杧果金煌×热农1号后代果实糖酸组分遗传分析
2023-06-12李星郑斌许文天马小卫王松标钱敏杰武红霞
李星 郑斌 许文天 马小卫 王松标 钱敏杰 武红霞

摘要:【目的】探讨杧果杂交后代果实糖酸组分等品质性状的遗传变异特点,为科学选配亲本提供理论依据。【方法】以杧果高糖品种金煌与低糖品种热农1 号及其杂交F1代47 个单株为材料,利用高效液相色谱方法(HPLC)测定果实糖酸组分含量,利用紫外分光光度计测定果实类胡萝卜素及总黄酮含量。【结果】蔗糖、果糖和葡萄糖是杂交后代果实的主要糖组分,且以积累蔗糖为主,各种糖组分含量均呈正态分布,总糖含量高于双亲的单株较多,超高亲率为38.30%。抗坏血酸、苹果酸、乙酸、甲酸和柠檬酸是主要酸组分,同时含有少量的草酸、酒石酸,且以积累抗坏血酸为主,各有机酸组分含量呈偏态分布,果实总酸含量低于双亲的单株比较多,超低亲率为57.45%。类胡萝卜素含量基本呈正态分布,含量高于双亲的单株比较多,超高亲率为65.96%。总黄酮含量基本呈正態分布,含量低于双亲的单株比较多,超低亲率为53.19%。【结论】蔗糖、果糖和葡萄糖是由多基因控制的数量性状;有机酸是由1 个或多个主效基因控制的数量性状;类胡萝卜素和总黄酮是由多基因控制的数量性状。杂交后代果实糖酸含量倾向高糖低酸亲本,且受母本影响较大;类胡萝卜素含量倾向高值亲本,受父本影响较大;总黄酮含量倾向低值亲本,受母本影响较大。
关键词:杧果;F1代;糖组分;酸组分;遗传
中图分类号:S667.7 文献标志码:A 文章编号:1009-9980(2023)04-0630-09
杧果(Mangifera indica L.)属于漆树科杧果属常绿果树,是重要的热带亚热带水果,为世界五大名果之一,为世界第二大热带水果。中国是世界主产国之一,2020年栽培面积34.34万hm2,产量约331.2万t(农业农村部发展南亚热带作物办公室统计数据)。果实品质是决定杧果市场竞争力的主要因素,品质好坏直接影响经济效益。可溶性糖和有机酸是杧果果实主要风味品质,其组分、含量及其比例是影响杧果果实食用品质的重要因素[1]。杧果果实品质性状研究主要集中在不同品种品质性状的变化上,而对其杂交群体遗传规律研究较少,原因可能是杧果大部分品种存在多胚现象,通过人工杂交难以获得一定数量的有性后代。
目前,品质性状遗传评价研究在苹果、葡萄、柑橘、枣等果树上报道比较多。普遍认为杂交后代果实中糖含量的遗传除存在加性效应外,还存在一定程度的非加性效应,且糖含量多呈正态分布,是由微效多基因控制的数量性状[2-4];而有机酸含量的遗传比较复杂,有机酸含量多呈偏态分布,推测是由主效基因和多个微效基因共同控制的数量性状[5-6];类胡萝卜素和总黄酮含量多呈正态分布,是由多基因控制的数量性状[7-8]。因此,研究杧果果实内在品质的遗传机制,对于杧果新品种的培育具有非常重要的作用。笔者在本研究中以果实糖酸含量存在较大差异的金煌×热农1 号杂交F1代47 个株系为研究对象,针对可溶性糖、有机酸、类胡萝卜素和总黄酮含量4 个品质性状,分析杂交后代果实性状的遗传特点和趋势,以期为杧果品质育种提供依据和参考。
1 材料和方法
1.1 试验材料
金煌和热农1 号及其杂交后代群体47 个单株均采自广东省湛江市中国热带农业科学院南亚热带作物研究所国家热带果树种质资源圃(杧果分圃),2012 年按2.0 m × 3.5 m株行距定植,立地条件及管理水平一致。采摘选择树冠外围不同方向色泽接近、大小一致、无病虫害的果实;5 个果实为1 个生物学重复,共3 次重复。完熟时将果实切成小块,液氮冷冻后于-80 ℃超低温冰箱保存备用。
1.2 试验方法
1.2.1 糖组分测定 样品制备参照刘胜辉等[9]的方法略有改进,准确称取1.0 g 果肉,用5 mL 80%乙醇研磨提取,4 ℃ 10 000 r · min-1 离心10 min,上清液转入10 mL离心管,残渣再次加入4 mL 80%乙醇继续离心,取上清液,90 ℃水浴蒸干,定容至10 mL,经0.45 μm水系滤膜过滤至样品瓶中,于-20 ℃保存待测。
利用LC-10ATVP 高效液相色谱仪,色谱柱为Prevail Carbohydrate ES 色谱柱(4.6 mm × 250 mm,5 μm),柱温40 ℃ ;流动相体积比为乙腈∶水=70∶30,流速为1 mL·min-1;进样量20 μL。所用标样为Sigma 公司提供的色谱纯蔗糖、果糖和葡萄糖,配成不同浓度的混标进行测定,绘制标准曲线。1.2.2 有机酸含量测定有机酸含量测定参考Nisperos-Carriedo 等[10]的方法。准确称取1.0 g 果肉加入5 mL 0.2%偏磷酸研磨提取,4 ℃ 10 000 r·min-1离心15 min,上清液转入10 mL离心管,残渣加入4 mL0.2% 偏磷酸再提取,合并上清液并定容至10 mL,经0.45 μm的水系滤膜过滤后装瓶待测。
酸组分测定使用仪器为DIONEX U3000 高效液相色谱,色谱柱为Agilent-C18 色谱柱(4.6 mm×250 mm,5 μm),流动相为0.2%偏磷酸,流速为1 mL·min-1,柱温30 ℃,进样量20 μL。所用标样为Sigma 公司提供的色谱纯抗坏血酸、苹果酸、乙酸、甲酸、柠檬酸、草酸、酒石酸,配成不同浓度的混标进行测定,绘制标准曲线。
1.2.3 类胡萝卜素含量测定 以V乙醇∶V丙酮∶V水=4.5∶4.5∶1 为提取液,准确称取样品0.3 g 于15 mL离心管中,加入10 mL提取液。封口膜封口,4 ℃过夜(12 h)避光保存。待样品变白色后,以提取液为空白对照,分别于波长663 nm、645 nm 和470 nm 处测定吸光度,3 次重复。按下列公式分别计算叶绿素a、b 和类胡萝卜素的质量浓度以及类胡萝卜素质量分数:叶绿素a 质量浓度(Ca,μg · mL- 1)=11.75×A662-2.35×A645;叶绿素b 质量浓度(Cb,μg ·mL-1)=18.61×A645-3.96×A662;类胡萝卜素质量浓度(μg ·mL-1)=(1000×A470-2.27×Ca-81.4×Cb)/227;类胡萝卜素质量分数(mg · g- 1)=c ×V/W/1000;式中c:色素质量浓度(μg ·mL-1);V:提取液体积(8 mL);W:鲜质量(g)。
1.2.4 总黄酮含量测定 总黄酮含量测定采用紫外-分光光度计法,参照刘常凯等[11]的方法,略有改进。标准线性的制备:称取12.60 mg芦丁对照品置于10 mL容量瓶中,加入甲醇溶解并稀释至刻度,摇匀,即得质量浓度为1.157 9 mg·mL-1的标准储备液,已按91.9%计算,再精确量取1.50 mL至10 mL容量瓶中,加甲醇溶液稀释至刻度,得到质量浓度为173.685 μg·mL-1的标准工作液,吸取0、0.5、1.0、2.0、3.0、4.0、5.0 mL芦丁标准溶液,分别置于10 mL的具塞比色管中,加入2.0 mL三氯化铝溶液(2.5 g·100 mL-1)混匀,标准管加水定容,静置,在30 min 内用1 cm比色皿,以零管调零,于波长415 nm处测定吸光度,以试剂空白作为参照。准确称取5 g 经过均质的杧果樣品,10 000 r · min-1离心10 min 后取上清液备用。吸取上述溶液1.0 mL于10 mL的具塞比色管中,分别加入2.0 mL三氯化铝溶液(2.5 g·100 mL-1)混匀,静置,在30 min 内用1 cm比色皿,以零管调零,于波长415 nm处测定吸光度,所得吸光度代入标准曲线中计算样品中总黄酮含量。
1.3 数据处理与分析
对糖酸等品质性状进行测定,参照董昕等[12]的方法,将品质性状进行分级,1 级:<平均值-2s,10级:≥平均值+2s;中间每级相差0.5s。s 为标准差。利用Microsoft Excel 2016 软件对数据进行处理分析,制作正态分布图,计算每个性状的遗传规律。变异系数(CV)/%=s/F × 100;遗传传递力(Ta)/%= F/MP × 100;优势率(Ha)/%=(F-MP)/MP × 100。式中s 为标准差,F 为F1代平均值,MP为亲本平均值(亲中值)。果实性状多样性用Shannon-Weaver 多样性指数(H)表示。
2 结果与分析
2.1 杂交后代果实糖酸组分、类胡萝卜素和总黄酮含量的遗传分析
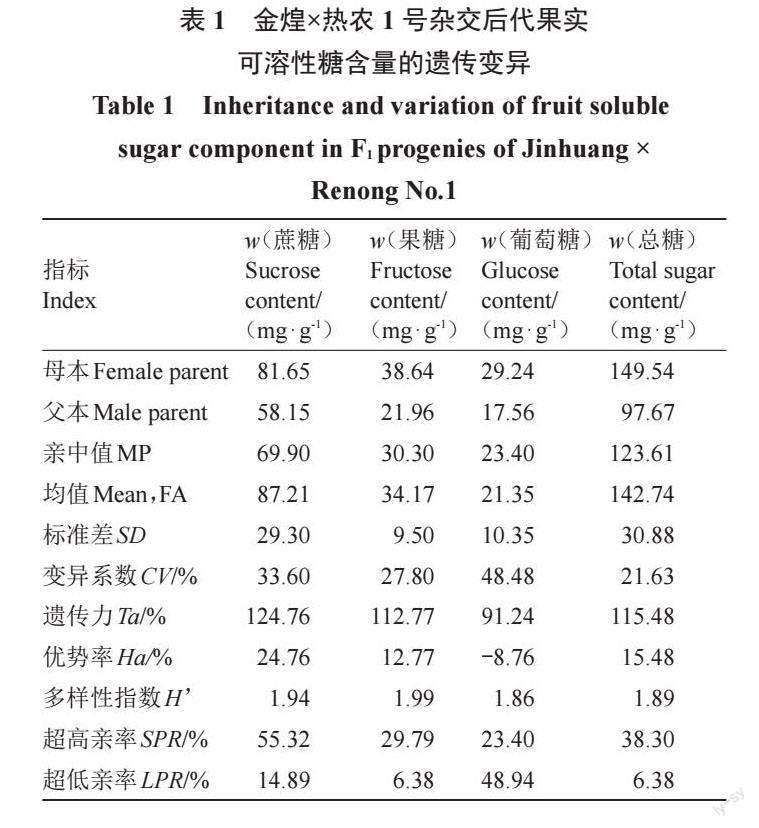
2.1.1 杂交后代果实糖组分的遗传分析杂交群体果实糖含量测定结果显示,后代果实中糖的主要成分与亲本一致,蔗糖是主要糖组分,其次为果糖,葡萄糖在成熟果实中相对含量低(表1)。金煌×热农1号杂交后代果实中可溶性糖含量分离广泛,变化范围跨度大,遗传变异分离现象明显。蔗糖、果糖、葡萄糖含量的变异范围分别为36.11~162.85、15.78~55.78 和8.57~51.05 mg·g-1。蔗糖和果糖含量平均值均高于亲中值,倾向于高糖亲本金煌,而葡萄糖含量平均值低于亲中值。杂交后代中蔗糖和果糖含量的超高亲率分别为55.32%、29.79%,且存在一定比例的超亲单株,说明蔗糖和果糖含量超亲遗传潜力大;葡萄糖含量的超低亲率为48.94%,且存在一定比例的低亲单株,说明葡萄糖含量低亲遗传潜力大。蔗糖、果糖、葡萄糖含量的遗传力分别为124.76%、112.77%和91.24%,变异系数(CV)分别为33.60%、27.80%和48.48%,Shannon-Weaver 多样性指数(H)分别为1.94、1.99 和1.86(表1),说明杂交后代果实糖组分多样性丰富,各物质均有较强的遗传传递力,在亲本与后代间可以稳定遗传。
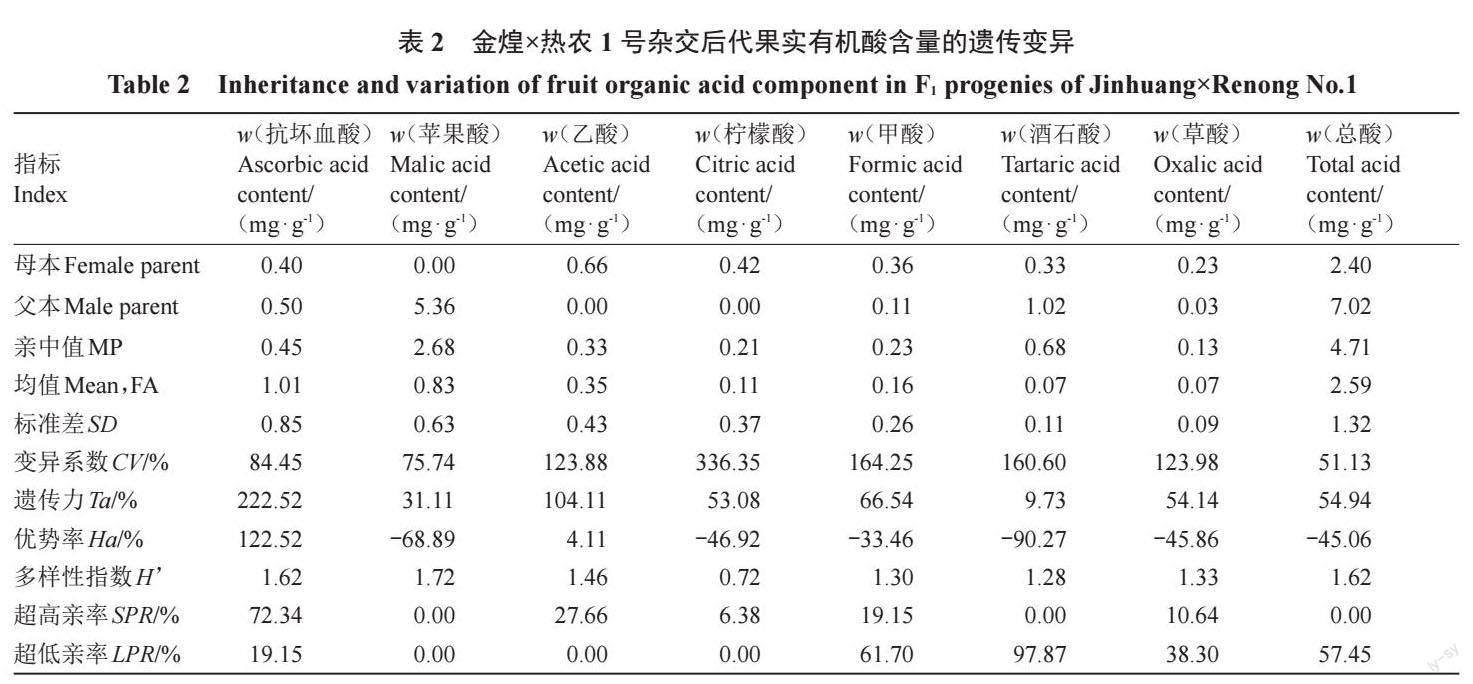
2.1.2 杂交后代果实酸组分的遗传分析 对亲本和杂交群体果实有机酸含量测定结果(表2)显示,亲本果实中苹果酸为主要酸组分,其次分别为酒石酸、抗坏血酸、乙酸、甲酸、柠檬酸、草酸。杂交后代果实中有机酸与亲本不同,以积累抗坏血酸为主,其次为苹果酸、乙酸、柠檬酸和甲酸,同时含有少量的草酸、酒石酸。杂交后代果实中有机酸含量分离广泛,变化范围跨度大,遗传变异分离现象明显。抗坏血酸、苹果酸、乙酸、柠檬酸、甲酸、酒石酸和草酸含量(w,后同)的分布范围分别为0.08~5.25、0.20~2.66、0.22~1.56、0.18~2.44、0.15~1.13、0.06~0.49 和0.005~0.35 mg·g-1。杂交后代中抗坏血酸和乙酸含量的平均值均高于亲中值,而苹果酸、柠檬酸、甲酸、酒石酸和草酸含量的平均值均低于亲中值,倾向于低酸亲本,后代中抗坏血酸、乙酸和柠檬酸含量的超高亲率分别为72.34%、27.66%和6.38%,且存在一定比例的高亲单株,说明抗坏血酸、乙酸和柠檬酸含量高亲遗传潜力大。苹果酸含量的超高亲率和超低亲率均为0,说明苹果酸含量的遗传潜力较小;甲酸、酒石酸和草酸含量的超低亲率分别为61.70%、97.87%和38.30%,且存在一定比例的低亲单株,说明甲酸、酒石酸和草酸含量低亲遗传潜力大。抗坏血酸、苹果酸、乙酸、柠檬酸、甲酸、酒石酸和草酸含量的遗传传递力分别为222.52%、31.11%、104.11%、53.08%、66.54%、9.73%、54.14%;变异系数(CV)范围分别为84.45% 、75.74% 、123.88% 、336.35% 、164.25% 、160.60%、123.98%,Shannon-Weaver多样性指数(H)分别为1.62、1.72、1.46、0.72、1.30、1.28、1.33(表2),说明杂交后代果实酸组分多样性丰富,抗坏血酸、乙酸、甲酸、柠檬酸和草酸含量均有较强的遗传传递力,在亲本与后代间可以稳定遗传,苹果酸和酒石酸含量遗传传递力较小,在亲本与后代间遗传不稳定。
2.1.3 杂交后代果实类胡萝卜素和总黄酮含量的遗传分析 金煌×热农1 号杂交后代果实中类胡萝卜素和总黄酮含量分离广泛,变化范围跨度大,遗传变异分离现象明显(表3)。其中类胡萝卜素含量的分布范围是0.001~0.073 mg·g-1;总黄酮含量的分布范围是0.004~0.019 mg· g-1。杂交后代果实类胡萝卜素含量平均值高于亲中值,而总黄酮含量平均值低于亲中值。后代中类胡萝卜素含量的超高亲率为65.96%,且存在一定比例的超亲单株,说明类胡萝卜素含量超亲遗传潜力大;总黄酮含量的超低亲率为53.19%,且存在一定比例的低亲单株,说明总黄酮含量低亲遗传潜力大。类胡萝卜素和总黄酮含量的遗传力范围分别为150.93%和60.00%,变异系数(CV)范围分别为51.68%和40.25%,Shannon-Weaver多样性指数(H)分别为2.03和1.88(表3),说明杂交后代果实类胡萝卜素和总黄酮含量多样性丰富,遗传传递力较强,在亲本与后代间可以稳定遗传。
2.2 杂交后代果实糖酸组分、类胡萝卜素和总黄酮含量的分布2
.2.1 糖组分含量的分布 由图1可知,蔗糖、果糖、葡萄糖和总糖含量的分布在杂交后代中基本符合正态分布,具有典型的数量性状遗传特征,推测杧果可溶性糖含量属于微效多基因控制的数量性状。蔗糖含量最低为43.26 mg·g-1,最高为160.44 mg·g-1,主要分布在72.56~131.16 mg·g-1之间,系中等甜度,共38株,占比80.85%;其次分布在43.26~57.91 mg· g-1之间,有7 株,蔗糖含量偏低,占比14.89%;蔗糖含量在145.80 mg·g-1以上的有2株,占比4.26%。果糖含量最低为19.92 mg·g-1,最高为53.18 mg·g-1,主要分布在24.67~43.68 mg· g-1之间,系中等甜度,共39 株,占比82.98%;其次分布在48.43~57.93 之间,有7 株,果糖含量较高,占比14.89%;果糖含量低于24.67 mg· g-1的杂交后代仅有1 株,含量较低,占比2.13%。
葡萄糖含量最低为11.00 mg · g- 1,最高为47.24 mg·g-1,主要分布在11.00~31.70 mg·g-1之间,系中等甜度,共40 株,占比85.11%;其次分布在36.88~47.24mg·g-1之间,有7株,葡萄糖含量较高,占比14.89%。总糖含量最低为80.98mg·g-1,最高为219.94mg·g-1,主要分布在111.86~189.06 mg· g-1之间,共42 株,占比89.36%;其次分布在80.98~96.42 mg·g-1之间,有3株,含量偏低,占比6.38%;总糖含量高于189.06 mg·g-1的杂交后代有2 株,含量较高,占比4.26%。
2.2.2 酸组分含量的分布 由圖2 可知,苹果酸、乙酸、甲酸、酒石酸含量都呈连续变异,频率分布呈偏态分布,属于微效多基因控制的数量性状;抗坏血酸、柠檬酸、草酸和总酸含量都呈不连续变异,频率分布呈偏态分布,推测抗坏血酸、柠檬酸和草酸含量的遗传有可能是由主效基因和多个微效基因共同控制的复杂遗传。抗坏血酸含量主要分布在0.16~2.29 mg·g-1之间,有46 株,占比97.88%,高于2.71 mg· g-1的只有1 株,占2.12%;苹果酸含量主要分布在0.20~1.46 mg· g-1,有41 株,占比87.23%,1.78~2.42 mg·g-1有6 株,占比12.77%;乙酸含量主要分布在0.13~0.99 mg·g-1,有43 株,占比91.49%,1.20~1.41 mg·g-1的有3 株,占6.39%;柠檬酸含量主要分布在0.11~0.67 mg·g-1,有46 株,占比97.88%,高于0.86 mg·g-1只有1 株,占比2.12%;甲酸含量主要分布在0.03~0.28 mg·g-1,有36 株,占比76.60%,0.41~0.80 mg·g-1的有11株,占比23.40%;酒石酸含量主要分布在0.01~0.17 mg·g-1,有41 株,占比87.23%,0.22~0.34 mg·g-1的有6 株,占比12.77%;草酸含量主要分布在0.03~0.11 mg·g-1,有41 株,占比87.23%,0.20~0.28 mg·g-1的有6 株,占比12.77%。总酸含量主要分布在1.26~3.91 mg·g-1,有38 株,占比80.85%,4.57~5.89 mg·g-1的有9 株,占比19.15%。
2.2.3 类胡萝卜素和总黄酮含量的分布 由图3 可知,F1果实类胡萝卜素和总黄酮含量整体上都呈连续变异,属于微效多基因控制的数量性状。类胡萝卜素含量主要分布在0.007~0.049 mg·g-1,有41株,占比87.23%,其次分布在0.057~0.073 mg·g-1,有6株,占比12.77%;总黄酮含量主要分布在0.008~0.013 mg·g-1,有37株,占比78.72%,其次分布在0.004~0.006 mg·g-1,有5 株,占比10.64%;高于0.016 7 mg· g-1的有5 株,占比10.64%。
3 讨论
果树杂交育种后代性状多呈现广泛分离,劣变率往往很高,但也会出现超高亲植株[13]。Sharma[14]对印度农业研究所培育的2个优良杂种Mallika和Amrapali及其亲本Dashehari和Neelum的β-类胡萝卜素和可溶性固形物含量的遗传进行研究,发现2个杂种的β-类胡萝卜素和可溶性固形物含量均超过了亲本中较高的一方,表明这是加性基因的作用或超亲分离的现象。Uddin 等[15]对杂交后代株系的农艺性状进行研究,发现杂交后代中存在高可溶性固形物含量的株系,最高可达27%,这些结果均证明可溶性固形物含量等果实品质性状存在超亲分离的现象。笔者前期研究发现,不同杧果品种果实品质性状丰富多样,杧果果实的可滴定酸、糖含量都存在较大的变异系数[16];金煌杧可溶性固形物和可溶性糖含量较高,而热农1 号属于典型的低可溶性固形物和可溶性糖含量的品种,且两个品种亲缘关系较远[16],这为培育具有合适糖酸度比例的高品质杧果商业品种提供了材料保障。本研究中,金煌×热农1 号杂交后代果实除葡萄糖含量的平均值略低于亲中值外,其余糖组分的平均值均高于亲中值,杂交后代果实中糖含量倾向于高糖亲本,超亲优势明显,说明果实中糖含量的遗传主要受加性效应的影响。这与前人在柑橘、葡萄、枣等果树中的研究结果有区别[4,6,8,17-18]。有机酸方面,杂交后代果实中的苹果酸、柠檬酸、甲酸、酒石酸、草酸和总酸含量平均值低于亲中值,且50%以上的单株总酸含量低于低值亲本,表现为超低亲遗传,遗传表现为基因的加性效应;而抗坏血酸和乙酸含量平均值高于亲中值,表明抗坏血酸和乙酸的遗传存在加性效应和非加性效应,通过杂交的方式选育高抗坏血酸和乙酸的株系或品种具有很大优势。杂交后代果实中类胡萝卜素含量平均值高于亲中值,表现出超高亲遗传,说明性状遗传以加性效应为主,并且存在正向的非加性效应[19];总黄酮含量平均值低于亲中值,表现出超低亲遗传倾向,受加性效应的影响,这与连红娟[20]、谢欢等[8]研究结果有区别。本研究结果将为下一步利用重测序技术,定位杧果糖酸性状的QTL位点,挖掘杧果糖酸性状的功能基因,筛选与糖酸性状紧密连锁的标记奠定基础,从而为分子辅助育种和筛选亲本提供前期基础和科学依据。
前人对越橘[2]、柑橘[4]、苹果[5]、葡萄[6,15]、梨[21]、枣[22]等果树研究发现,果实糖含量一般为多基因控制的数量性状,遗传效应以加性效应为主。本研究中杂交后代群体果实蔗糖、果糖、葡萄糖和总糖含量均呈正态分布,表明果实可溶性糖含量的遗传模式与已报道的其他果树相似,推测也属于微效多基因控制的数量性状遗传。与可溶性糖含量相比,前人发现有机酸含量的遗传比较复杂,多呈偏态分布,推测有机酸含量是由1 个或多个主效基因控制的数量性状[4-5,23-26]。在本研究中,杂交后代群体果实有机酸含量均呈偏态分布,表明果实有机酸含量可能是由1 个或多个主效基因控制的数量性状。果实类胡萝卜素和总黄酮含量基本呈正态分布,属于多基因控制的数量性状,这与前人在枣、柑橘等果树中的研究结果基本一致[7-8,19,27]。
4 结论
以金煌为母本、热农1 号为父本进行杂交获得的杂交后代群体果实中,蔗糖、果糖和葡萄糖是主要糖组分,抗坏血酸、苹果酸、乙酸、柠檬酸和甲酸是主要酸组分,同时含有少量的草酸、酒石酸。糖酸含量分布广泛,糖含量呈正态分布,推测属于多基因控制的数量性状,遗传潜能大,其含量趋向高糖亲本,具有母性遗传倾向。酸含量呈偏态分布,抗坏血酸和乙酸的遗传潜能大,而其他的酸遗传潜能略小,含量趋向母本,明显受母本影响,推测可能是由1 个或多个主效基因控制的数量性状。类胡萝卜素含量呈正态分布,含量倾向父本,具有父性遗传倾向,属于多基因控制的数量性状。总黄酮含量呈正态分布,含量倾向母本,明顯受母本影响,属于多基因控制的数量性状。研究表明杧果杂交后代果实糖酸含量和总黄酮含量受母本影响较大,类胡萝卜素含量受父本影响较大,这将为今后科学配置杂交组合、培育出品质优良的杧果提供理论依据。
参考文献References:
[1] 石胜友,马小卫,许文天,周毅刚,武红霞,姚全胜,罗纯,王松标. 不同芒果种质果实品质性状多样性分析[J]. 热带作物学报,2014,35(11):2168-2172.
SHI Shengyou,MA Xiaowei,XU Wentian,ZHOU Yigang,WUHongxia,YAO Quansheng,LUO Chun,WANG Songbiao. Fruitquality characters of various mango germplasms[J]. ChineseJournal of Tropical Crops,2014,35(11):2168-2172.
[2] 刘有春,魏永祥,王兴东,刘成,蒋明三,张舵,袁兴福,陶承光.南高丛越橘品种‘Sapphire和北高丛品种‘Berkeley正反交后代果实糖酸组分含量的遗传倾向[J]. 中国农业科学,2014,47(24):4878-4885.
LIU Youchun,WEI Yongxiang,WANG Xingdong,LIU Cheng,JIANG Mingsan,ZHANG Duo,YUAN Xingfu,TAO Chengguang.Inheritance tendency of sugar and acid contents in the reciprocalcross progeniesfruits of southern × northern high bushblueberry (Vaccinium) [J]. Scientia Agricultura Sinica,2014,47(24):4878-4885.
[3] LIU L,CHEN C X,ZHU Y F,XUE L,LIU Q W,QI K J,ZHANG S L,WU J. Maternal inheritance has impact on organicacid content in progeny of pear (Pyrus spp. ) fruit[J]. Euphytica,2016,209(2):305-321.
[4] 王婷婷,周阳广,朱虹娴,张苗,段耀园,曹惠祥,管书萍,解凯东,伍小萌,龙春瑞,高俊燕,郭文武. 2 个柑橘三倍体有性群体果实糖酸性状遗传评价[J]. 果树学报,2022,39(7):1147-1156.
WANG Tingting,ZHOU Yangguang,ZHU Hongxian,ZHANGMiao,DUAN Yaoyuan,CAO Huixiang,GUAN Shuping,XIEKaidong,WU Xiaomeng,LONG Chunrui,GAO Junyan,GUOWenwu. Inheritance of sugar and acid contents in the fruits oftriploid hybrids originated from two 2x × 4x crosses with Nadorcotttangor as a female parent[J]. Journal of Fruit Science,2022,39(7):1147-1156.
[5] 闫忠业,伊凯,刘志,王冬梅,吕天星,李春敏,陈东玫.‘红玉ב金冠苹果杂交后代果实糖酸组分遗传分析[J]. 果树学报,2017,34(2):129-136.
YAN Zhongye,YI Kai,LIU Zhi,WANG Dongmei,L? Tianxing,LI Chunmin,CHEN Dongmei. A study of genetic trend of sugarand acid components in the fruits of apple hybrid progeny of‘JonathanבGolden Delicious[J]. Journal of Fruit Science,2017,34(2):129-136.
[6] 郭权,郭印山,郭修武. 葡萄‘红地球与‘双优杂交后代果实糖酸的遗传规律[J]. 分子植物育种,2021,19(10):3424-3431.
GUO Quan,GUO Yinshan,GUO Xiuwu. Genetic law of sugarand acid in the fruits of hybrids between‘Red Globeand‘Shuangyou[J]. Molecular Plant Breeding,2021,19(10):3424-3431.
[7] 汤雨晴. 柑橘果肉色泽的遗传研究及类胡萝卜素含量的QTL定位[D]. 武汉:华中农业大学,2018.
TANG Yuqing. The genetic studies of pulp color and mapping ofQTL for carotenoids content in citrus[D]. Wuhan:Huazhong AgriculturalUniversity,2018.
[8] 谢欢,王中堂,李明玥,李新岗. 枣杂交后代果实性状遗传分析[J]. 经济林研究,2022,40(2):125-134.
XIE Huan,WANG Zhongtang,LI Mingyue,LI Xingang. Geneticanalysis of fruit characters in hybrid progeny of Chinese jujube[J]. Non-wood Forest Research,2022,40(2):125-134.
[9] 劉胜辉,魏长宾,孙光明. 高效液相色谱法测定台农19 号菠萝糖分[J]. 广东农业科学,2008,35(12):133-134.
LIU Shenghui,WEI Changbin,SUN Guangming. Determinationof carbohydrates in Tainung 19 pineapple by HPLC[J].Guangdong Agricultural Sciences,2008,35(12):133-134.
[10] NISPEROS-CARRIEDO M O,BUSLIG B S,SHAW P E. Simultaneousdetection of dehydroascorbic,ascorbic,and some organicacids in fruits and vegetables by HPLC[J]. Journal of Agriculturaland Food Chemistry,1992,40(7):1127-1130.
[11] 刘常凯,杨黎,何林飞. 芒果果肉中总黄酮及芒果苷含量的测定[J]. 湖北农业科学,2020,59(8):138-140.
LIU Changkai,YANG Li,HE Linfei. Determination of total flavonoidsand mangiferin in mango flesh[J]. Hubei AgriculturalSciences,2020,59(8):138-140.
[12] 董昕,李淑君,杨华,官玲,付忠军,祁志云,金川,余雪源,易红华,陈荣丽,张丕辉. 重庆玉米地方品种表型多样性分析[J].植物遗传资源学报,2019,20(4):861-870.
DONG Xin,LI Shujun,YANG Hua,GUAN Ling,FU Zhongjun,QI Zhiyun,JIN Chuan,YU Xueyuan,YI Honghua,CHENRongli,ZHANG Pihui. Phenotypic diversity of maize landracescollected from Chongqing[J]. Journal of Plant Genetic Resources,2019,20(4):861-870.
[13] LABROOMR,STUDERAJ,RUTKOSKI J E. Heterosis and hybridcrop breeding:Amultidisciplinary review[J]. Frontiers in Genetics,2021,12:643761.
[14] SHARMA D K. Mango Breeding[J]. Acta Horticulturae,1987,196:61-67.
[15] UDDIN M Z,ISLAM M S,MORTUZAM G,BHUYAN M,ALAMINM. Studies on physico-morphological characteristics andyield of some mango hybrid lines[J]. Journal of Agroforestryand Environment,2011,5(1):131-134.
[16] 马蔚红,谢江辉,武红霞,王松标. 杧果种质资源果实数量性状评价指标探讨[J]. 果树学报,2006,23(2):218-222.
MA Weihong,XIE Jianghui,WU Hongxia,WANG Songbiao.Evaluating criteria of some fruit quantitative characteristics ofmango (Mangifera indica Linn.) germplasm resources[J]. Journalof Fruit Science,2006,23(2):218-222.
[17] 刘家成,章秋平,牛铁泉,刘宁,张玉萍,徐铭,马小雪,张玉君,刘硕,刘威生.‘串枝红与‘赛买提杏正、反交后代果实性状遗传倾向分析[J]. 果树学报,2020,37(5):625-634.
LIU Jiacheng,ZHANG Qiuping,NIU Tiequan,LIU Ning,ZHANG Yuping,XU Ming,MA Xiaoxue,ZHANG Yujun,LIUShuo,LIU Weisheng. Analysis of inherited tendency of fruitcharacteristics in F1 group of reciprocal crossing between‘Chuanzhihongand‘Saimaitiin apricots[J]. Journal of Fruit Science,2020,37(5):625-634.
[18] 刘政海,董志刚,李晓梅,谭敏,杨镕兆,杨兆亮,唐晓萍.‘威代尔与‘霞多丽葡萄杂交F1 代果实性状遗传倾向分析[J].果树学报,2020,37(8):1122-1131.
LIU Zhenghai,DONG Zhigang,LI Xiaomei,TAN Min,YANGRongzhao,YANG Zhaoliang,TANG Xiaoping. Inheritancetrend of fruit traits in F1 progenies of‘Vidaland‘Chardonnayof grape[J]. Journal of Fruit Science,2020,37(8):1122-1131.
[19] 赵亚楠,欧春青,王斐,张艳杰,马力,姜淑苓. 梨F1 代群体果实性状的遗传分析及相关性研究[J]. 西北农业学报,2018,27(12):1811-1818.
ZHAO Yanan,OU Chunqing,WANG Fei,ZHANG Yanjie,MALi,JIANG Shuling. Genetic analysis and correlation study onpear fruit characteristics in F1 generation[J]. Acta AgriculturaeBoreali-Occidentalis Sinica,2018,27(12):1811-1818.
[20] 連红娟.‘赤霞珠和‘西拉正反交后代果实主要性状遗传倾向研究[D]. 太原:山西农业大学,2020.
LIAN Hongjuan. Genetic tendency of main fruit characters in reciprocalcrosses hybrid progeny of‘Cabernet Sauvignonand‘Shiraz[D]. Taiyuan:Shanxi Agricultural University,2020.
[21] 崔艳波,陈慧,乐文全,张树军,伍涛,陶书田,张绍铃.‘京白梨与‘鸭梨正反交后代果实性状遗传倾向研究[J]. 园艺学报,2011,38(2):215-224.
CUI Yanbo,CHEN Hui,LE Wenquan,ZHANG Shujun,WUTao,TAO Shutian,ZHANG Shaoling. Studies on genetic tendencyof fruit characters in reciprocal crosses generation between‘Jingbailiand‘Yalipear cultivars[J]. Acta Horticulturae Sinica,2011,38(2):215-224.
[22] 夏怡蕾.‘JMS2ב邢16杂交后代糖酸性状遗传规律及品质性状QTL 定位[D]. 阿拉尔:塔里木大学,2022.
XIA Yilei. Genetic law of sugar and acid traits and QTL mappingof quality traits in Chinese jujube hybrid progenies [D].Alar:Tarim University,2022.[23] 李宝江,景士西,丁玉英,张景娥. 苹果糖酸遗传和选择研究[J].遗传学报,1994,21(2):147-154.LI Baojiang,JING Shixi,DING Yuying,ZHANG Jinge. Studiesof the inheritance and selection of sweetness and acidity inapples[J]. Acta Genetica Sinica,1994,21(2):147-154.[24] 李俊才,伊凯,刘成,隋洪涛,王家珍. 梨果实部分性状遗传倾向研究[J]. 果树学报,2002,19(2):87-93.LI Juncai,YI Kai,LIU Cheng,SUI Hongtao,WANG Jiazhen.Studies on the trend of inheritance of some characters of pearfruit[J]. Journal of Fruit Science,2002,19(2):87-93.[25] 王宏伟,王成荣,于淼,戴洪义,王然. 梨杂交后代果实主要有机酸遗传动态的研究[J]. 青岛农业大学学报(自然科学版),2008,25(3):231-235.WANG Hongwei,WANG Chengrong,YU Miao,DAI Hongyi,WANG Ran. Genetic tendency of organic acid in the pear hybridprogeny[J]. Journal of Qingdao Agricultural University (NaturalScience),2008,25(3):231-235.[26] LIAO L,ZHANG W H,ZHANG B,HAN Y P. Unraveling a geneticroadmap for improved taste in the domesticated apple[J].Molecular Plant,2021,14(9):1454-1471.[27] 夏怡蕾,仇倩倩,杨植,鲍荆凯,闫芬芬,王玖瑞,吴翠云,刘孟军. 枣果实总黄酮和总酚含量性状的QTL 定位分析[J/OL].分子植物育种:1-16[2022-09-02]. http://kns.cnki.net/kcms/detail/46.1068.S.20220505.1649.024.html.XIA Yilei,QIU Qianqian,YANG Zhi,BAO Jingkai,YAN Fenfen,WANG Jiurui,WU Cuiyun,LIU Mengjun. The QTL mappinganalysis of fruit total flavonoids and phenols in jujube[J/OL]. Molecular Plant Breeding:1- 16[2022- 09- 02]. http://kns.cnki.net/kcms/detail/46.1068.S.20220505.1649.024.html.
