生育后期干旱胁迫对玉米碳氮分配及生理特征的影响
2023-06-02孙焕烜尚宏越郭银银董志强李亚君
孙焕烜,尚宏越,郭银银,张 佩,董志强,李亚君
(西北农林科技大学 农学院,陕西杨凌 712100)
碳、氮代谢是作物最基本的两大代谢过程,二者相互影响,协调统一[1]。以光合作用为核心的碳代谢为作物生长发育提供最基本的结构和能量来源,也为氨基酸等含氮化合物合成提供物质基础。氮代谢为碳代谢提供酶和光合色素,作物氮素营养状况是影响碳同化效率的重要因素[2-3]。干旱胁迫会引起农作物碳、氮代谢失衡[4]。在营养生长阶段,干旱胁迫下光合速率下降,碳水化合物供应不足,氮素吸收受限,作物生长发育不良[5]。在生育后期,干旱胁迫可能会导致植株早衰、籽粒早熟,碳、氮同化物在源库间的转运和分配发生改变[6]。
作物生育后期自然衰老过程中伴随着养分由营养器官向生殖器官的转移和重新分配,这是其高效利用养分的重要生存策略之一[7]。干旱胁迫会改变这一进程,而碳、氮代谢等相关途径参与其中多方面的调控作用[2,4]。在生理水平上,干旱胁迫下光合机能下降,营养器官碳水化合物合成受阻,过剩的光能可能导致活性氧的大量积累,加速衰老进程,导致植株不能维持正常的形态及生理功能[8]。氮素养分状况是影响植株衰老的重要因素之一,营养器官较高的氮素水平有利于干旱胁迫下光合机能的维持,延缓衰老,提高耐旱性[9-10]。在代谢水平上,一些碳、氮代谢产物,如可溶性糖、糖醇类、脯氨酸、谷胱甘肽、多胺等是干旱胁迫下重要的渗透调节物质、抗氧化物质和信号分子等,参与作物抵御干旱胁迫的多种适应途径[8,11-12]。
植物碳代谢过程中,光合产物以蔗糖的形式输出并转运,其后在库器官中转化为淀粉等储存,蔗糖和淀粉相互转化受碳代谢关键酶的调节[13]。其中蔗糖合成酶主要参与催化将蔗糖转化为UDPG和果糖,可溶性淀粉合成酶则主要参与支链淀粉的合成[14-15]。谷氨酰胺合成酶是谷氨酰胺合成的关键酶,在植物氮素吸收及同化过程中具有重要作用。在植物生育后期,谷氨酰胺是营养器官向生殖器官氮素转运的重要载体,因而谷氨酰胺合成酶也是生育后期氮素转运的关键[2]。
生育后期干旱胁迫会严重影响玉米籽粒发育,导致产量下降[16]。目前,尽管已有部分研究对干旱胁迫下玉米碳、氮同化物积累及分配特征进行了探讨,但对于其生理及代谢调节机制仍较少报道。基于此,本研究从碳、氮同化物分配角度出发,通过测定碳、氮代谢关键酶活性及转录组测序分析,探讨干旱胁迫对玉米生育后期物质代谢的影响特征。为解析干旱胁迫下的代谢调节机制提供参考。
1 材料与方法
1.1 试验设计
试验在陕西省杨凌示范区西北农林科技大学防雨棚中池栽进行,棚中设2 m×1.5 m的水泥池小区,各池间筑水泥墙防止测渗。池内土壤为娄土,除水分处理外,池栽玉米各项栽培措施与常规田间管理一致。所用玉米品种为‘郑单958’,吐丝期以前所有处理均充分供水,吐丝期以后,使用TDR法监测并控制土壤供水量,对照为正常供水处理(Well watered,WW) ,保持土壤相对含水量在75%以上。干旱胁迫处理(Drought stress,DS) 土壤相对含水量为45%~55%至取样,每处理3个重复小区。
1.2 项目测定方法
1.2.1 生物量,碳、氮同化物含量及分配 分别于玉米成熟期取整株地上部分(不包括雄穗),按叶片、茎秆、穗轴和籽粒分开,烘干用测定各部位(茎秆、叶片、穗轴、苞叶、籽粒等)生物量、可溶性糖、蔗糖、淀粉、全氮等碳、氮同化物含量。其中生物量为各部位干质量,可溶性糖、蔗糖、淀粉含量的测定参照高俊凤的方法[17],氮含量(全氮)的测定使用凯氏定氮法[18]。相关参数计算如下:
各部位(茎秆、叶片、穗轴、苞叶、籽粒)碳、氮同化物积累量 = 干质量×碳、氮同化物含量
碳、氮同化物分配比例(%) =(碳、氮同化物积累量/地上部分总积累量)× 100%
1.2.2 碳氮代谢关键酶活性 生育后期(吐丝后50 d)分别取叶片(取样部位为穗位叶)和籽粒鲜样。用于测定碳氮代谢关键酶活性及基因表达分析。其中蔗糖合成酶(sucrose synthase,SS)活性(合成方向活性)的测定使用UDPG反应法进行[17]。可溶性淀粉合成酶(soluble starch synthase,SSS) 和谷氨酰胺合成酶(GS) 活性的测定参照Zhang等[19]的测定方法进行。
1.2.3 叶片转录组测序分析 对干旱胁迫(DS)与对照(WW)叶片进行转录组测序,分析干旱胁迫对生育后期叶片差异基因表达的影响。方法如下:分别取干旱胁迫(DS)和对照(WW)玉米叶片鲜样用于RNA提取和cDNA文库制备。总RNA的提取使用TRIzol试剂完成,使用DNase Ⅰ消化基因组DNA,纯化后的RNA用于构建cDNA文库,每处理3重复。构建好的cDNA文库使用Illumina平台进行转录组测序,经测序获得的高质量序列与玉米参考基因组比对并进行基因差异基因筛选及分析,其中|log2fold-change(FC)|≥1且false discovery rate <0.05判定为差异基因。对差异表达基因的代谢途径分析使用Mapman完成[20]。
1.3 数据分析
数据统计分析使用R软件(Version 3.6.1)完成,数据显著性检验采用单因素方差分析(ANOVA)法完成。图表的制作使用Microsoft office 2013完成。
2 结果与分析
2.1 干旱胁迫下成熟期各部分生物量及碳氮同化物分配状况
如图1所示,生育后期干旱胁迫处理显著降低了成熟期地上各部分生物量积累,其中整株生物量和穗粒质量分别降低28.3%和32.5%,可见干旱胁迫显著抑制了产量的形成,对穗轴和苞叶生物量的影响较小。在碳水化合物含量方面(表1),干旱胁迫下玉米茎秆、叶片、穗轴中可溶性糖含量与正常供水处理相比均显著降低,降低幅度分别为21.5%、10.7%、7.3%,籽粒可溶性糖含量在两处理间无显著差异。干旱胁迫下茎秆、叶片蔗糖含量分别显著降低29.8%、18.6%。干旱胁迫显著降低了成熟期籽粒淀粉含量,幅度为 2.6%,而茎秆和叶片中淀粉含量则分别增加41%和26.3%。在氮含量方面,干旱胁迫下玉米叶片和籽粒氮含量分别降低29.7%和4%,而穗轴中氮含量则显著增加。

表1 两种水分处理下玉米不同部位碳、氮同化物的含量Table 1 Carbohydrate,nitrogen content among different parts under two water treatments

不同小写字母表示两种水分处理在P<0.05水平下差异显著,下同
表2给出了生育后期干旱胁迫对成熟期各部分生物量及碳、氮同化物分配比例的影响。干旱胁迫下生物量在各部位的分配比例显著改变,表现为生物量在茎秆中的分配比例升高3.1%,而在籽粒中的分配比例则下降6.1%,可见干旱胁迫降低了玉米籽粒生物量分配比例(收获指数)。在碳水化合物分配方面,干旱胁迫对玉米茎秆、叶片中可溶性糖分配无明显影响,籽粒可溶性糖分配比例下降4.0%,穗轴和苞叶中的分配比例则分别提高2.5%和0.8%。干旱胁迫下茎秆、叶片蔗糖分配比例在处理间无显著差异。成熟期淀粉大量分配于籽粒中,干旱胁迫下籽粒淀粉分配比例有所下降,而茎秆和叶片中的分配比例则分别增加0.67%和0.36%。在氮素分配方面。干旱胁迫下叶片氮素分配比例下降3%,籽粒氮分配比例(氮收获指数)下降2.7%,而茎秆、穗轴中氮素分配比例则显著增加。可见干旱胁迫下生育后期氮素由营养器官向籽粒的分配受到了抑制,更多的氮素分配于茎秆等部位。
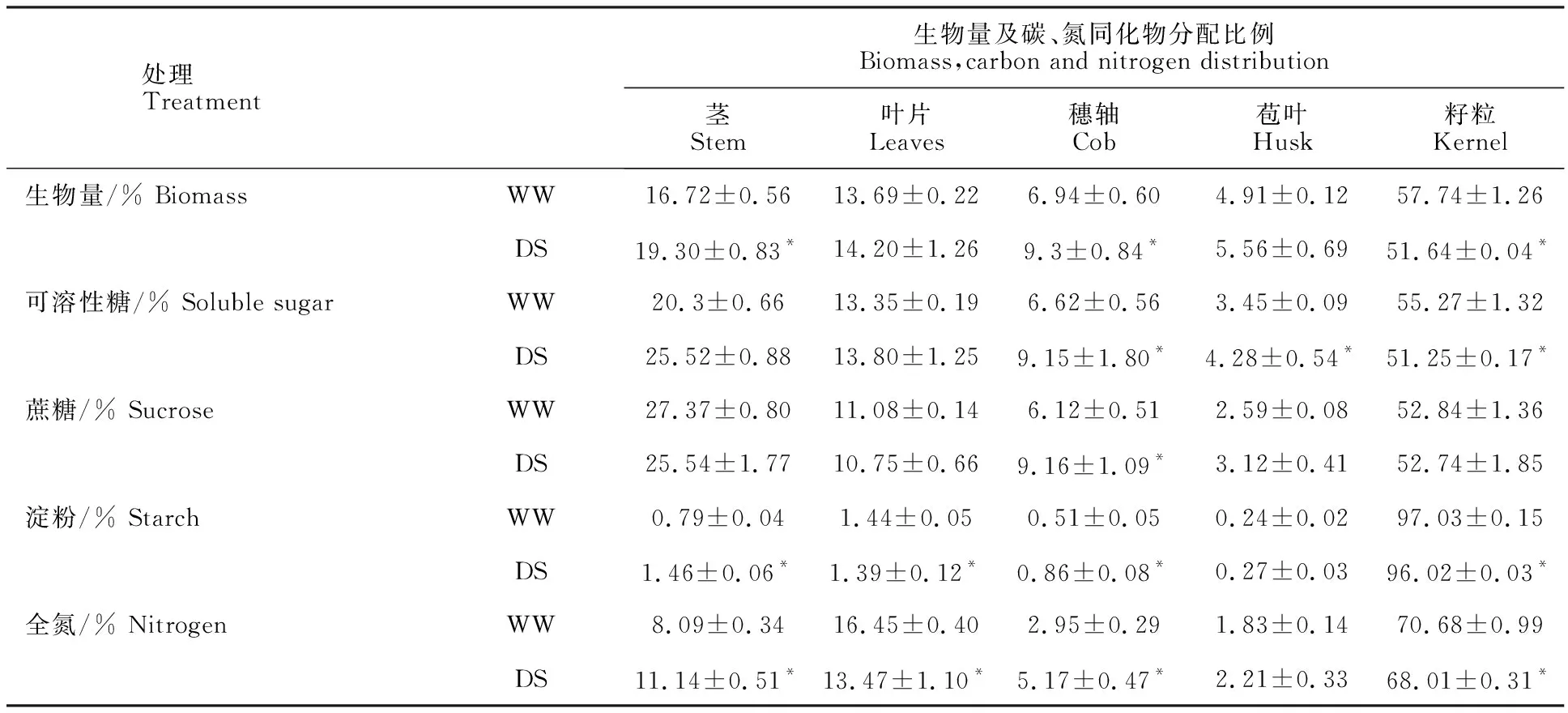
表2 两种水分处理下不同部位生物量及碳、氮同化物分配的比例Table 2 Biomass,carbohydrate,nitrogen distribution rate among different parts under two water treatments
2.2 干旱胁迫下碳氮代谢关键酶活性
对生育后期(吐丝后50 d)不同部位碳、氮代谢关键酶活性进行测定,结果如图2所示。干旱胁迫与对照相比叶片及籽粒中蔗糖合成酶活性均无明显差异。干旱胁迫下叶片可溶性淀粉合成酶活性显著高于正常供水处理,而籽粒中则显著降低。干旱胁迫处理显著提高吐丝后50 d叶片谷氨酰胺合成酶(GS)活性,两种水分处理对籽粒中的GS活性影响较小。
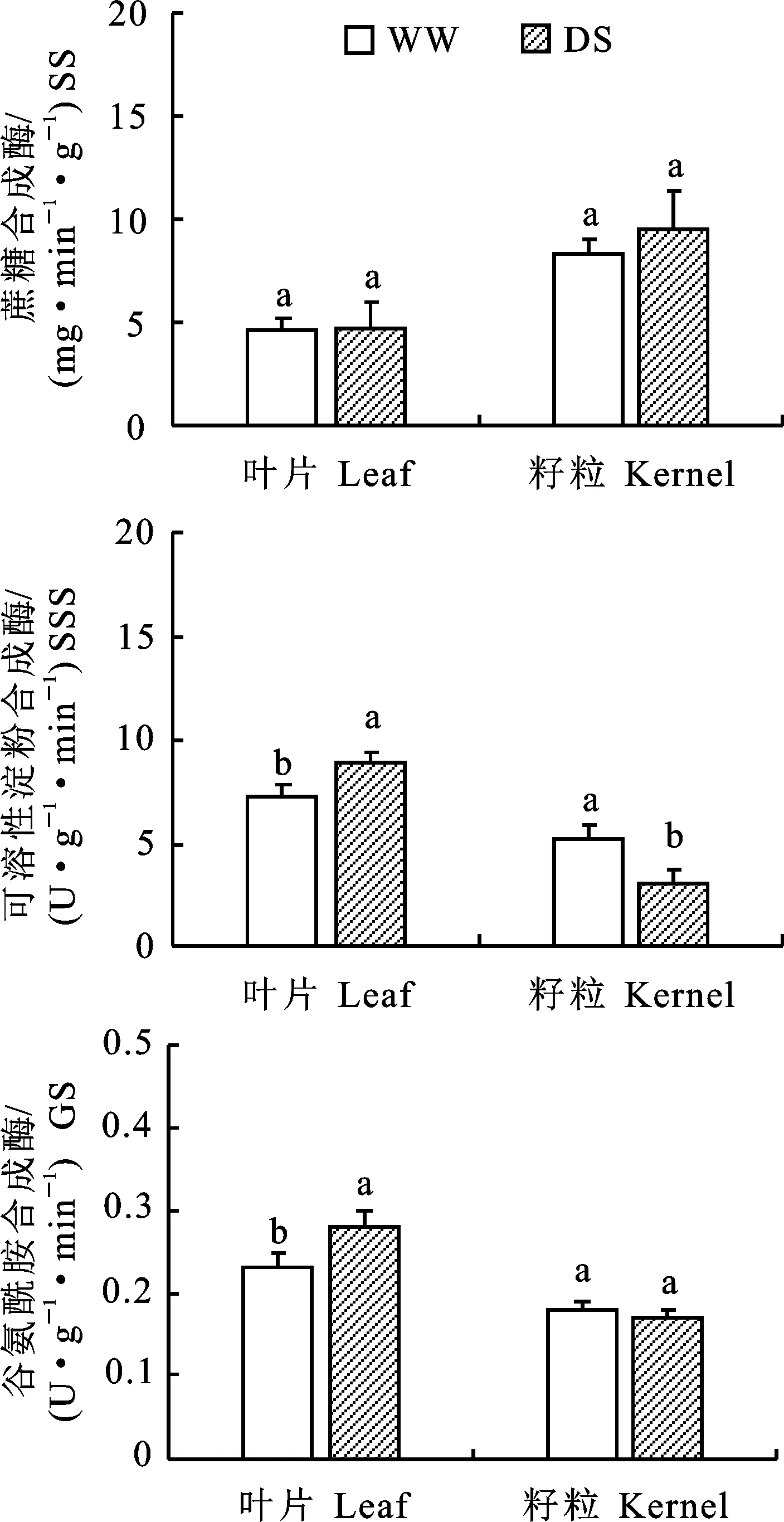
图2 两种水分处理对生育后期叶片和籽粒中蔗糖合成酶、可溶性淀粉合成酶和谷氨酰胺合成酶活性的影响Fig.2 SS,SSS and GS activity in leaves and grains at later growth stage under two water treatments
2.3 干旱胁迫对玉米叶片差异基因表达分析
比较干旱胁迫与对照叶片的转录组基因表达谱差异,筛选差异表达基因(DEGs)并进行代谢途径分析,结果表明:与对照(CK)相比,干旱胁迫下共检测到51个DEGs,其中上调表达28个,下调表达22个(图3-A)。对上述DEGs的代谢途径分析结果表明,部分差异基因分布在一些碳、氮及相关代谢途径(图3-B),例如细胞壁组织(Cell wall organisation)、碳水化合物代谢(Carbohydrate metabolism)、光合作用(Photosynthesis)、氨基酸代谢(Amino acid metabolism)、蛋白质合成(Protein biosynthesis)等。

图3 干旱胁迫下差异基因统计(A)及代谢途径分析(B)Fig.3 Statistics of DEGs(A) and metabolic pathway classification(B)under drought stress
对上述途径中与碳、氮代谢相关的部分DEGs进行了进一步分析,相关DEGs信息见表3。在细胞壁组织代谢途径中的6个DEGs在干旱胁迫下均显著下调表达,这些DEGs主要包括纤维素合成酶基因(zm00001d043477、zm00001d032776),木质素漆酶(zm00001d042905、 zm00001d012408)等。碳水化合物代谢途径中编码磷酸甘露糖异构酶(zm00001d040020)基因在干旱胁迫下显著上调表达,己糖激酶基因(hexokinase)显著下调表达。光合作用途径的两个DEGs中上调表达的基因为编码NAD(P)H醌氧化还原酶基因(zm00001d019605),下调表达的基因为编码丝氨酸乙醛酸氨基转移酶基因(zm00001d031403)。蛋白质合成途径中30S 核糖体蛋白基因(zm00001d053713)表达量显著降低。总体而言,干旱胁迫对上述代谢途径基因表达主要呈抑制趋势。

表3 干旱胁迫下部分碳、氮代谢相关途径差异表达基因详情Table 3 Information of DEGs for carbohydrates and nitrogen related metabolic pathways in maize leaves under DS treatment
3 讨 论
本研究中,生育后期干旱显著降低玉米粒质量和生物量积累,且生物量在营养器官(茎秆、叶片等)中的分配比例增加,而籽粒中的分配比例(收获指数)则显著下降(图1),这与前人的研究结果一致[21]。干旱胁迫下玉米叶片种的可溶性糖含量及分配比例降低,籽粒淀粉含量降低,这与干旱胁迫下光合机能的下降有关[14]。在氮代谢方面,干旱胁迫下叶片氮素含量及分配比例均显著降低,这可能与干旱胁迫下叶片早衰导致的氮素流失有关[6]。干旱胁迫下籽粒氮收获指数显著降低,而茎秆中分配则显著增加。类似的结果前人的研究中也有报道,籽粒是玉米生育后期最大的氮汇,干旱胁迫显著限制了籽粒生长,导致籽粒氮素库容量显著降低,从而使得更多的氮素停留在茎秆等部位[22]。可见干旱胁迫对生育后期碳、氮同化物向籽粒的分配总体呈抑制作用,而使其更多地分布于营养器官中[23]。
在碳、氮代谢关键酶活性方面,干旱胁迫下叶片可溶性淀粉合成酶活性增加,可溶性糖含量降低,而淀粉含量显著增加,可见干旱胁迫促进叶片中可溶性糖向淀粉的转化。这与Abdelgawad等[14]的研究结果一致,即干旱胁迫可能会促进营养器官淀粉的生物合成,而较高的淀粉含量更有利于维持胁迫条件下的叶片生长及碳水化合物供应,缓解氧化伤害。与叶片中不同,籽粒可溶性淀粉合成酶活性在干旱胁迫下显著降低,这与籽粒淀粉含量及分配比例均下降的结果一致[15]。吐丝后50 d,干旱胁迫下叶片GS活力显著提高,这有助于清除逆境胁迫下产生的过量氨[24]。同时,干旱胁迫下GS活力的升高加速了叶片氮素转运,这与本研究中干旱胁迫下叶片氮含量及分配比例均显著降低的结果一致,类似的趋势在其他研究中也有报道[25]。
转录组测序结果表明(表3),在细胞壁组织代谢途径中,纤维素合成酶基因在干旱胁迫下均显著下调表达,其原因可能与干旱胁迫下碳水化合物供应受阻,纤维素合成酶活性及纤维素含量降低有关[14]。木质素是细胞壁的重要组分,植物组织木质化程度的高低对维持逆境胁迫下植株固有的形态至关重要,因而显著影响其抵御干旱等非生物胁迫的能力[26]。木质素漆酶是木质素的生物合成的关键[27],本研究中,干旱胁迫下两个木质素漆酶基因均显著下调表达,表明干旱胁迫下木质素的合成受到抑制,而纤维素、木质素等合成的抑制不利于植株维持其正常形态,从而引起植株萎蔫,加速衰老进程[28]。在光合作用代谢途径中,编码NAD(P)H醌氧化还原酶基因在干旱胁迫下显著上调表达,类似的趋势在其他研究中也有报道,其作用可能与维持干旱胁迫下的能量供应及蛋白的正常生理功能有关[29]。丝氨酸乙醛酸氨基转移酶基因显著下调表达,该酶主要参与光呼吸过程,随着叶片的衰老及光呼吸的降低,其活性显著下降[30]。在碳水化合物代谢途径中,干旱胁迫下编码磷酸甘露糖异构酶基因显著上调表达,该酶参与合成干旱胁迫下重要的渗透调节物质——甘露醇。因此,该基因的上调表达可能与干旱胁迫下甘露醇的合成有关[11,31]。己糖激酶基因显著下调表达,其与干旱胁迫下碳同化水平的降低有关[31]。
4 结 论
生育后期干旱胁迫下籽粒中可溶性糖、淀粉、氮素等的分配比例均显著降低,而茎秆及穗轴中的分配比例则显著增加,可见干旱胁迫不利于碳、氮同化物向籽粒分配。干旱胁迫下生育后期叶片GS活性显著提高,籽粒SSS活性显著降低,这与干旱胁迫下叶片氮含量及籽粒淀粉含量的降低有关。转录组测序结果表明,干旱胁迫对叶片细胞壁形成、碳水化合物代谢等代谢途径基因表达总体呈抑制作用,其中对木质素等合成途径的抑制作用可能是其引起叶片早衰的重要原因之一。
