钙离子和茉莉酸灌根对菜豆叶片防御酶活性和西花蓟马取食偏好性的影响
2023-05-30邱馨月郅军锐张涛曾广
邱馨月 郅军锐 张涛 曾广
摘要 為探明外源物质对菜豆植株防御酶活性和西花蓟马取食偏好性的影响。本试验分别采用CaCl2和茉莉酸(JA)进行灌根,研究了西花蓟马取食前后菜豆叶片脂氧合酶(LOX)、丙二烯氧化物合酶(AOS)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)和多酚氧化酶(PPO)活性的变化,并研究了西花蓟马对JA和CaCl2处理后的菜豆植株的取食偏好性。结果表明:不论CaCl2还是JA灌根均可诱导菜豆防御酶AOS、LOX、CAT和SOD活性的上升,其中对CAT活性影响最大,其活性分别上升至对照的2.83倍和3.57倍,但对POD和PPO活性没有影响。西花蓟马取食CaCl2处理的菜豆植株,能诱导LOX、POD和PPO活性分别上升1.40、2.03倍和2.26倍,而SOD活性则被显著抑制。西花蓟马取食JA灌根的菜豆植株能明显诱导POD和PPO活性的升高,二者活性分别是未取食植株的1.67倍和1.45倍,但AOS、LOX、CAT和SOD活性被抑制。西花蓟马2龄若虫取食CaCl2和JA处理的菜豆植株,造成的损伤面积只有对照的53%和28%,取食选择率分别降低71%和66%。上述结果表明,CaCl2和JA灌根能诱导菜豆植株的防御反应,减少蓟马的为害;西花蓟马取食能影响JA和CaCl2灌根的菜豆植株防御酶活性的变化。
关键词 西花蓟马; 菜豆; 茉莉酸; 氯化钙; 灌根诱导; 防御酶; 取食偏好
中图分类号: S 436.412.2
文献标识码: A
DOI: 10.16688/j.zwbh.2022034
Abstract This study aimed to investigate the effects of exogenous calcium and jasmonic acid (JA) on activities of defense enzyme in kidney bean plant and feeding preference of Frankliniella occidentalis. The kidney bean plants were separately treated with CaCl2 or JA by root irrigation. The changes of the activities of lipoxygenase (LOX), allene oxide synthase (AOS), superoxide dismutase (SOD), catalase (CAT), peroxidase (POD) and polyphenol oxidase (PPO) in the leaves of kidney bean before and after F.occidentalis feeding were assessed. At the same time, the feeding preference of F.occidentalis to kidney bean plant induced by CaCl2 and JA was revealed. The results showed that both CaCl2 and JA could increase the activities of AOS, LOX, CAT and SOD of kidney bean leaf. Among them, CAT was most activated, its activity increased 2.83 and 3.57 times of that of the control, respectively, while the activities of POD and PPO didnt significantly change. When F.occidentalis fed kidney bean leaf induced by CaCl2, the activities of LOX, POD and PPO increased by 1.40, 2.03 and 2.26 times, respectively, but SOD activity was significantly inhibited. When F.occidentalis fed leaf of kidney bean plant irrigated with JA, the activities of POD and PPO increased significantly to 1.67 and 1.45 times of that of healthy plant, while the activities of AOS, LOX, CAT and SOD were inhibited. When the 2nd instar larvae of F.occidentalis fed on leaves treated with CaCl2 and JA, the feeding areas were only 53% and 28% of that of the control, and the percentage of selectivity decreased 71% and 66%, respectively. The above results indicate that CaCl2 and JA irrigation can induce the defense of kidney bean plant and reduce the damage of F.occidentalis, and F.occidentalis feeding can cause the changes of defense enzyme activities of kidney bean plant irrigated with JA and CaCl2.
Key words Frankliniella occidentalis; kidney bean plant; jasmonic acid; calcium; irrigation root induction;defense enzyme; feeding preference
西花蓟马Frankliniella occidentalis是世界范围内重要的经济作物害虫[1]。作为典型的锉吸式口器昆虫,除了对寄主植物取食和产卵造成直接为害外,还能传播多种病毒形成间接为害[2]。目前对西花蓟马主要采用化学防治,但随着化学药剂的大量施用,环境污染以及西花蓟马的抗药性问题也随之产生[3]。而利用植物自身的诱导防御是减少化学药剂施用的理想选择,这种方法不仅有利于筛选抗性品种,也能缓解化学药剂带来的各类问题[4]。
茉莉酸(jasmonic acid,JA)及其衍生物是诱导植物防御反应的重要信号分子[5],在植物响应生物与非生物胁迫反应中起到举足轻重的作用[6]。目前关于外源JA类化合物诱导寄主植物防御西花蓟马的研究已有许多报道,但其处理方法多采用叶面喷施。例如JA喷施菜豆植株会对取食的西花蓟马造成不利影响,导致其提前转移到土壤中化蛹[7]。对菊花叶片局部喷施JA能降低西花蓟马的取食损害[8]。钙离子(Ca2+)不仅能促进植物生长发育,而且在植物信号转导中作为第二信使参与调控细胞生理代谢[9-10],其中就包括了对JA途径的调控。León等[11]的研究表明,在拟南芥中JA介导的信号转导途径受到钙调素的调节。Liu等[12]发现Ca2+参与了JA诱导番茄植株对烟粉虱Bemisia tabaci直接和间接的防御反应。除此之外,外施JA类化合物以及钙素还能诱导植物相关防御酶活性的变化。外源JA[13]、 CaCl2[14]处理菜豆植株均能诱导植株体内苯丙氨酸解氨酶(phenylalanine ammonialyase,PAL)和脂氧合酶(lipoxidase,LOX)升高。同时,CaCl2处理的菜豆植株被西花蓟马取食后其体内LOX、PAL以及β-1,3-葡聚糖酶的活性升高[14-15]。这种外源物对植株的诱导作用能增强植株对西花蓟马的防御反应,这种增强效应被定义为防御启动[16]。
植物作为一个有机整体,地上与地下部分关系十分紧密。目前,外源JA类化合物及钙离子通过处理根系对地上部抗虫性的诱导机制报道较少。Hamm等[17]发现JA处理水稻根系后植株上稻水象Lissorhoptrus oryzophilus的产卵量降低。Pierre等[18]发现JA处理西兰花根部能降低甘蓝地种蝇Delia radicum幼虫的化蛹率。Keeping等[19]发现用CaSiO3处理甘蔗根系周围土壤,能减少甘蔗非洲茎螟Eldana saccharina的数量及对茎秆的损害。
为进一步证明外源信号物质灌根对菜豆防御反应的诱导作用,本试验测定了外源JA和CaCl2灌根对菜豆叶片中6个防御酶活性的诱导作用,同时分析了西花蓟马取食对JA和CaCl2灌根诱导后的菜豆叶片防御酶活性的影响,并研究了JA和CaCl2灌根处理菜豆对西花蓟马取食偏好性的影响。研究结果为深入利用外源JA和钙离子增强植株抗虫性提供理论依据。
1 材料与方法
1.1 供试植物
菜豆Phaseolus vulgaris品种为‘金束鹿泰国地豆王(河北省辛集市盛农种子公司)。选取饱满的菜豆种子用灭菌营养土单株播种于营养钵内,于人工气候室内,在温度(25±1)℃,相对湿度(70±5)%,光周期L∥D=14 h∥10 h条件下培育,并保持无病虫为害,萌芽后11 d,待长出第一个三出复叶时,选取长势一致的植株供试。
1.2 供试昆虫
西花蓟马采自贵州省贵阳市花溪区的豆科植物上,并在贵州大学昆蟲研究所人工气候箱内[温度(25±1)℃,相对湿度(70±5)%,光周期L∥D=14 h∥10 h]用菜豆豆荚饲养50代以上。取生长发育一致的2龄初期若虫供试。
1.3 试验处理及取样
以0.10 mmol/L的JA[20]和20 mmol/L的CaCl[14]2作为处理浓度。选取生长发育一致的菜豆植株,每株浇灌30 mL JA或CaCl2溶液,以等体积去离子水灌根处理作为对照。于灌根后2 d,在菜豆植株第1对真叶上释放西花蓟马2龄初期若虫20头(10头/叶),以不接西花蓟马的健康植株为对照。接入西花蓟马后1 d,从各处理植株的第1对真叶上随机剪取0.1 g样本,测定防御酶活性。各处理设置5个生物学重复,共处理30株菜豆植株。
1.4 酶活性的测定
丙二烯氧化物合酶(AOS)活性测定参照泉州睿信生物公司提供的酶联免疫(ELISA)试剂盒说明书进行。脂氧合酶(LOX)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、多酚氧化酶(PPO)和过氧化物酶(POD)活性测定参照苏州科铭公司提供的试剂盒说明书进行。测定原理如下:
AOS:采用双抗体一步夹心法酶联免疫吸附试验(ELISA)。往预先包被植物AOS捕获抗体的包被微孔中依次加入样本、标准品、辣根过氧化物酶(HRP)标记的检测抗体,经过温育并彻底洗涤。用底物3,3′,5,5′-四甲基联苯胺(TMB)显色,TMB在HRP的催化下转化成蓝色,并在酸的作用下转化成最终的黄色。颜色的深浅和样品中的植物AOS浓度呈正相关。测定450 nm波长下吸光度,计算AOS活性。
LOX:LOX催化亚油酸氧化,氧化产物在280 nm处有特征吸收峰;测定280 nm波长下吸光度,计算LOX活性。
CAT:H2O2在240 nm下有特征吸收峰,CAT能够分解H2O2,使反应溶液在240 nm下的吸光度随反应时间延长而下降,测定240 nm波长下吸光度,计算CAT活性。
SOD:通过黄嘌呤及黄嘌呤氧化酶反应系统产生超氧阴离子(O-2),O-2可与水溶性四唑盐WST-8反应产生水溶性染料甲臜,后者在450 nm处有吸收峰;SOD可清除O-2,从而抑制了甲臜的形成。 测定450 nm波长下吸光度,计算SOD活性。
PPO:PPO能够催化邻苯二酚产生醌,后者在525 nm处有特征光吸收。测定525 nm波长下吸光度,计算PPO活性。
POD:POD催化H2O2氧化特定底物,在470 nm处有特征光吸收。测定470 nm波长下吸光度,计算POD活性。
1.5 西花蓟马取食损伤面积的测定
菜豆植株经不同处理溶液灌根后2 d,在每株菜豆第一对真叶的任意一片叶子上释放20头西花蓟马2龄若虫,被取食的菜豆叶片用培养皿(15 cm)托住,并用双通玻璃管(直径3.5 cm,高4 cm)将西花蓟马圈住,玻璃管底端用湿纸巾封住缝隙,顶端以纱网封口。于西花蓟马取食后1 d从叶柄剪取菜豆叶片,用1 mm2每格的坐标纸测定西花蓟马取食斑痕,累计西花蓟马取食损伤面积。各处理设置7个生物学重复。
1.6 西花蓟马取食偏好性
将塑料盒(底部直径23.5 cm,高11.5 cm)底部用马克笔均匀分为3个区域,然后将不同溶液灌根2 d的第二片菜豆真叶分别放置于不同区域内,再在装置中央接入60头西花蓟马2龄初期若虫,顶端以塑料盖封住。盖子中央用电热刀切割后用纱网封口。于蓟马取食后2 d,对不同处理的叶片及区域内的西花蓟马进行统计,以分布于各处理叶片区域的西花蓟马数量除以3个处理区域统计到的西花蓟马总数,换算成西花蓟马取食选择率。各处理设置10个生物学重复。
1.7 数据处理
采用Microsoft Excel 2003对数据进行汇总和处理,以SPSS 21.0对数据进行统计分析,不同溶液处理的健康菜豆叶片或被西花蓟马取食叶片的各种酶活性、取食面积和取食选择率差异显著性采用Tukey检验(P<0.05);同一溶液处理下西花蓟马取食前后酶活性的差异显著性采用t测验(P<0.05)。利用SigmaPlot 14.0 软件作图。
2 结果与分析
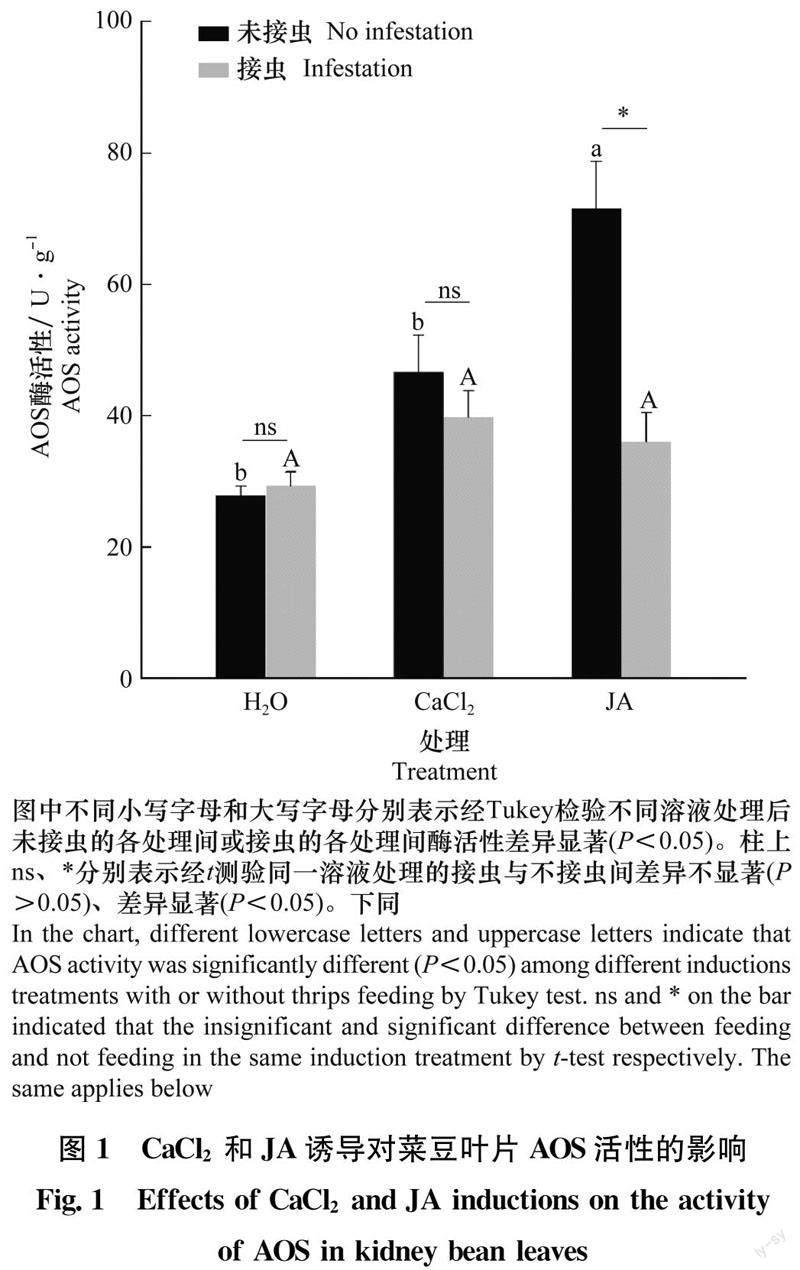
2.1 不同处理对菜豆叶片AOS活性的影响
对于健康植株,经CaCl2灌根后菜豆植株叶片AOS活性无显著变化(P>0.05),但经JA灌根后叶片AOS活性显著升高(P<0.05),为清水对照的2.57倍(图1)。西花蓟马取食清水、CaCl2和JA灌根的植株后,叶片AOS活性无明显差异(P>0.05)。同一溶液灌根后,西花蓟马取食未引起清水处理和CaCl2灌根植株叶片AOS活性出现显著变化(P>0.05),但显著抑制了JA灌根植株AOS活性(P<0.05),其AOS活性仅为健康植株的50%。
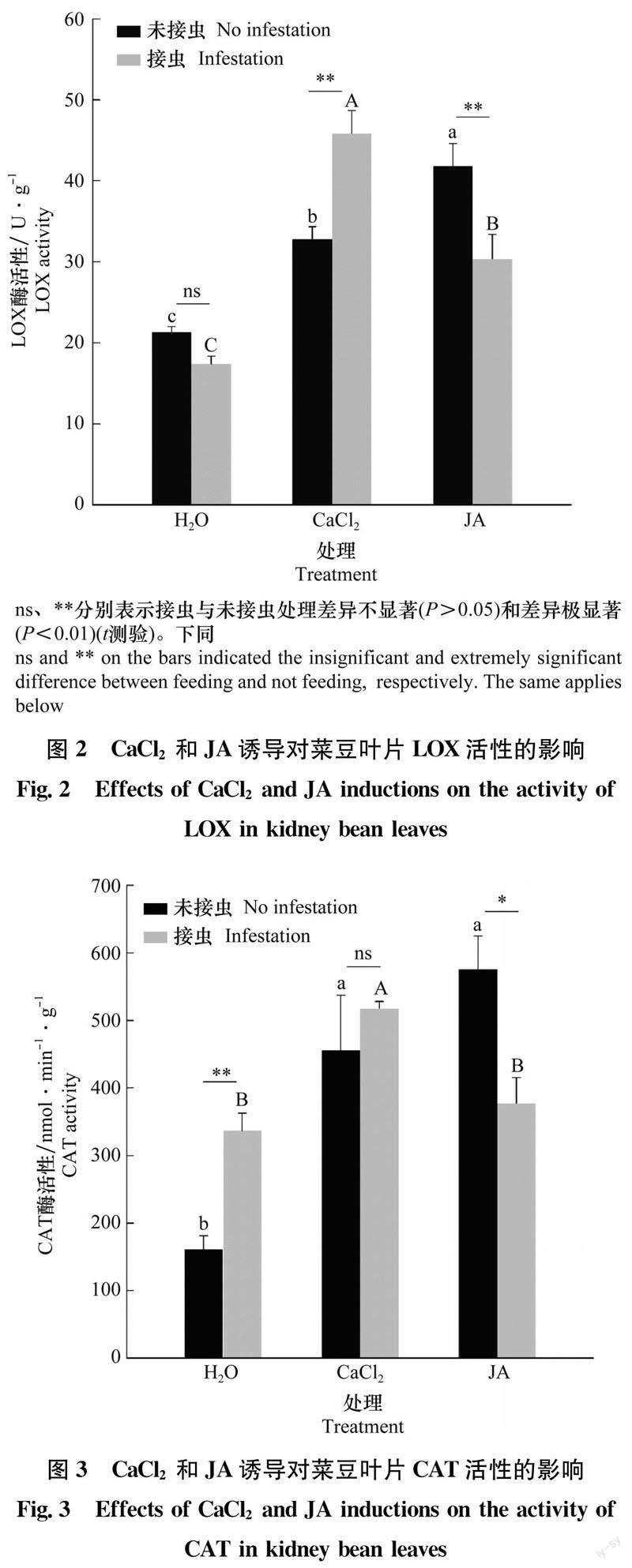
2.2 不同处理对菜豆叶片LOX活性的影响
经CaCl2和JA灌根后,无论是健康植株还是接种西花蓟马的植株,其叶片LOX活性均較清水处理的植株显著升高(P<0.05),但变化程度不同(图2)。CaCl2和JA灌根后叶片LOX活性分别为清水处理植株的1.54倍和1.96倍。接种西花蓟马后,菜豆叶片LOX活性分别为清水处理植株的2.64和1.75倍。相同溶液处理下,西花蓟马取食未引起清水对照植株LOX活性的变化(P>0.05),但使CaCl2灌根后叶片LOX活性提高了1.40倍(P<0.01),显著抑制了JA灌根后植株LOX的活性,仅为未取食植株的73%(P<0.01)。
2.3 不同处理对菜豆叶片CAT活性的影响
对于健康植株,经CaCl2和JA灌根后叶片CAT活性均显著升高(P<0.05),分别为对照的2.83倍和3.57倍(图3)。西花蓟马取食后,CaCl2灌根处理的植株叶片CAT活性显著高于清水对照的植株(P<0.05)。同一溶液处理下,西花蓟马取食前后菜豆叶片CAT活性变化不同,西花蓟马取食极显著提高清水处理植株叶片CAT活性(P<0.01),但未引起CaCl2灌根植株叶片CAT活性变化(P>0.05),显著抑制了JA灌根植株的CAT活性(P<0.05)。
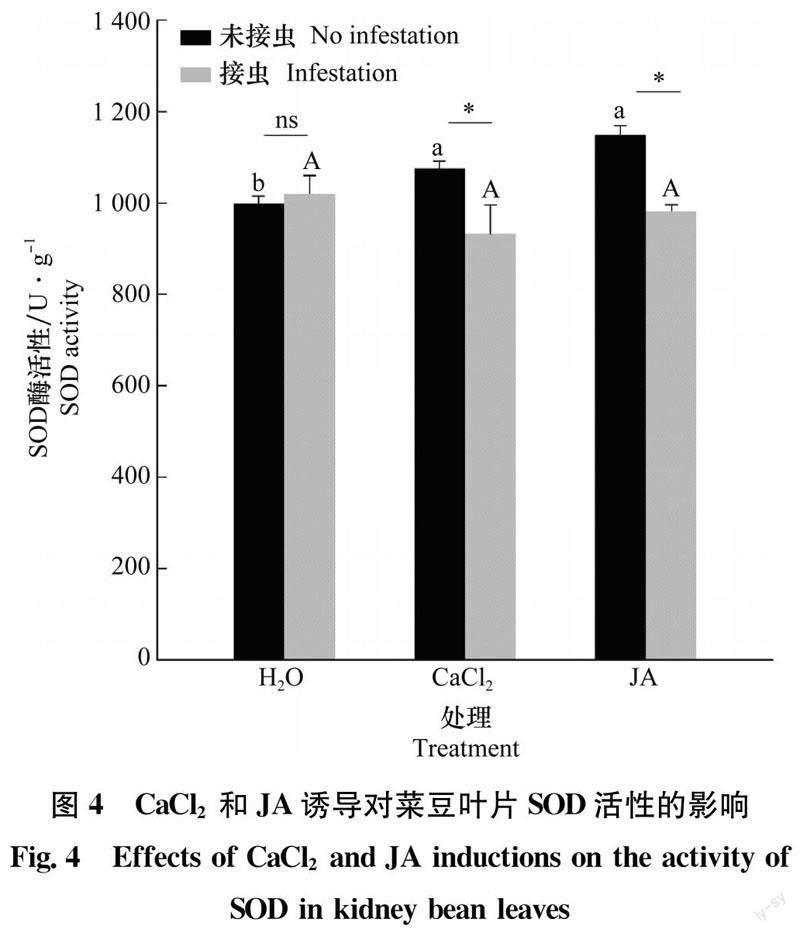
2.4 不同处理对菜豆叶片SOD活性的影响
健康植株经CaCl2和JA灌根后叶片SOD活性均显著升高(P<0.05),分别为对照的1.07和1.15倍(图4);但与清水处理的植株相比,CaCl2和JA灌根的植株被西花蓟马取食后,其叶片SOD活性均无显著变化(P>0.05)。同一溶液处理下,西花蓟马取食未引起清水处理植株叶片的SOD活性变化(P>0.05),但显著抑制了CaCl2和JA灌根植株的SOD活性(P<0.05),二者的活性分别只有健康植株的87%和86%。
2.5 不同处理对菜豆叶片POD活性的影响
健康植株经CaCl2和JA灌根后,叶片POD活性虽有所变化,但与清水处理相比均无明显差异(P>0.05)(图5)。CaCl2和JA灌根的植株经西花蓟马取食后,叶片POD活性均较清水处理后被取食的植株显著升高(P<0.05),分别为清水处理植株的1.74和2.08倍。相同溶液处理下,CaCl2和JA灌根植株被西花蓟马取食后,其POD活性是清水处理后被取食植株的2.03和1.67倍(P<0.01)。
2.6 不同处理对菜豆叶片PPO活性的影响
对健康植株,CaCl2灌根后PPO活性显著降低(P<0.05),仅为清水处理的60%,而JA灌根后叶片PPO活性无明显变化(P>0.05)(图6);与清水处理后接种西花蓟马相比,CaCl2灌根植株被西花蓟马取食后,叶片PPO活性虽有所升高,但未达到显著水平(P>0.05);而JA灌根植株被西花蓟马取食后,叶片PPO活性提升了1.37倍(P<0.05)。相同溶液处理下,西花蓟马取食未引起清水处理植株PPO活性的明显变化(P>0.05),但能诱导CaCl2和JA灌根植株叶片PPO的活性升高,其活性分别提高了2.26倍和1.45倍(P<0.01)。
2.7 不同处理对西花蓟马取食损伤面积和取食偏好性的影响
CaCl2和JA灌根后,西花蓟马对菜豆叶片的取食损伤面积和取食选择率相比清水处理都显著降低(P<0.05)(图7),西花蓟马对CaCl2和JA处理的菜豆植株叶片的损伤面积仅为对照的53%和28%,且对菜豆叶片的取食选择率也显著下降,分别较对照下降了71%和66%,但对两种溶液灌根处理叶片的取食选择率无显著差异。
3 结论与讨论
植物在应对来自于外界环境胁迫时,会产生一系列防御性生理生化反应,其中防御酶AOS、LOX、CAT、SOD、POD和PPO是植物增强防御能力的关键调控因子[21]。AOS作为JA生物合成途径中的主要限速酶,可以通过控制JA的合成而参与调节植物的许多生理过程[22]。LOX同样是JA合成途径中的特异性酶,能催化不饱和脂肪酸氧化形成JA,在植物受伤信号识别与转导中起着重要作用[23]。CAT、POD、SOD是植物体内酶促清除系统的关键酶类,能有效抑制活性氧自由基对植物的伤害,从而提高植株的抗性[24]。PPO在植物受到外界胁迫时催化酚类生成醌,醌能作为抗营养因子抵御昆虫取食和抑制细菌繁殖[25]。目前已有研究证明,外施钙素、JA类化合物以及昆虫取食都能诱导植物相关防御酶活性的变化。
钙离子作为胞内第二信使,调节植物体内各防御信号途径使之井然有序地进行[26]。本研究发现外源CaCl2灌根同JA处理一样能诱导菜豆植株叶片LOX、CAT、SOD活性升高,说明外源CaCl2与JA灌根对菜豆叶片内防御酶活性有相似的诱导作用。他人也有类似的报道,如外源CaCl2处理菜豆植株能诱导叶片LOX活性显著上升[15],处理花生叶片能诱导CAT活性显著升高[27]。同时CaCl2还能抑制强光对小麦叶片细胞膜的破坏,提高高温强光胁迫下的小麦叶片内SOD含量[28]。除此之外,本研究还发现CaCl2对菜豆叶片PPO活性具有抑制作用。这与刘娜等[29]发现不同钙肥配施处理的果皮PPO活性显著降低相似。原因可能是植株在生长过程中能产生大量的次生代谢物酚类,而Ca2+能够维持细胞膜系统完整性与稳定性,从而减少了酚类物质与PPO的接触[30]。
JA是自然界中广泛存在的一种信号分子,参与植物生长和发育过程,并且在调节植物应对生物和非生物逆境的反应中起着重要的作用[31]。本研究发现外源JA灌根可诱导菜豆植株叶片AOS、LOX、CAT、SOD活性升高,但未引起POD和PPO活性发生明显变化。Laudert等[22]证实AOS作为JA合成途径中的第一个特异性酶,其AOS基因的启动子可在JA作用下被激活。其他报道中也有和本研究相似的结果,如外源JA处理番茄、棉花和油菜植株能分别诱导植株LOX、CAT和SOD活性升高,从而增强植株的抗虫性[32-34]。并且在本研究中,西花蓟马取食JA与CaCl2灌根处理的菜豆叶片后AOS和SOD活性无显著变化,这可能由于植物防御反应是一个消耗大量能量且存在时间效应的过程[35],或者是植株為了权衡生长和防御,在生长阶段转移了原本可以用于防御的资源[36]。
昆虫取食能诱导激活植物体内一系列信号转导途径从而产生多种抗性机制[37]。本研究发现西花蓟马取食健康菜豆植株能诱导CAT活性上升,这与黄钰等[38]发现水稻在受白背飞虱Sogatella furcifera取食后,其茎、叶中CAT活性显著增加的研究结果一致。但在本研究中西花蓟马取食并不能诱导健康菜豆植株LOX、AOS、SOD、PPO和POD活性发生变化,这与本课题组前期的研究结果存在差异[32]。可能由于本试验是在西花蓟马取食1 d后测定的指标,而昆虫取食诱导是一个渐变的过程,除与害虫数量、分布等因子有关外,和时间也密切相关,从而导致酶活性的变化存在时间效应[39]。
除此之外,西花蓟马取食CaCl2处理的菜豆后能明显激活LOX活性。这与Zeng等[14]及刘勇等[15]的研究结果相同,即外源CaCl2可以通过蓟马取食增强植物的LOX酶活性。在本研究中还发现西花蓟马取食CaCl2、JA处理的菜豆后POD和PPO活性显著升高,说明蓟马取食对CaCl2和JA的诱导具有加强作用,促进了植株的防御启动,但同时JA处理的AOS、CAT和SOD活性受到一定程度的抑制,这可能是因为植食性昆虫取食时分泌的唾液中某些物质诱导的结果[40]。
施用外源JA和CaCl2增加了植株抗性,从而对西花蓟马产生不利影响。本研究发现外源JA和CaCl2灌根后,西花蓟马的取食面积和取食选择率较对照都显著降低,表明JA和CaCl2灌根诱导增强了菜豆植株对西花蓟马的防御作用。前人还发现外源JA和CaCl2还能影响西花蓟马的生长发育和种群动态。牟峰等[41]研究发现西花蓟马取食喷施外源JA的菜豆植株后其发育期延长,存活率下降,种群数量也有所降低。Zeng等[42]研究发现外源CaCl2处理的菜豆植株上西花蓟马的数量较少,发育时间较长,寿命和繁殖量都较低。
本研究证实了外源JA和CaCl2灌根均能诱导菜豆植株的防御反应,从而影响西花蓟马的取食偏好性。叶片中各防御酶活性的变化程度不同,可能与植物能量的分配有关,且各酶的合成途径也不相同[13]。因此,防御酶活性变化在诱导防御中的功能和各种酶的协调作用有待进一步探讨。目前对于JA和钙素灌根对植物抗虫性的相关研究较少,灌根诱导和喷施诱导的具体作用机制的差异也有待深入研究。
本论文只研究了CaCl2和JA分别灌根和灌根后接种西花蓟马叶片各种酶活性的变化,及CaCl2和JA灌根对西花蓟马取食和选择性的影响,但CaCl2和JA灌根是否对菜豆本身生长发育和植株生物量增长有影响未做研究。前人研究发现CaCl2和JA除了影响植物防御酶活性外,对植物本身也有一定的作用。JA类化合物能提高柑橘光合作用,促进叶绿素的积累[43],在干旱胁迫下促进水稻的生长[44],外源Ca2+也能提高花生光合速率和产量[45]。后续试验将进一步开展CaCl2和JA对植株生长发育影响的研究,以期更加系统全面地掌握CaCl2和JA介导的寄主植物和西花蓟马的互作。
参考文献
[1] 吴青君, 张友军, 徐宝云, 等. 入侵害虫西花蓟马的生物学、危害及防治技术[J]. 中国植保导刊, 2005, 42(1): 11-14.
[2] KIRK W D J, TERRY I. The spread of the western flower thrips Frankliniella occidentalis (Pergande) [J]. Agricultural and Forest Entomology, 2003, 5(4): 301-310.
[3] 陈雪林, 杜予州, 王建军. 西花蓟马抗药性研究进展[J]. 植物保护, 2011, 37(5): 34-38.
[4] 张海静, 严盈, 彭露, 等. 韧皮部取食昆虫诱导的植物防御反应[J].昆虫学报, 2012, 55(6): 736-748.
[5] 于涌鲲, 郝玉兰, 万善霞, 等. 茉莉酸类物质的生物合成及其信号转导研究进展[J]. 自然科学进展, 2008, 18(9): 961-967.
[6] MUR L A J, KENTON P, ATZORN R, et al. The outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death [J]. Plant Physiology, 2006, 140(1): 249-262.
[7] EGGER B, KOSCHIER E H. Behavioural responses of Frankliniella occidentalis Pergande larvae to methyl jasmonate and cis-jasmone [J]. Journal of Pest Science, 2014, 87(1): 53-59.
[8] CHEN Gang, KIM H K, KLINKHAMER P G, et al. Site-dependent induction of jasmonic acid-associated chemical defenses against western flower thrips in Chrysanthemum [J/OL]. Planta, 2020, 251(1): 8. DOI:10.1007/s00425-019-03292-2.
[9] ALLEN G J, CHU S P, HARRINGTON C L, et al. A defined range of guard cell calcium oscillation parameters encodes stomatal movements [J]. Nature, 2001, 411(6841): 1053-1057.
[10]DAVID L, RAOUL R, ALAIN P. Calcium in plant defence-signalling pathways [J]. The New Phytologist, 2006, 171(2): 249-269.
[11]LEN J, ROJO E, TITARENKO E, et al. Jasmonic acid-dependent and -independent wound signal transduction pathways are differentially regulated by Ca2+/calmodulin in Arabidopsis thaliana [J]. Molecular & General Genetics, 1998, 258(4): 412-419.
[12]LIU Xin, SABIR H, XIE Wen, et al. Ca2+ signal contributing to jasmonic acid-induced direct and indirect defense against the whitefly Bemisia tabaci in tomato plants [J]. Entomologia Experimentalis et Applicata, 2021, 169(9): 848-858.
[13]从春蕾, 郅军锐, 廖启荣, 等. 蓟马取食、机械损伤以及外源水杨酸甲酯和茉莉酸对菜豆叶片防御酶活性的影响[J]. 昆虫学报, 2014, 57(5): 564-571.
[14]ZENG Guang, ZHI Junrui, YE Mao, et al. Inductive effects of exogenous calcium on the defense of kidney bean plants against Frankliniella occidentalis (Thysanoptera: Thripidae) [J]. Arthropod-Plant Interactions, 2020(1): 473-480.
[15]劉勇, 郅军锐. 钙素在西花蓟马取食诱导菜豆防御酶活性中的作用[J]. 西南师范大学学报(自然科学版), 2018, 43(3): 58-64.
[16]VAN HULTEN M, PELSER M, VAN LOON L, et al. Costs and benefits of priming for defense in Arabidopsis [J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(14): 5602-5607.
[17]HAMM J C, STOUT M J, RIGGIO R M. Herbivore- and elicitor-induced resistance in rice to the rice water weevil (Lissorhoptrus oryzophilus Kuschel) in the laboratory and field [J]. Journal of Chemical Ecology, 2010, 36(2): 192-199.
[18]PIERRE P S, DUGRAVOT S, CORTESERO A M, et al. Broccoli and turnip plants display contrasting responses to belowground induction by Delia radicum infestation and phytohormone applications [J]. Phytochemistry, 2012, 73(1): 42-50.
[19]KEEPING M G, MEYER J H. Calcium silicate enhances resistance of sugarcane to the African stalk borer Eldana saccharina Walker (Lepidoptera: Pyralidae) [J]. Agricultural and Forest Entomology, 2015, 4(4): 265-274.
[20]李顺欣, 郅军锐, 杨广明, 等. 外源茉莉酸诱导的菜豆叶片生化抗性及其对西花蓟马体内保护酶和解毒酶活性的影响[J]. 应用生态学报, 2017, 28(9): 2975-2983.
[21]郭祖国, 王梦馨, 崔林, 等. 6种防御酶调控植物体应答虫害胁迫机制的研究进展[J]. 应用生态学报, 2018, 29(12): 360-370.
[22]LAUDERT D, WEILER E W. Allene oxide synthase: a major control point in Arabidopsis thaliana octadecanoid signalling [J]. The Plant Journal, 1998, 15(5): 675-684.
[23]KESSLER A, HALITSCHKE R, BALDWIN I T. Silencing the jasmonate cascade: induced plant defenses and insect populations [J]. Science, 2004, 305(5684): 665-668.
[24]PUTHOFFD P, SMIGOCKI A C. Insect feeding-induced differential expression of Beta vulgaris root genes and their regulation by defense-associated signals [J]. Plant Cell Reports, 2007, 26(1): 71-84.
[25]李天来, 李淼, 孙周平. 钙和水杨酸对亚高温胁迫下番茄叶片保护酶活性的调控作用[J]. 应用生态学报, 2009, 20(3): 586-590.
[26]檀龙颜, 马洪娜. 植物响应钙离子胁迫的研究进展[J]. 植物生理学报, 2017, 53(7): 1150-1158.
[27]孟丹娜. 外源化学物质对花生叶斑病诱导抗性及其机理研究[D]. 沈阳: 沈阳农业大学, 2018.
[28]杨亚军, 李利红, 赵会杰, 等. Ca2+对高温强光胁迫下小麦叶片光合机构的保护作用[J]. 麦类作物学报, 2008, 28(6): 1042-1047.
[29]刘娜, 金昕, 谢昶琰, 等. 诱导条件下不同配施钙肥处理减缓‘黄冠梨果面褐斑病的研究[J]. 南京农业大学学报, 2017, 40(5): 867-873.
[30]龚新明, 关军锋, 张继澍,等. 钙、硼营养对黄冠梨品质和果面褐斑病发生的影响[J]. 植物营养与肥料学报, 2009, 15(4): 942-947.
[31]CREELMAN R A, MULLET J E. Biosynthesis and action of jasmonates in plants [J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1997, 48(1): 355-381.
[32]蒲恒浒, 郅军锐, 曾广, 等. 不同诱导处理对番茄叶片營养物质含量及防御酶活性的影响[J]. 环境昆虫学报, 2019, 41(2): 405-412.
[33]杨世勇, 王蒙蒙, 谢建春. 茉莉酸对棉花单宁含量和抗虫相关酶活性的诱导效应[J]. 生态学报, 2013, 33(5): 1615-1625.
[34]冯建雄, 常静, 李海平, 等. 外源茉莉酸诱导对油菜防御酶活性和4种物质含量及植株生长的影响[J]. 植物保护, 2021, 47(1): 128-134.
[35]ZAINAB A, CHEN Mingshun. Indirect plant defense against insect herbivores: a review [J]. Insect Science, 2018, 25(1): 2-23.
[36]ZHANG Nailou, ZHAO Bin, FAN Zhijing, et al. Systematic identification of genes associated with plant growth-defense tradeoffs under JA signaling in Arabidopsis [J/OL]. Planta, 2020, 251(2):43. DOI: 10.1007/s00425-019-03335-8.
[37]余源婵, 杨茂发, 商胜华, 等. 不同昆虫取食对烟草信号分子和防御酶的影响[J].环境昆虫学报, 2021, 43(4): 967-977.
[38]黄钰, 王玥, 杨金睿, 等. 云南哈尼梯田地方水稻品种月亮谷对白背飞虱取食的主要防御反应[J]. 南方农业学报, 2021, 52(1): 11-19.
[39]严善春, 袁红娥, 王琪, 等. 叶损伤诱导兴安落叶松针叶中10种酚酸的变化[J]. 应用生态学报, 2010, 21(4): 1000-1006.
[40]TURLINGS T, ALBORNl H T, LOUGHRIN J H, et al. Volicitin, an elicitor of maize volatiles in oral secretion of Spodoptera exigua: isolation and bioactivity [J]. Journal of Chemical Ecology, 2000, 26(1): 189-202.
[41]牟峰, 郅军锐, 从春蕾, 等. 外源茉莉酸和水杨酸甲酯对西花蓟马生长发育和种群动态的影响[J]. 植物保护, 2014, 40(6): 70-74.
[42]ZENG Guang, ZHI Junrui, YE Mao, et al. Life table and preference choice of Frankliniella occidentalis (Thysanoptera: Thripidae) for kidney bean plants treated by exogenous calcium [J/OL]. Insects, 2021, 12(9): 838. DOI: 10.3390/insects12090838.
[43]QIU Xia, XU Yinghuan, XIONG Bo, et al. Effects of exogenous methyl jasmonate on the synthesis of endogenous jasmonates and the regulation of photosynthesis in citrus [J]. Physiologia Plantarum, 2020, 170(3): 398-414.
[44]LLYAS N, GULL R, MAZHAR R, et al. Influence of salicylic acid and jasmonic acid on wheat under drought stress [J]. Communications in Soil Science and Plant Analysis, 2017, 48(22): 2715-2723.
[45]王飛, 何春梅, 李清华, 等. 外源钙水平与花生下针期不同土壤水分对植株生理特性的影响[J]. 植物营养与肥料学报, 2013, 19(3): 623-631.
(责任编辑:杨明丽)
