴坡土壤因子指标均大于阳坡,说明丘间、阴坡土壤肥力较丰富,土壤质地更适合植物生长。
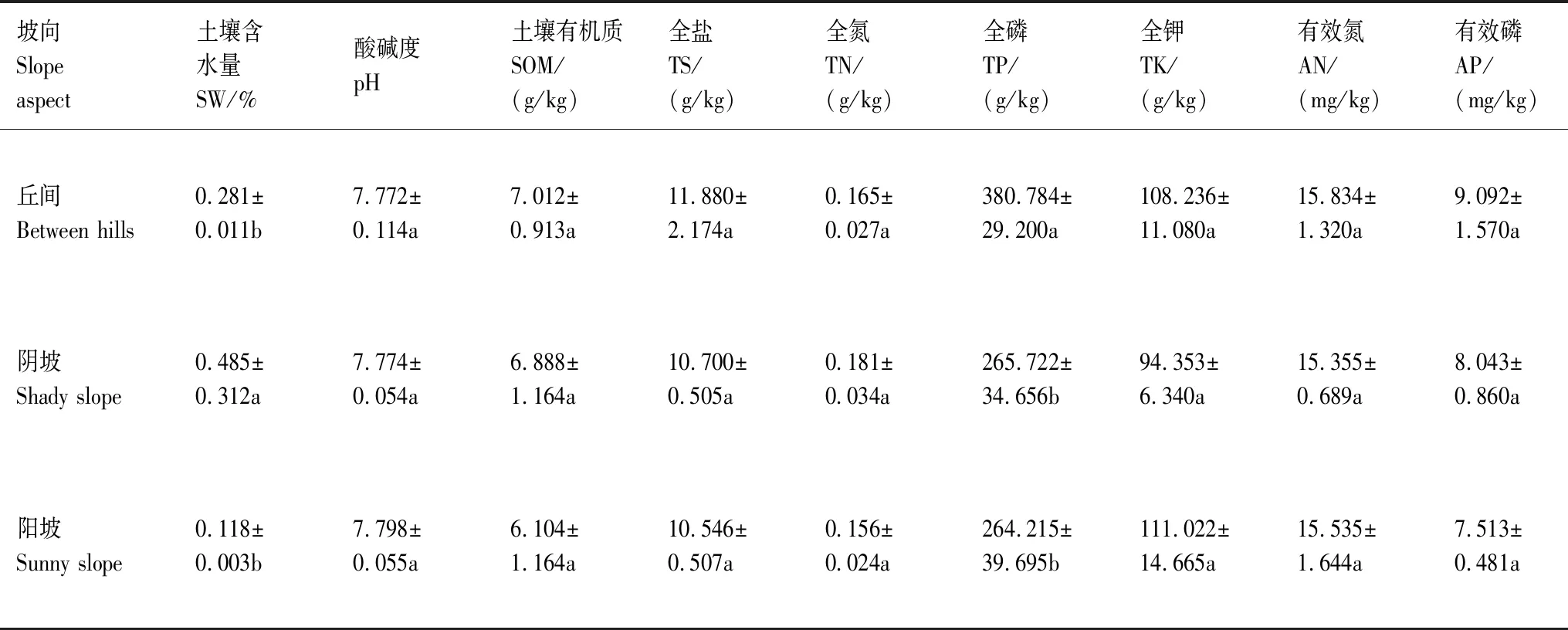
表1 不同坡向土壤环境因子分布特征及优势种组成Table 1 Distribution Characteristics of soil environmental factors and composition of dominant species in different slope directions
研究区植被主要以灌木和草本为主,其中不同坡向主要的灌木优势种有膜果麻黄(Ephedraprzewalskii)、黑纱蒿(Artemisiaordosica)、梭梭(Haloxylonammodendron)、沙拐枣(Calligonummongolicum)(表1)。总体上,不同坡向主要的优势种以耐旱的沙漠植物小灌木-黑纱蒿及藜科草本植物为主,反映了博斯腾湖沙化封禁保护区环境严苛、干旱的条件下,植被组成和物种生活型逐渐趋于单一。
2.2 植物群落物种功能性状的系统发育信号
由表2可知,样地中植物群落物种的10个功能性状指标的K值均小于1,表现出不具有较强的系统发育保守性。其中叶片碳含量表现出显著的比随机进化模型更弱的系统发育信号(P<0.05),其余9个功能性状均未表现出显著的系统发育信号(P>0.05)。说明该区域植物系统进化关系对不同功能性状的影响程度不同,除叶片碳含量相对影响程度较大外,整体受系统进化影响较小,主要表现在干旱区环境条件下,植物功能性状长期的趋同适应,植物功能性状进化存在一定的非保守性。叶片碳含量表现出系统发育反信号(K<1,P<0.05),说明系统发育距离相近的物种,但其功能性状亲缘关系反而疏远,因而综合系统发育信号检验结果,可以初步判定群落物种功能性状结构和系统发育结构具有较大差异。
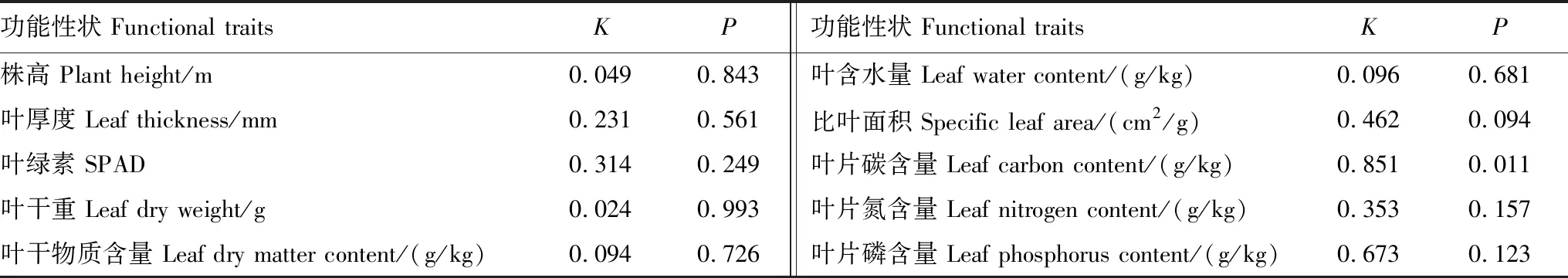
表2 植物功能性状的系统发育信号Table 2 The phylogenetic signal of plant functional traits
2.3 不同坡向的系统发育多样性和功能多样性
坡向等小尺度范围的地形变化可以使得生境条件发生改变,如光辐射强度、光辐射时间、土壤温度、湿度及养分含量等,从而影响局域群落植物群落的生长、分布和物种组成等[14]。系统发育结构指数 NRI和NTI 在3个不同坡向均表现为发散结构,且各坡向不存在显著差异(P>0.05)(图3),谱系多样性指数PD在各坡向无显著差异(P>0.05)(图4)。说明研究区干旱恶劣的环境及匮乏的资源不利于植被生长与繁殖,植物群落结构简单,可能由亲缘关系较近且资源利用方式相似的物种相互竞争形成或是由亲缘关系较疏远的物种相互促进而形成[6];其中阴坡有部分样方表现出聚集结构(图3),说明物种的共存受环境过滤和限制相似性共同作用。

图3 植物群落物种的NRI和NTI在不同坡向上的分布Fig.3 Distribution of NRI and NTI of plant community in different slope directions

图4 植物群落物种的PD和FRic在不同坡向上的分布Fig.4 Distribution of PD and FRic of plant community in different slope directions
由图5可知,功能结构指数 trait SES(MPD)和 trait SES (MNTD)在 3个坡向上表现不同的结构,丘间和阴坡功能结构表现为发散,限制相似性起主导作用,而在阳坡上显著不同(P<0.05),植物功能性状主要受到环境过滤的驱动而表现出聚集模式,这可能是由于丘间与阴坡水热条件相对充足,植物通过竞争有限的资源而具有一定的适应策略,从而形成不同的生态位,因此功能丰富度指数FRic相较于阳坡偏高(图4),功能性状所占据的空间更大,生态空间利用程度更高[26]。

图5 植物群落物种的trait SES(MPD)和trait SES(MNTD)在不同坡向上的分布Fig.5 Distribution of trait SES (MPD) and trait SES (MNTD) of plant community in different slope directions
2.4 影响群落构建的环境因子筛选
为筛选出影响研究区植物群落的主要环境因子,对环境因子进行PCA主成分分析,并通过Pearson相关性分析观察PCA判定的主要环境因子与其他环境因子的相关性[27]。PCA分析显示前3轴的环境因子解释量分别为20.3%,17.2%,13.0%,负荷绝对值大于0.5的环境因子有:坡向、土壤含水量、有机质、全盐、全氮、全钾和有效磷(表3)。由表4可知,土壤含水量与坡向负显著相关(P<0.05);有机质与全氮有显著正相关关系(P<0.05);全盐和pH极显著正相关(P<0.01);全钾与全氮、有效钾极显著正相关(P<0.01);有效磷与全磷、有效氮显著正相关(P<0.05)。因此,可以推测土壤含水量、有机质、全盐、全钾、有效磷是研究区环境变化的重要因子。

表3 研究区环境因子的主成分分析Table 3 Principal component analysis (PCA) of environmental factors in the study area
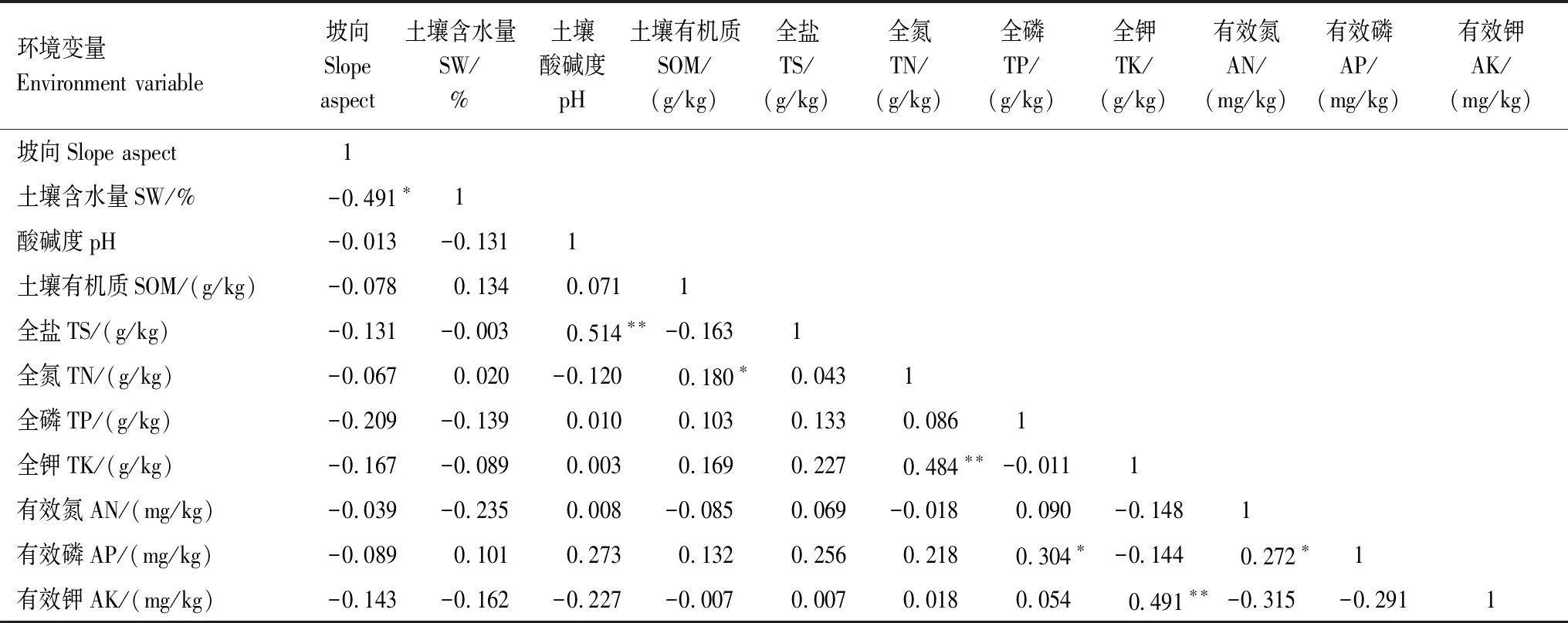
表4 环境因子间的相关性Table 4 Correlation between environmental factors
运用单元线性回归分析环境因子与功能多样性指数间的相关性,可以进一步筛选博湖县沙化封禁保护区的植物群落物种共存的环境筛。研究结果表明,有效磷与FRic负显著线性相关(P<0.05);土壤含水量对trait SES(MNTD)有显著影响(P<0.05);有机质对trait SES(MNTD)影响显著(P<0.05),而其余环境因子对多样性指数没有显著影响(P>0.05)(未在表5中列出)。

表5 多样性指数与环境因子间的线性回归分析Table 5 Linear regression analysis between diversity index and environmental factors
3 讨论
3.1 功能性状的非保守性进化
共存物种的生态和进化相似性是验证群落构建机制的关键信息之一,通过检验植物群落功能性状的系统发育信号,可更加准确地推断驱动群落构建的主要生态过程[28]。功能性状进化模式不同,生态过程构建的群落功能结构和系统发育结构有所差异。性状进化保守的情况下,可能由生境过滤会聚集相似的近缘物种,也可能由竞争排斥导致具有相似特征的近缘物种分散;而性状进化非保守的情况下,通过生境过滤作用保留下的性状相似物种,可能亲缘关系并不相近,或是通过竞争排斥作用保留性状不同的共存物种,其亲缘关系规律不一定[29]。通过检验博湖县沙化封禁保护区样方中植物物种10个功能性状的系统发育信号发现,K值均小于1,说明该研究区植物群落受进化影响较小,功能性状未表现出系统发育保守性,系统发育结构和功能结构不具有匹配对应关系,可能由于植物群落在世代更新的过程中,受生物和非生物因素影响(如生境过滤、竞争排斥等),选出一组与沙漠干旱脆弱条件相适应的具有相似功能性状的物种,从而增强了群落内物种性状的趋同性[30]。不同于本研究结果,也有许多研究学者发现群落物种功能性状具有显著的系统发育信号,系统发育结构与功能结构具有一致性[27],这是因为不同研究区生境异质,植物群落及所选择的植物功能性状不同,其结果也所不同,甚至同一植物群落选择不同的植物功能性状,结果也有所差异[31];同时Swenson[32]也表明,系统发育关系的远近并不能和功能性状的相似性一一对应,还应该考虑所选择分析的性状种类、数量、竞争能力等级和研究尺度等影响[33]。值得注意的是,研究结果表明仅叶片碳含量表现出显著的比随机进化模型更弱的系统发育信号(K=0.851,P<0.05),较其他功能性状受进化关系的影响较大,但K值仍小于1。综合前人研究发现在给予土壤施氮和施碳的情况下,不同植物的叶片碳含量随土壤碳、氮含量变化具有相似的响应方式,且叶片碳含量及其可塑性均未检测出显著的系统发育信号,说明相对于物种遗传背景,叶片碳含量主要受环境变化的影响[34],因此研究区群落物种功能性状整体呈非保守性进化,受生态过程影响较大。
3.2 不同坡向系统发育多样性和功能多样性变化及环境因子响应
群落构建的成因受多种因素的影响与控制,在阳坡—阴坡梯度上由于地形学的缘故100 m的距离可能会造成环境因素极大的变化,植物群落组成和物种多样性等随之也会发生很大的改变[25]。本研究表明,系统发育结构在丘间、阴坡、阳坡3个微生境上整体表现为发散模式,主要的优势科有菊科和藜科,而藜科占绝对优势,其不同属植被(如盐生草、刺沙蓬等)具有良好的耐盐碱、抗风沙能力,面对干旱的环境压力不同种、属植被利用有限的资源发生竞争排斥,从而引起生态位的分化以实现物种共存[35]。不可忽略的是,Solivers[36]等人发现西班牙半干旱的草原生态系统中,植物-植物相互作用增加了系统发育结构分散,而土壤水分条件的改善又促进了系统发育聚类,两种相反的作用力使得整体系统发育结构随机,而本研究区属极端干旱的荒漠生态系统,土壤含量较少,水分等环境资源不足以完全缓解植物间因竞争排斥而形成的发散结构,因此,虽然阴坡上植物群落整体受生境过滤和限制相似性共同作用的影响,但竞争排斥占主导,系统发育结构整体呈现发散模式。综上所述,尽管研究区相较于森林和草原生态系统通常被认为是相对严苛的环境,但生境过滤作用并没有决定该地区系统发育结构,一方面可能是干旱的环境条件下会由扩散能力和胁迫耐受性较强的物种首先定植生长,但迫于资源的匮乏,近缘物种间相互竞争排斥,种间生态位分化加强,最终导致系统发育结构发散;另一方面典型荒漠内部生境相对均匀,系统发育结构在坡向这样微生境梯度上受生境过滤影响相对于海拔等大尺度的生境梯度作用较小[35],微气候的改变不足以作为谱系聚集的重要过滤作用,因此本研究中环境因子对谱系多样性指数和系统发育结构指数都不具有显著影响。
功能结构指数在不同坡向上具有显著差异(P<0.05),丘间和阴坡功能结构表现为发散,限制相似性起主导作用。由于博湖县沙化封禁保护区北临博斯腾湖,常年受以北风为主的湖风影响[37],使得阴坡面水汽吸收更为充足,蒸发较少,受水养条件更好,土壤含水量显著高于丘间和阳坡(P<0.05);而丘间土壤养分含量较高,土壤肥力更为丰富,分析原因可能是由于土壤的肥力和湿度与坡度陡峭程度呈反比,从而积累了更多的土壤有效成分[25];因此,丘间和阴坡总生态位更宽,物种较丰富,竞争排斥更为激烈,以形成物种间互补地最大化利用资源,功能丰富度指数相对较大。同时,研究区丘间和阴坡分布着以黑纱蒿、膜果麻黄、梭梭等灌木为主的优势种,可截获土壤风蚀颗粒、沙尘和凋落物等,促进灌丛“肥岛”,改善了土壤肥力[38]。阳坡则显著不同(P<0.05),功能结构表现出聚集模式,生境过滤为驱动因素,由于阳坡光照强且时间长,水分蒸发量较大,环境筛除需要充足水分的物种,群落主要由更耐旱的植物物种组成,性状趋同。基于环境因子分析,阳坡由于水热资源的影响,植被覆盖度相较丘间、阴坡较低,再加之风蚀作用于土壤可蚀性颗粒,导致有机质通过土壤-植物-凋落物再补充进入土壤系统的含量易流失[39],受有机质的影响有机碳库、有效氮等土壤养分减少,环境愈加恶劣使得阳坡功能结构聚集。为适应干旱的环境,植物通过获取土壤中的磷含量以此来提高自身功能性状的抗旱性[40],植株的保水能力增强的同时各功能性状趋同进化,功能丰富度指数较低。
4 结论
博湖县沙化封禁保护区系统发育结构和功能结构不具有一致性,植物群落物种共存及生物多样性的维持受生态位过程主导,以土壤含水量、有机质和有效磷为主的关键环境因子筛选出具有耐旱、耐盐碱、抗风沙能力强的植物定植生长;丘间和阴坡由于地形优势,环境和资源得以缓和,近缘物种竞争加剧,植物通过塑造自身功能性状的表型实现对有限资源的最大化利用,进一步实现物种共存;反映了生境过滤和限制相似性对群落物种多样性的共同促进作用及不同物种适应微生境的不同生存策略。博湖县沙化封禁保护区生态环境脆弱,植被生态修复尤为重要,一方面在未来的研究中可以选取地上、地下不同的功能性状结合时间尺度进一步扩展物种多样性的研究;另一方面,在生态重建过程中应在基于当地气候条件的前提下综合考虑不同坡向的土壤水分、养分以及土壤种子库,选择耐旱的荒漠物种来实现人工造林和对林木的抚育。
