中国狸尾豆属及其近缘属植物的叶脉序特征分析
2023-05-30段微微周盛茂赵雪利
段微微 周盛茂 赵雪利


摘 要:探究貍尾豆属Uraria Desv.及其近缘属蝙蝠草属Christia Moench、算珠豆属Urariopsis Schindl.叶脉序特征在属间及种间的变异规律和分类学意义,为其作为药用植物资源的鉴定、开发利用及保护提供资料。采用清净法制作标本,观察和描述了该类群12个物种的叶脉序特征,基于叶脉序特征进行主成分分析和聚类分析,并编制分种检索表。结果表明:(1)狸尾豆属及其近缘属叶脉序特征具有一致性,主要体现在其都为直行羽状脉,间二级脉为复合型(铺地蝙蝠草除外),三级脉为网结型与贯穿型兼有,叶缘末级脉汇聚于叶缘;(2)属间或属内种间有分类价值的叶脉序特征有顶生小叶叶形、顶生小叶长宽比、二级脉对数及间距、二级脉及三级脉与主脉夹角的变化、脉间区发育情况、盲脉分支情况等;(3)聚类分析显示这12个种聚为两大支,狸尾豆属、蝙蝠草属和算珠豆属物种未各自聚为一支,其中台湾蝙蝠草和长圆叶狸尾豆与狸尾豆属物种聚为一支,而短序算珠豆与蝙蝠草属物种聚为一支,分别显示出较近的亲缘关系。研究显示顶生小叶叶形、顶生小叶长宽比、二级脉类型、二级脉对数及脉间距、二级脉与主脉夹角及其变化、三级脉与主脉夹角变化、脉间区发育情况、盲脉分支情况是狸尾豆属及其近缘属具有分类价值的叶脉序特征。
关键词:豆科;狸尾豆属;蝙蝠草属;叶脉序;分类学
中图分类号:Q 948 文献标志码:A 文章编号:0253-2301(2023)02-0034-12
DOI: 10.13651/j.cnki.fjnykj.2023.02.006
Abstract: By exploring the variation pattern and taxonomical significance of the leaf venation characteristics between the genera and species of Uraria Desv. and its relative genera (Christia Moench and Urariopsis Schindl.), it could provide data for the identification, development, utilization and protection when they were regarded as the medicinal plant resources. The leaf venation characteristics of 12 species of this group were observed and described by using the cleaning method to make the specimens. The principal component analysis and cluster analysis were carried out based on the characteristics of leaf venation, and a classification retrieval table of the species was compiled. The results showed that: (1) The characteristics of the leaf venation of Uraria Desv. and its relative genera had consistency, which were mainly reflected in the fact that they were straight pinnate veins, the intersecondary veins were compound (except for Christia obcordata), the tertiary veins were both reticular and penetrating, and the last veins of the leaf margin converged at the leaf margin. (2) The characteristics of the leaf venation with taxonomic value among the genera or the species within the genera included the leaf shape of the terminal leaflet, the lengthwidth ratio of the terminal leaflet, the logarithm of the secondary veins and the interveinal distance, the angle between the secondary vein / tertiary vein and the main vein and its change, the areole development, and the branch of blind veins. (3) The cluster analysis showed that the 12 species were clustered into two major branches, and the species of Uraria Desv., Christia Moench and Urariopsis Schindl. were not clustered into one branch. Among them, the species of Christia campanulata and Uraria oblonga were clustered into one branch with the species of Uraria Desv., while the species of Urariopsis brevissima was clustered into one branch with the species of Christia Moench, showing a relatively close genetic relationship. The results showed that the leaf shape of the terminal leaflet, the lengthwidth ratio of the terminal leaflet, the type of the secondary veins, the logarithm of the secondary veins and the interveinal distance, the angle between the secondary vein and the main vein and its change, the angle between the tertiary vein and the main vein and its change, the areole development, and the branch of blind veins were the characteristics of the leaf venation with taxonomic value of Uraria Desv. and its relative genera.
Key words: Fabaceae; Uraria Desv. Christia Moench; Leaf venation; Taxonomy
狸尾豆属Uraria Desv.隶属于豆科Fabaceae山蚂蝗族Desmodieae,全世界约有20种,主要分布于热带非洲、亚洲和大洋洲[1];中国有7~9种,主要分布于25°N以南的地区,以云南的种类为最多[2-3]。貍尾豆属植物最显著的形态特征是其荚果的各荚节反复折叠,同样具有该荚果类型的为其近缘属蝙蝠草属Christia Moench和算珠豆属Urariopsis Schindl.。狸尾豆属、蝙蝠草属及算珠豆属部分种类为重要的药用植物资源,如猫尾草Uraria crinita(L.)Desv.ex DC.,散瘀止血、清热止咳的功效;美花狸尾豆Uraria picta(Jacq.)Desv.ex DC.有平肝、宁心、健脾的功效;蝙蝠草Christia vespertilionis(L.f.) Bahn.F.可治肺结核、虫蛇咬伤,叶外敷为跌打接骨药等[3]。作为重要的药用植物资源,准确的物种鉴定是对其资源开发利用和保护的前提,而狸尾豆属及其近缘属的分类和进化一直存在争议[2-3]。叶脉序是指叶脉在每片叶子中粗细差异、走向独特分布排序构成的特征图案[10]。叶脉序特征具有变异丰富且稳定,易于观察等特点,可以为植物分类鉴定和系统发育构建提供依据,准确鉴定物种和推断其亲缘关系[4],从而为药用植物的保护利用以及通过系统研究方法寻找新的药用植物资源提供可靠的参考资料。
狸尾豆属及其近缘属与豆科其他类群较易区分,然而狸尾豆属与蝙蝠草属、算珠豆属的属间界限、系统位置以及属内种间关系,以及一些分类有争议的物种,如台湾蝙蝠草Christia campanulata(Wall.) Thoth.、长圆叶狸尾豆Uraria oblonga(Wall.ex Benth.) H.Ohashi & K.Ohashi、短序算珠豆Urariopsis brevissimaYang et Huang等的系统位置,都存在争议[3,5-7]。叶脉研究起源于20世纪植物系统分类学和植物化石鉴定建立的技术方法,其稳定性对于植物的起源与分类研究具有重要价值[11]。古植物学家可以通过植物化石的叶形、主脉、二级脉和间二级脉、三级脉、叶缘末级脉序走向及类型等特征鉴定化石植物[12]。叶脉序结构特征不仅可以为植物化石的研究提供重要信息,也可以为现代植物分类学研究提供参考,Hickey[8]通过对叶脉序研究发现,次级脉序和高级脉序结构特征在植物识别和分类中具有重要作用;Ellis等[9]对这一方法进一步研究,并形成描述分类的标准方法。叶脉序特征已在不同的植物类群中被应用,并为一些复杂的分类问题提供了重要证据,为科间、属间以及种间的分类鉴定提供了参考[13-15, 20, 30-32]。
狸尾豆属及其近缘属的叶脉序特征变异规律,及其在物种鉴定中的分类价值有待深入研究。本研究对中国狸尾豆属、蝙蝠草属和算珠豆属植物的叶脉序特征进行观察和比较,探讨叶脉序特征在狸尾豆属及近缘属中的变异规律及分类学意义,以期为狸尾豆属及其近缘属的分类提供新依据,并为其作为药用植物的鉴定、开发利用及保护提供资料。
1 材料与方法
1.1 试验材料
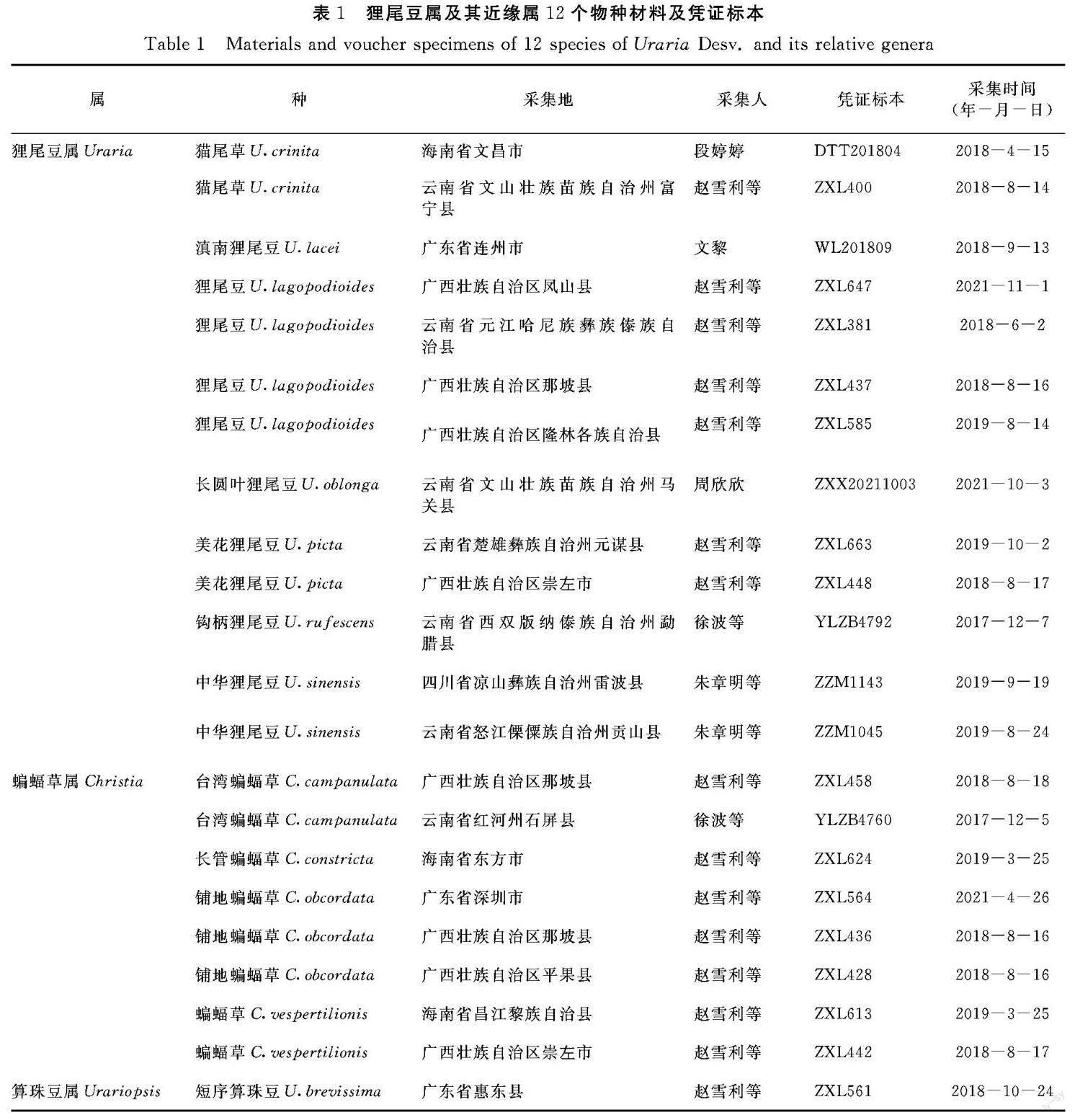
以中国狸尾豆属、蝙蝠草属及算珠豆属的12个物种为研究对象,试验材料取自移栽植株或蜡叶标本(表1)。
1.2 试验方法
叶脉标本的处理参考喻诚鸿等[16]描述的清净法。具体步骤为:(1)将叶片置于水浴锅中煮沸10~15 min,具体时间根据叶片质地调整;(2)待叶片软化且排出气体后,置入9%的NaOH溶液中,恒温加热2~3 h,至半透明,用毛刷刷去残余叶肉;(3)将清洗干净的叶脉标本置于30%的NaClO溶液中,30℃漂白5~20 min;(4)在0.5%番红溶液中染色30 min,然后用清水多次冲洗以分色,得到完整清晰的叶脉标本并封片,在LEICA MC170 HD显微镜下观察、拍照。
描述术语参考喻诚鸿等[16]、孙启高等[17]对叶脉序特征的研究。根据研究类群的特征,选取了17个叶脉序特征(表2)。根据实际野外采集和植物生长情况,每个物种选取至少10片成熟顶生小叶片进行观察、统计。
1.3 数据分析
使用游标卡尺对叶片长度和宽度进行精确测量。脉间区发育情况、盲脉分支情况以及叶缘末级脉序类型在光学显微镜下观察;二级脉与主脉夹角、二级脉与主脉夹角变化、三级脉与主脉夹角基于拍摄的图片,使用imageJ图像处理软件进行测量。叶形、二级脉类型及对数、脉间距变化规律、间二级脉发生频率以及类型通过直接观察获得。对原始数据进行编码(表2),利用SPSS Statistics 26将所得的数据矩阵进行标准化处理,然后进行Pearson相关性分析、主成分分析,并基于组间欧氏距离进行聚类分析。
2 结果与分析
2.1 狸尾豆属及其近缘属的叶脉序特征分析
由图2、表3可知,狸尾豆属、蝙蝠草属及算珠豆属12个种的叶脉序特征存在丰富的变异,也具有一定的共性特征,具体为:主脉纤细,直向行走;除铺地蝙蝠草无间二级脉,其余各物种间二级脉为复合型;三级脉结构均为结网型与贯穿型相结合;叶缘末级脉序汇聚于边缘。
狸尾豆属7个物种的顶生小叶以长椭圆形为主;主脉纤细,直向行走;二级脉为直达叶缘型,6~19对,脉间距不规则;二级脉与主脉夹角,为窄锐角;二级脉与主脉夹角向基部递增;间二级脉为复合型;三级脉由网结型与贯穿型结合而成;三级脉与主脉夹角由主脉向叶缘以窄锐角向近直角变化;脉间区发育中等,形状不规则,大小略有变化,4~5边;盲脉分支多为不均等分支;叶缘末级脉序汇聚于边缘(图2A~G)。
蝙蝠草属4个物种的顶生小叶,除台湾蝙蝠草、长管蝙蝠草外,均为倒三角形;主脉纤细,直向行走;二级脉为直达叶缘型,3~18对,脉间距不规则;二级脉与主脉夹角由窄锐角至宽锐角变化;除铺地蝙蝠草外,间二级脉均为复合型;三级脉由网结型与贯穿型结合而成;三级脉与主脉夹角由主脉向叶缘以中锐角至近直角变化;叶缘末级脉序汇聚于边缘(图2H~K)。
算珠豆属有2个物种,本研究仅有短序算珠豆1种,其顶生小叶心形;主脉纤细,直向行走;二级脉为直达叶缘型,9~11对,脉间距规则;二级脉与主脉夹角,为中锐角;二级脉与主脉夹角向基部递减;间二级脉为复合型;三级脉与主脉夹角由主脉向叶缘以中锐角至宽锐角变化;脉间区发育中等,形状、大小较一致,以4边居多;盲脉一次分支,叶缘末级脉序汇聚于边缘(图2L)。
2.2 狸尾豆属及其近缘属叶脉序特征的分种描述
2.2.1 猫尾草Uraria crinita(L.)Desv.ex DC. 叶卵形或长椭圆形,长宽比为1.62~4.87;主脉纤细,直向行走;二级脉9~12对,为环结曲行脉或真曲行脉型,其脉间距常不规则;间二级脉为复合型;主脉与二级脉夹角一致,为窄锐角;三级脉与主脉夹角由主脉向叶缘以窄锐角至近直角变化;脉间区发育中等,形状不规则、大小略有变化,多为4边;盲脉纤细为不均等分支,叶缘末级脉序汇聚于边缘(图2A)。
2.2.2 滇南狸尾豆Uraria lacei Craib 叶倒卵形或长椭圆形,长宽比为1.38~2.11;主脉纤细,直向行走;二级脉9~13对,为直达叶缘型,其脉间距常不规则;间二级脉为复合型;主脉与二级脉夹角向基部递增,为窄锐角;三级脉与主脉夹角由主脉向叶缘以中锐角至宽锐角变化;脉间区发育中等,形状不规则、大小略有变化,多为4~5边;盲脉为不均等分支,叶缘末级脉序汇聚于边缘(图2B)。
2.2.3 狸尾豆Uraria lagopodioides(L. ) Desv.ex DC. 叶卵形,长宽比为1.01~1.99;主脉纤细,直向行走;二级脉6~11对,为真曲行脉、环结曲行脉或直达叶缘脉型,其脉间距规则;间二级脉为复合型;主脉与二级脉夹角向基部递增,常为窄锐角;三级脉与主脉夹角由主脉向叶缘以中锐角至近直角变化;脉间区发育中等,形状不规则、大小略有变化,4边居多;盲脉纤细为不均等分支,叶缘末级脉序汇聚于边缘(图2C)。
2.2.4 长圆叶狸尾豆Uraria oblonga(Wall.ex Benth.) H.Ohashi & K.Ohashi 葉长椭圆形,长宽比为3.65~3.91;主脉纤细,直向行走;二级脉15~16对,为真曲行脉型,其脉间距规则;间二级脉为复合型;主脉与二级脉夹角一致,常为窄锐角;三级脉与主脉夹角由主脉向叶缘变化一致,为宽锐角;脉间区发育中等,形状不规则、大小略有变化,为4~5边;盲脉一次分支,叶缘末级脉序汇聚于边缘(图2D)。
2.2.5 美花狸尾豆Uraria picta(Jacq.)Desv.ex DC. 叶狭披针形,长宽比为4.15~12.62;主脉纤细,直向行走;二级脉8~13对,为直达叶缘脉或真曲行脉型,其脉间距规则;间二级脉为复合型;主脉与二级脉夹角向基部递增,常为窄锐角;三级脉与主脉夹角由主脉向叶缘以宽锐角至近直角变化;脉间区发育中等,形状不规则、大小略有变化,多为4边;盲脉一次分支,叶缘末级脉序汇聚于边缘(图2E)。
2.2.6 钩柄狸尾豆Uraria rufescens(DC.) Schindl. 叶长椭圆形,长宽比为1.69~2.35;主脉纤细,直向行走;二级脉14~19对,为直达叶缘型,其脉间距常不规则;间二级脉为复合型;主脉与二级脉夹角向基部递增,为窄锐角;三级脉与主脉夹角由主脉向叶缘以中锐角至宽锐角变化;脉间区发育差,形状大小不规则,5边以上;无盲脉,叶缘末级脉序汇聚于边缘(图2F)。
2.2.7 中华狸尾豆Uraria sinensis(Hemsl.) Franch. 叶倒卵形,长宽比为1.01~2.31;主脉纤细,直向行走;二级脉8~10对,为混合型或直达叶缘型,其脉间距常不规则;间二级脉为复合型;主脉与二级脉夹角向基部递增,为窄锐角;三级脉与主脉夹角由主脉向叶缘以窄锐角至宽锐角变化;脉间区发育中等,形状不规则,多为4边,偶尔出现5边;盲脉纤细一次分支,叶缘末级脉序汇聚于边缘(图2G)。
2.2.8 台湾蝙蝠草Christia campanulata(Wall.)Thoth. 叶长椭圆形,长宽比为1.53~2.22;主脉纤细,直向行走;二级脉12~18对,为直达叶缘脉或混合型脉型,其脉间距不规则;间二级脉为复合型;主脉与二级脉夹角向基部递增,为窄锐角;三级脉与主脉夹角由主脉向叶缘以中锐角至宽锐角变化;脉间区发育中等,形状不规则、大小略有变化,以4边居多;盲脉一次分支,叶缘末级脉序汇聚于边缘(图2H)。
2.2.9 长管蝙蝠草Christia constricta(Schindl.) T.Chen 叶近四方形,长宽比为0.79~0.99;主脉纤细,直向行走;二级脉4~6对,为环结曲行脉型,其脉间距规则;间二级脉为复合型;主脉与二级脉夹角向基部递增,为中锐角;三级脉与主脉夹角由主脉向叶缘变化稳定,为中锐角;脉间区发育良好,形状、大小一致,以4边居多;盲脉二次分支,叶缘末级脉序汇聚于边缘(图2I)。
2.2.10 铺地蝙蝠草Christia obcordata(Poir.) Bahn.F. 叶近圆形或倒三角形,长宽比为0.52~0.91;主脉纤细,直向行走;二级脉3~5对,为混合型脉或环结曲行脉型,其间距规则;无间二级脉;主脉与二级脉夹角稳定,为中锐角;三级脉与主脉夹角由主脉向叶缘以中锐角至宽锐角变化;脉间区发育良好,形状、大小较一致4边居多;盲脉纤细为不均等分支,叶缘末级脉序汇聚于边缘(图2J)。
2.2.11 蝙蝠草Christia vespertilionis(L.f.)Bahn.F. 叶倒三角形,长宽比为0.12~0.24;主脉纤细,直向行走;二级脉序3~4对,为直达叶缘脉型,其间距规则;间二级脉为复合型;主脉与二级脉夹角稳定,为宽锐角;三级脉与主脉夹角由主脉向叶缘以宽锐角至近直角变化;脉间区发育中等,形状、大小较一致4边居多;无盲脉,叶缘末级脉序汇聚于边缘(图2K)。
2.2.12 短序算珠豆Urariopsis brevissimaYang et Huang 叶心形,长宽比为1.04~1.22;主脉纤细,直向行走;二级脉序9~11对,为直达叶缘脉型,其脉间距规则;间二级脉为复合型;主脉与二级脉夹角向基部递减,为中锐角;三级脉与主脉夹角由主脉向叶缘以中锐角至宽锐角变化;脉间区发育中等,形状、大小较一致,以4边居多;盲脉一次分支,叶缘末级脉序汇聚于边缘(图2L)。
2.3 狸尾豆属及其近缘属叶脉序特征的主成分分析
由于主脉粗细、三级脉类型和叶缘末级脉序类型没有变异,不具有统计学意义。主成分分析前进行相关性分析,结果显示各性状之间的相关性较小,因此选用剩余性状进行主成分分析。主成分分析结果(表4)表明,前5个主成分的累计贡献率为84.587%,二级脉间距(0.860)、二级脉对数最小值(0.845)、二级脉对数最大值(0.841)、顶生小叶叶形(-0.832)、盲脉分支情况(-0.851)、顶生小叶长宽比(0.841)、三级脉与主脉角度变化(0.916)、顶生小叶宽(0.800)、二级脉与主脉夹角变化(0.885)负荷信息量大。
2.4 狸尾豆属及其近缘属的聚类分析
由图3可知,在组间欧式距离为20时,狸尾豆及其近缘属的12个物种聚为2个支系,狸尾豆属、算珠豆属、蝙蝠草属物种均没有各自聚为一个支系。其中,台湾蝙蝠草与狸尾豆属的物种聚为一个支系,短序算珠豆与其余蝙蝠草属的物种聚为一个支系。组间欧式距离为15时,12个物种聚为4个支系。长圆叶狸尾豆单独为一个支系,其余狸尾豆属物种与台湾蝙蝠草聚为一个支系;短序算珠豆与蝙蝠草聚为一个支系;长管蝙蝠草与铺地蝙蝠草聚为一个支系。组间欧式距离为5时,钩柄狸尾豆与滇南狸尾豆聚为一个支系,中华狸尾豆、猫尾草和台湾蝙蝠草聚为一个支系,其余物种各自独立为一个支系。
2.5 狸尾豆属及其近缘属叶脉序特征分种检索表编制
基于本研究对狸尾豆属及其近缘属12个物种叶脉序特征的比较分析,编制了如下分种检索表:
1.主脉与二级脉夹角小于45°,为窄锐角。
2.主脉与二级脉夹角变化相对一致。
3.二级脉对数小于或等于12对,盲脉不均等分支猫尾草Uraria crinita
3.二级脉对数大于12对,盲脉一次分支 长圆叶狸尾豆Uraria oblonga
2.主脉与二级脉夹角变化向基部递增。
4.盲脉一次分支……美花狸尾豆Uraria picta
4.盲脉二次分支、不均等分支或无盲脉。
5.二级脉对数大于或等于14对,脉间区发育差钩柄狸尾豆Uraria rufescens
5.二级脉对数小于14对,脉间区发育中等。
6.顶生小叶卵形,二级脉间距规则 狸尾豆Uraria lagopodioides
6.顶生小叶倒卵形或长椭圆形,二级脉间距不规则。
7.顶生小叶长大于9 cm,盲脉不均等分支 滇南狸尾豆Uraria lacei
7.顶生小叶长小于8 cm,盲脉二次分支 中华狸尾豆Uraria sinensis
1.主脉与二级脉夹角大于或等于45°,为宽锐角或中锐角(台湾蝙蝠草除外)。
8.顶生小叶长宽比大于或等于1,二级脉对数大于或等于9对。
9.顶生小叶长椭圆形,主脉与二级脉夹角變化向基部递增,二级脉间距不规则 台湾蝙蝠草Christia campanulata
9.顶生小叶心形,主脉与二级脉夹角变化向基部递减,二级脉间距规则 短序算珠豆Urariopsis brevissima
8.顶生小叶长宽比小于1,二级脉对数小于9对。
10.主脉与二级脉夹角大于45°并且小于65°,为中锐角。
11.顶生小叶近四方形,主脉与二级脉夹角变化向基部递增,盲脉二次分支 长管蝙蝠草Christia constricta
11.顶生小叶近圆形或倒三角形,主脉与二级脉夹角变化相对一致,盲脉不均等分支 铺地蝙蝠草Christia obcordata
10.主脉与二级脉夹角大于65°并且小于80°,为宽锐角 蝙蝠草Christia vspertilionis
3 讨论
叶脉序变异受基因和环境共同调配,叶脉变异在Sachs等[29]认为叶脉特征受生长素调节影响,叶片在幼叶阶段,基础细胞与脉细胞分裂过程中释放生长素,周围细胞吸收生长素到一定程度,产生高浓度生长素位点和浓度梯度,他们决定新脉生长方位;当叶脉主脉密度发育到一定的临界值,叶脉逐渐分为不同等级,最后密度不断降低直至呈现一致性[4],对探究植物属间或种间分类起到重要作用。
3.1 叶脉序特征在属间的分类学意义
叶脉序特征可以为植物近缘属间的分类提供依据,如解表木属Pilocarpus Vahl及其近缘属、杉龙丹属Lavoisiera DC.及其近缘属的二级脉走向、对数及类型、主脉与二级脉夹角、盲脉分支情况等叶脉序特征[18, 14]。狸尾豆属、蝙蝠草属和算珠豆属间在花和果实的形态特征上存在过渡特征,因此基于宏观形态特征的属间分类界限一直存在争议[7]。本研究结果表明叶脉序特征上的区别主要集中在顶生小叶叶形、二级脉对数及脉间距、二级脉与主脉夹角及夹角变化,这些特征是狸尾豆属及其近缘属有价值的属间分类特征。基于叶脉序特征进行的聚类分析表明,研究中涉及到的3个属的物种聚为2个单系支,其中狸尾豆属的物种与台湾蝙蝠草聚为一支,而其余蝙蝠草属物种与短序算珠豆聚为一支,本研究结果可以为今后狸尾豆属、蝙蝠草属和算珠豆属的分类修订提供参考。
3.2 叶脉序特征在属内种间的分类学意义
叶脉序特征可以为属内种间分类提供依据,如二级间脉类型、三级脉与较高级脉序类型、网眼发育情况以及盲脉的分支方式在柳属Salix L.具有重要的分类价值[19];网眼发育程度以及盲脉分支情况可以作为显脉木樨Osmanthus hainanensis与细脉木樨Osmanthus gracilinervis种间分类的依据[20];二级脉对数和主脉与夹角的大小、脉间区发育程度可以作为中间锦鸡儿Caragana liouana与拧条锦鸡儿Caragana korshinskii的种间分类依据[21];二级脉基部是否呈直线且相互平行是小花清风藤Sabia parviflora与四川清风藤Sabia schumanniana的重要区分特征[22]。本研究通过对叶脉序特征的观察和统计,发现狸尾豆属内种间变异主要集中在顶生小叶叶形、顶生小叶长宽比、二级脉对数、二级脉类型及间距、二级脉与主脉夹角变化、三级脉与主脉夹角变化、脉间区发育情况、盲脉分支情况。蝙蝠草属内种间变异主要集中在顶生小叶叶形、顶生小葉长宽比、二级脉对数、二级脉类型及间距、二级脉与主脉夹角及夹角变化规律、三级脉与主脉夹角变化、脉间区发育情况、盲脉分支情况。总体来看,顶生小叶叶形、顶生小叶长宽比、二级脉与主脉夹角及夹角变化、二级脉类型及间距、二级脉对数、三级脉与主脉夹角变化、脉间区发育情况、盲脉分支情况是狸尾豆属及近缘属种间分类的有价值的分类特征。
基于叶脉序特征的聚类分析结果显示,台湾蝙蝠草与狸尾豆属的物种具有较近的亲缘关系,其叶脉序特征与狸尾豆属植物较一致,主要体现在其顶生小叶叶形为长椭圆形,小叶长宽比1.53~2.22;二级脉12~18对,为直达叶缘型;二级脉与主脉夹角向基部递增,为窄锐角;间二级脉为复合型;脉间距不规则;发育中等,形状不规则,大小略有变化,4~5边。钩柄狸尾豆与滇南狸尾豆亲缘关系较近,其顶生小叶均为长椭圆形;二级脉直达叶缘型且脉间距不规则;二级脉与主脉夹角向基部递增,为窄锐角;间二级脉为复合型;三级脉与主脉夹角由主脉向叶缘以中锐角至宽锐角变化。中华狸尾豆、猫尾草和台湾蝙蝠草聚为一个支系,其二级脉间距均不规则;二级脉与主脉夹角,为窄锐角;间二级脉为复合型;脉间区发育中等,形状不规则,大小略有变化,4~5边。长圆叶狸尾豆单独聚为一支,其二级脉为真曲行;二级脉与主脉夹角变化相对一致;三级脉与主脉夹角由主脉向叶缘变化一致,为宽锐角。
短序算珠豆与蝙蝠草属物种聚为一支,显示出了较近的亲缘关系,其二级脉与主脉夹角均为中锐角,二级脉间距规则。其中,短序算珠豆与蝙蝠草亲缘关系近,其二级脉均为直达叶缘型;间二级脉为复合型;二级脉间距规则;脉间区发育中等,形状不规则,大小略有变化,4~5边。长管蝙蝠草与铺地蝙蝠草亲缘关系较近,铺地蝙蝠草顶生小叶长宽比0.52~0.91在长管蝙蝠草顶生小叶长宽比0.79~0.99的区间内;二级脉环结曲行脉;二级脉与主脉夹角,为中锐角;二级脉间距规则;发育良好,形状大小一致,多为4边。
3.3 叶脉序特征对于分类有争议物种的分类学意义
台湾蝙蝠草的分类长期存在争议。一些学者认为其体态极似狸尾豆属,应归为狸尾豆属[23];还有一些学者认为其花萼花后增大且荚果完全藏于宿存的萼内,应归为蝙蝠草属[2,7]。依《中国植物志》和Flora of China记载,台湾蝙蝠草隶属于蝙蝠草属,认为其表现出狸尾豆属与蝙蝠草属的中间过渡形态,证明这两个属间的密切亲缘关系[2,7]。从本研究中观察到的叶脉序特征来看,台湾蝙蝠草与狸尾豆属物种在顶生小叶叶形、顶生小叶长宽比、二级脉对数、二级脉与主脉夹角等方面的特征相似,而与蝙蝠草属物种明显不同。另外,基于叶脉序特征的聚类分析结果也表明,台湾蝙蝠草与狸尾豆属亲缘关系更近。因此,本研究结果支持将台湾蝙蝠草归入狸尾豆属。
长圆叶狸尾豆Uraria oblonga(Wall.ex Benth.)H.Ohashi & K.Ohashi是Ohashi等[24]2018年修订的物种。Ohashi等[24]基于ITS和9个叶绿体片段构建的山蚂蝗族的系统发育关系表明,山蚂蝗属并不是一个单系,长圆叶山蚂蝗Desmodium oblongum Wall.ex Benth.)与狸尾豆属的物种构成一个支持率很高的单系支,因此,将长圆叶山蚂蝗归入狸尾豆属,并命名为长圆叶狸尾豆Uraria oblonga。在本研究中,基于叶脉序特征的聚类分析结果表明,长圆叶山蚂蝗与狸尾豆属的物种聚为一个支系,支持Ohashi等[24]的分类修订。同时,聚类分析结果显示长圆叶狸尾豆与美花狸尾豆U.picta具有较近的亲缘关系,这两个物种的叶脉序特征的相似性主要在于顶生小叶长,二级脉类型为直达叶缘脉或真曲行脉,脉间距规则且二级脉对数相近,与主脉夹角均为窄锐角,三级脉与主脉角度由主脉向叶缘保持宽锐角变化,脉间区发育中等,盲脉均为一次分支等方面。
算珠豆属Urariopsis Schindl.由Schindler于1916年从狸尾豆属中分离出来,作为一个独立的属[25],仅包含算珠豆Urariopsis cordifolia(Wall.)Schindl.和短序算珠豆Urariopsis brevissima Yang et Huang 2个种。然而,算珠豆属的分类地位一直存在争议,一些学者认为算珠豆属荚果各结连接点在中央,由叶复叶退化为单小叶,演化水平高,应该作为一个独立的属[25-26,8,7];另外一些学者则认为算珠豆属形态与狸尾豆属相似,应该并入狸尾豆属[5,6,23,27,28]。本研究聚类分析结果显示算珠豆属与蝙蝠草属具有较近的亲缘关系,其二级脉间距规则、二级脉与主脉夹角多为中锐角、三级脉与主脉角度由主脉向叶缘以中锐角至宽锐角变化,与蝙蝠草属植物的叶脉序特征较一致。然而,由于算珠豆属植物在野外的分布记录少,种群小,采集困难,本研究仅包含了短序算珠豆一个物种,算珠豆属准确的分类地位还需要更全面的取样和分子系统学等方面的证据。
4 结论
综上所述,狸尾豆属及近缘属的叶脉序特征既变异丰富,又具有一定的一致性。叶脉序特征变异的丰富性主要体现在顶生小叶叶形、顶生小叶长宽比、二级脉与主脉夹角及夹角变化、二级脉类型及间距、二级脉对数、三级脉与主脉夹角变化、脉间区发育情况、盲脉分支情况等方面;一致性主要体现在中国狸尾豆属、蝙蝠草属和算珠豆属植物叶片均为纤细且直向行走的主脉,复合型的间二级脉(铺地蝙蝠草除外),结网型与贯穿型相结合的三级脉,汇聚于叶片边缘的末级脉本研究结果表明属间具有分类价值的叶脉序特征有:顶生小叶叶形、顶生小叶长宽比、二级脉对数及脉间距、二级脉与主脉夹角及其变化、三级脉与主脉夹角变化;种间具有分类价值的特征有:顶生小叶叶形、顶生小叶长宽比、二级脉类型、二级脉对数及间距、二级脉与主脉夹角、二级脉与主脉夹角变化、三级脉与主脉夹角变化、脉间区发育情况、盲脉分支情况。研究者发现造成叶脉序形态可塑性和变异性与阳光辐射、温度和养分供应有着密不可分的关系[29]。本研究为狸尾豆属及近缘属的分类提供了叶脉序特征的证据,为其作为药用植物资源的鉴定提供了参考。
参考文献:
[1]OHASHI H, IOKAWA Y, PHON P D.The genus Uraria (Leguminosae)in China[J].The Journal of Japanese Botany, 2006, 81(6): 332-361.
[2]杨衔晋,黄普华.国产狸尾豆属的研究[J].植物研究,1981,1(3):1-20.
[3]杨衔晋,黄普华.中国植物志[M].北京:科学出版社,1995,41:1-161.
[4]孙素静,李芳兰,包维楷.叶脉网络系统的构建和系统学意义研究进展[J].热带亚热带植物学报,2015,23(3):353-360.
[5]LEWIS G, SCHRIRE B, MACKINDER B, et al.Legumes of the world.Royal botanic gardens[J].Kew, Surrey, UK, 2005,433-446.
[6]OHASHI H, IOKAWA Y, PHON P D.The genus Uraria (Leguminosae) in China[J].The Journal of Japanese Botany, 2006, 81(6): 332-361.
[7]HUANG PH, OHASHI H, IOKAWA Y.UrariaDesvaux.In: Wu ZY, Raven PH eds.Flora of China, vol.10.[M].Beijing: Science Press, St.Louis: Missouri Botanical GardenPress,2010: 286-288.
[8]HICKEYL J.Casification of the architecture of dicotyledonous laves[J].American Journal of Botany, 1973, 60: 17-33.
[9]EILLS B, DALY D C, HICKEY L J,et al.Manual of leaf architecture[M].New York cornell University Press, 2009:7-123.
[10]李世晉,张奠湘,黄向旭,等.中国云实属植物叶脉形态学[J].热带亚热带植物学报,2004,12(2):133-141.
[11]KELLNER A, BENNER M, WALTHER H, et al.Leaf architecture of extant species of Rosa L.and the species Rosa lignitum Heer (Rosaceae)[J].Interational Journal of Plant Sciences, 2012, 173(3):239-250.
[12]SILVA D N T, LOPES D S A, LIMA D, et al.New records of fossil leaves from Abun Basin, Upper Pleistocene, Rondnia, Brazil[J].Historical Biology,2021, 33(12): 3454-3470.
[13]SUTAR S S, SALUNKE R J.Study of leaf venation in some species of genus Bauhinia L.[J].Phytochemistry, 2016, 5(4):122.
[14]MASUNGSONG L, BELARMINO M M.,BUOT J.Delineation of the selected Cucumis L.species and accessions using leaf architecture characters[J].Biodiversitas,2019, 20(3): 629-635.
[15]KADIRI A B, MUELLNERRIEHL A N.Comparative leaf micromorphology of Drypetes and Putranjiva (Putranjivaceae) and its taxonomic significance[J].Botanical Journalofthe Linnean Society,2021, 195(2): 139-160.
[16]喻诚鸿,陈泽濂.华南木本双子叶植物叶的宏观结构资料Ⅰ.术语与方法[J].中国科学院华南植物所集刊,1986,2:83-97.
[17]孙启高,宋书银,王宇飞,等.介绍双子叶植物叶结构分类术语[J].植物分类学报,1997(3):275-288.
[18]DO C A, PACIFICO R, GONCALESSILVA R, et al.Epidermal micromorphology and venation patterns of Microlicieae (Melastomataceae) leaves: Looking for new characters for the taxonomy of this neotropical tribe[J].Flora,2019, 261: 151494.
[19]孟祥玉,于慧莹,张志翔.国产柳属繁柳组植物叶脉序研究及其分类学意义[J].植物研究,2017,37(4):481-491.
[20]许炳强,夏念,王少平,等.中国木犀属植物叶脉形态及其分类学意义[J].广西植物,2007,27(5):697-705,696.
[21]江莎,暴艳,古松,等.内蒙古地区锦鸡儿属3种植物叶脉序比较研究[J].西北植物学报,2010,30(2):275-286.
[22]陆祥,孙庆文,郭文凯.清风藤属9种药用植物叶脉序研究[J].中药材,2019,42(5):1012-1017.
[23]DY P P.Uraria Desvaux.In: Morat Ph., Flore du Cambodge du Laos et du Viêtnam 23.Nguyenvan Thun, P Dy Phon et C Niyomdham, avec la colaboration de Y.Vidal, LégumineusesPapilionodées.Muséum National d′Histoire Naturele[M].Paris: Laboratoire de Phanérogamie, 1987, 95-116.
[24]OHASHI K, OHASHI H, NEMOTO T, et al.Phylogenetic analyses for classification of the Desmodium group of Leguminosae tribe Desmodieae 2.Two new genera separated from Desmodium and two new combinations in Grona and Sohmaea[J].The Journal of Japanese Botany, 2018, 93(5): 2018, 293-306.
[25]SCHINDLER AK.Desmodiinae novae[J].Englers Botanische Jahrbücher, 1916, 54: 51-53.
[26]VAN MEEUWEN MS, NOOTEBOOM HP, VAN STEENIS C.Preliminary revisions of some genera of Malaysian PapilionaceaeⅠ[J].Reinwardtia,1961, 5(4): 419-456.
[27]HUTCHINSON J.The genera of flowering plants.Dicotylédones vol.1[J].The Genera Of Flowering Plants, 1964, 1: 481.
[28]DE H, MTM B.Urariopsis reduced to Uraria(LeguminosaePapilionoideae)[J].Blumea, 1980, 26(2): 439-444.
[29]SACK L, SCOFFONI C, MCKOWN A D, et al.Developmentally based scaling of leaf venation architecture explains global ecological patterns[J].Nat Commun, 2012, 3: 837.
[30]杨庆锋,蔡雪珍,陈涛.长果安息香属和秤锤树属植物叶片脉序研究[J].广西植物,1997,17(2):145-148.
[31]LU H, JIANG W, GHIASSI M, et al.Classification of Camellia(Theaceae) species using leaf architecture variations and pattern recognition techniques[J].PLoS ONE, 2012, 7(1): e29704.
[32]邵文,楊鲁红,周喜乐.石韦属植物脉序特征及其分类学意义[J].植物科学学报,2016,34(2):191-199.
[33]田昌芬,李蒙,黄亚健,等.中国花楸属单叶类群叶脉序特征研究[J].广西植物,2022,42(1):122-132.
(责任编辑:柯文辉)
