早花象牙参为母本的3个种间杂交F1表型多样性研究
2023-05-30高玉红胡秀毕雨欣伊力孜亚·特尼斯别克牟凤娟
高玉红 胡秀 毕雨欣 伊力孜亚·特尼斯别克 牟凤娟
关键词:早花象牙参;F1代;种间杂交:表型多样性
中图分类号:S567.01 文献标识号:A 文章编号:1001-4942(2023)03-0030-08
象牙参属(Roscoea)植物为姜科(Zingiberace-ae)中较为耐寒的类群,适应于高山寒温带气候。该属植物全世界有20余种,主要分布于印度、尼泊尔、克什米尔以及我国西南地区。我国共有该属植物14种和3变种,其中云南有11种和2变种,四川有8种和1变种,西藏有5种,是我国象牙参属植物的主要产区。我国西南部高山地区象牙参属植物资源丰富,值得进一步深入研究和开发利用。该属植物为多年生小草本,肉质根簇生、纺锤状,宿存过冬;叶2列,3~6片,条形或圆形,长8~25 cm,宽2~4 cm,无叶柄;花为白色、紫色、黄色及红色。该属植物多以块根入药,因块根在极度缩短的根茎上呈放射状排列,形如鸡爪,民间习称土闻中、鸡爪参、鸡脚参、佛掌参等,西南地区常作藏药和民间草药用,有补肺定喘、温中散寒、止痛消食等功效。如:四川会理、会东等地常用长柄象牙参(R.debilis)医治肾炎;藏象牙参(R.tibetica)在西藏可用于温中散寒、止痛消食;云南大理、丽江等地将多种象牙参属植物用于益气补血,治疗咳嗽、心累等。该属植物花形酷似兰花,美丽且花色多样,具有较高观赏价值,许多种类适合作小盆栽或花坛配置的优良素材。早在1920年,英国皇家园艺学会(RHS)就曾授予大花象牙参(R.humeana)最高奖,1932年再被高山植物园学会(AGS)授予最高奖。欧美国家19世纪就已经栽培象牙参属植物以供观赏,现已培育出多个栽培品种并广泛应用于盆栽观赏和庭院美化,而在我国象牙参属植物完全处于野生状态,尚未对其进行开发利用。
目前,国内外对于象牙参属植物的研究主要涉及分类学、解剖学、离体培养、传粉生物学以及植物资源调查方面。欧云锋等研究发现,高山象牙参(R.alpina)是自交为主兼具异交的混合交配体系:Du等依据形态学和分子生物学信息研究证明大花象牙参(R.humeana)和早花象牙参(R.cautleoides)的种间自然杂交存在于同域群体中:杜光辉等通过HAT-RAPD方法对象牙参属植物的自然杂交进行个体鉴定,发现疑似杂交个体的形态表现介于早花象牙参和大花象牙参之间,并与大花象牙参的亲缘关系更接近。上述研究结果可为象牙参属植物进行种间远缘杂交提供理论依据。象牙参属与姜花属(Hedychium)的亲缘关系较近,姜花属的相关研究结果可为象牙参属植物种间杂交和种质创新研究提供参考依据。目前有不少关于姜花属的种间杂交报道:Wood等研究了野外姜花属植物的花部特征,发现其自然居群疑似杂交类群,暗示姜花属植物在野外可以进行自然杂交且发生频繁:姜花属植物进行种间杂交较易成功;Yost等发现花粉粘附性、萌发力和花粉管生长的降低会导致姜科闭鞘姜属(Costus)两种密切相关的同域物种间生殖隔离。
遗传多样性是研究物种多样性以及生态系统多样性的基础,是物种进化的本质。植物作为生态系统中的生产者,其遗传多样性水平对于生态系统功能调节有着举足轻重的作用。植物种群的遗传变异和维持变异的机制可以通过遗传多样性来体现。许多研究者已对多种植物杂交后代的遗传多样性进行研究,如王丽萍等[29]发现冬油菜和春油菜的正交或反交F,代均以冬性表现为主,证明植物的抗寒性受多基因数量遗传控制;王东雪等对油茶(Camellia oleifera)F1代叶表型遗传多样性的研究表明,F1代存在较高的遺传变异和遗传传递力:王永淇等对姜科植物新品种“红丰收山姜”的遗传变异研究表明,该材料的变异较大。本研究对象牙参属的早花象牙参与其它3个种的杂交F1代的表型变异进行分析,以期为象牙参属植物种质资源的创新及开发利用提供参考。
1材料与方法
1.1试验材料
2019年6月至11月,以西南林业大学苗圃引种栽培的早花象牙参为母本,以粉叶象牙参(Roscoea glaucifolia)、绵枣象牙参(R.scillifolia)、长柄象牙参(R.debilis)为父本进行种间杂交,获得“早花×粉叶”“早花×绵枣”“早花×长柄”3个种间杂交F1代种子各105、78、95粒。
1.2试验方法
2020年6月将3个种间杂交F1代种子各60粒浸水催芽后播种。2020年整个生长期内观测3个种间杂交F1一年生幼苗的各项表型指标,待2021年3月挖出每棵植株的宿根,再测定其块根各项表型指标,分析叶和块根表型的遗传多样性。由于播种时间较晚,F1一年生植株当年未开花。为获得其表型多样性的完整数据,于2021年继续观测由宿根萌芽的二年生植株的各项表型特征,并记录开花情况。
株高:于10月15日(七叶一心)使用卷尺测量株高。
叶表型:于10月15日(七叶一心)记录每棵植株的叶片数,用游标卡尺测量每片叶的长度和宽度,并计算叶形指数(叶片长/叶片宽)。
块根表型:于2021年3月1日挖出块根后,用电子天平称取各植株块根重,记录每棵植株的块根数,用游标卡尺测量每个块根的长度及最粗处直径。
花表型:于2021年植株开花时,选3个种间杂交F1各20株,每株调查2朵花,记录兜状花冠裂片、侧生花冠裂片、侧生退化雄蕊以及唇瓣数,分别测量其长度和宽度,并记录花冠颜色特征。
1.3数据处理与分析
每个材料各表型数据均以平均值作为统计单元。利用Microsoft Excel、SPSS软件进行数据处理与分析。
2结果与分析
2.1 3个杂交组合Fi群体一年生苗的表型分析
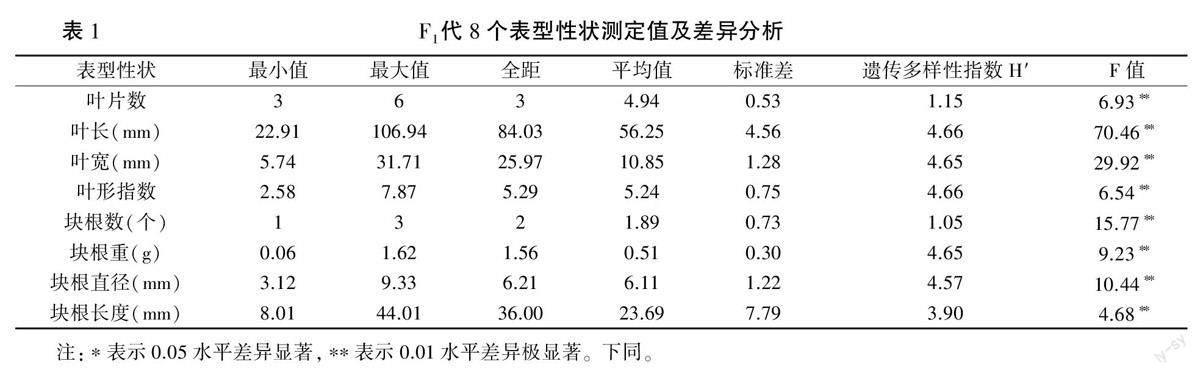
2.1.1 F1群体8个表型性状测定值及差异分析由表1可知,杂交象牙参F1群体存在丰富的遗传变异。其中,叶长为22.91~106.94 mm,平均56.25mm;叶宽为5.74~31.71mm,平均10.85 mm;叶形指数为2.58~7.87,平均5.24;块根重为0.06~1.62 g,平均0.51 g;块根直径为3.12~9.33 mm,平均6.11 mm;块根长度为8.01~44.01
mm,平均23.69 mm。8个表型性状的遗传多样性指数为1.05~4.66,且在3个杂交群体间和群体内均存在极显著差异(P<0.01),表明以早花象牙参为母本的种间杂交F1代遗传变异水平较高。
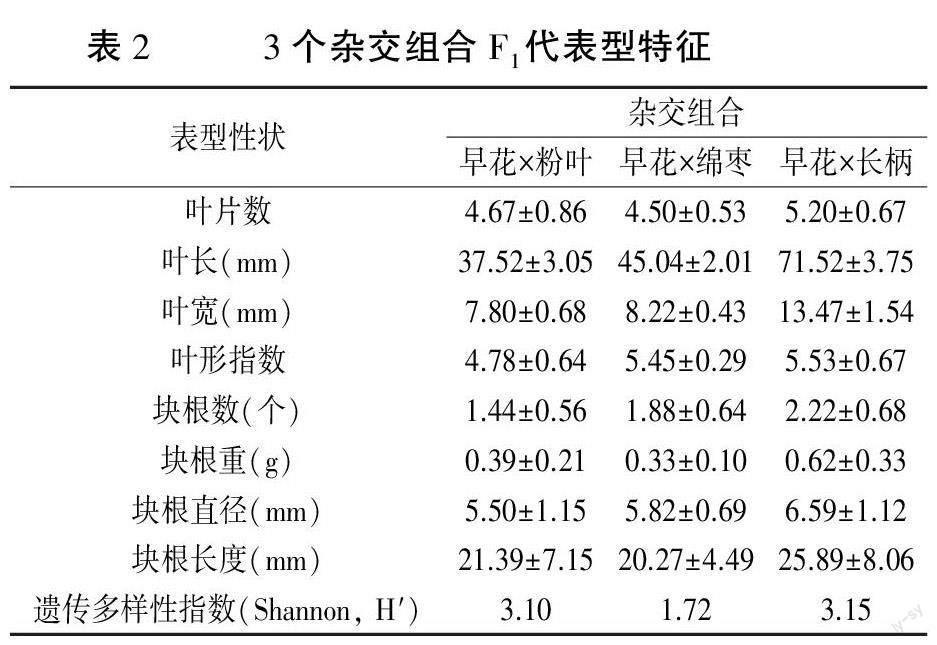
2.1.2 3个杂交组合的F1表型比较分析 3个杂交组合F1代表型特征见表2。从叶表型来看,叶片数、叶长、叶宽、叶形指数值均为“早花×长柄”F1最大,“早花×粉叶”F1(除叶片数外)最小,说明“早花×长柄”F1的叶片较发达;从块根表型来看,块根数、块根重、块根直径、块根长度的值均为“早花×长柄”F1最大,“早花×粉叶”F1块根数量最少、直径最小。遗传多样性指数表明,“早花×粉叶”和“早花×长柄”的多样性水平较高,而“早花×绵枣”的遗传变异最小。综合上述分析,“早花×长柄”F1叶和根表型均表现较好,该组合具有选育观赏用和药用早花象牙参优良品种的潜力。
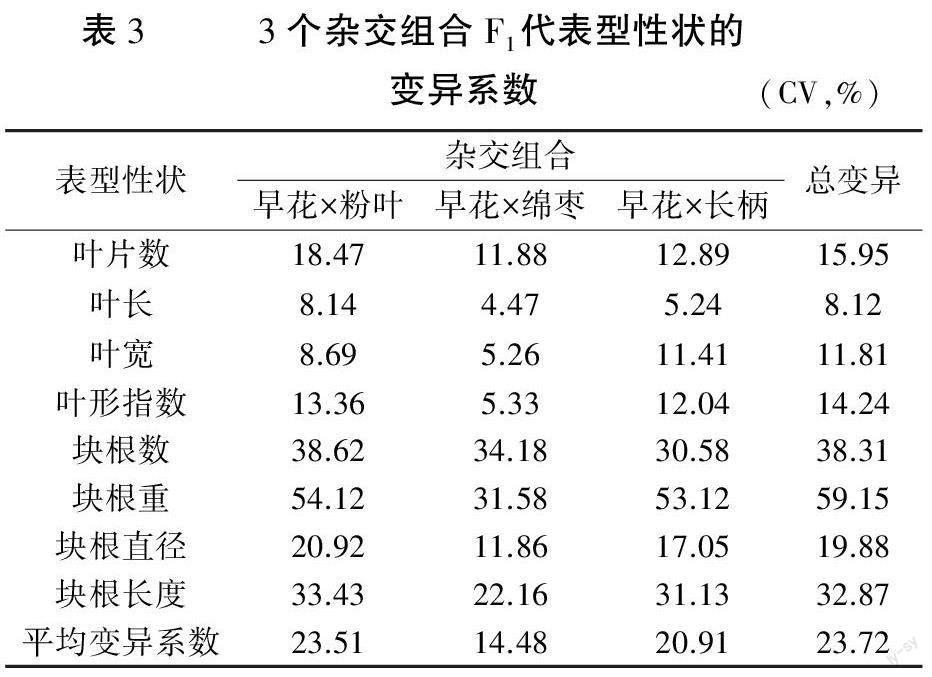
2.1.3 F1群体8个表型性状的变异分析 以早花象牙参为母本的3个不同杂交组合F1群体表型性状的变异系数见表3。各杂交组合F1群体内表型的变异系数介于4.47%~54.12%之间,最高的为“早花×粉叶”块根重,其次是“早花×长柄”块根重(53.12%),最小的为“早花×绵枣”叶长。3个杂交群体内和群体间叶长的变异系数均最小,群体内分别是8.14%、4.47%和5.24%,群体间为8.12%,表明叶长与其它表型相比表现最为稳定,可将其作为判别杂交亲本特别是母本来源的有效指标。
F1群体中8个表型性状的平均变异系数为23.72%,其中,根表型变异相对较大,变异系数为19.88%~59.15%,变异最大的是块根重,最小的是块根直径,表明不同杂交群体的块根重差异较大;而叶表型变异相对较小,变异系数为8.12%~15.95%,变异最大的是叶片数,最小的是叶长(8.12%)。根据变异系数的分级梯度可知,叶表型的变异程度为低级或中级,根表型的变异程度为中级或高级,表明以早花象牙参为母本的杂交F1代中,根表型遗传变异水平均高于叶表型。
3个杂交群体F1表型的平均变异系数从大到小排序为“早花×粉叶”(23.51%)>“早花×长柄”(20.91%)>“早花×绵枣”(14.48%),表明在这3个杂交组合中,“早花×绵枣”组合的F1群体多样性水平最低。
2.1.4 F1群体8个表型性状的相关分析 由表4可见,3个杂交组合F1代群体的叶表型和根表型之间存在不同程度的相关性。其中,叶片数与叶形指数间为显著相关,与其它6个表型性状为极显著正相关;叶长与块根长度为显著正相关,与其它表型均为极显著正相关;叶宽与块根的数量、直径、长度之间均为极显著正相关;块根的长度和直径与块根重之间无显著相关性。
2.2 3个杂交组合F1群体二年生苗的表型分析
2.2.1 F1代二年生苗花朵表型差异
由表5可知,3个杂交组合F1群体二年生苗的开花率为“早花×长柄”(60.78%)>“早花×粉叶”(8.33%)>“早花×绵枣”(0),“早花×长柄”F1有4种花色,且花朵结构发生变异。表明3个杂交群体中“早花×长柄”后代的花色和花朵结构变异最丰富。鉴于此,可将其作为选育观赏类象牙参的种质来源。
2.2.2 F1群体二年生苗表型性状测定值及差异分析 3个杂交组合F1群体二年生苗主要叶表型和花表型的测定值及差异分析见表6和表7。由F值可知,叶表型性状的差异达显著或极显著水平,而花表型性状的差异不显著。F1群体二年生苗株高为70.02~240.01mm,差异显著,其中“早花×长柄”的植株较高,“早花×粉叶”的植株较矮;叶长为53.60~207.50 mm,平均126.59 mm;叶宽为6.75~26.20nm,平均16.91 mm;叶形指数为3.83~13.69,平均7.85。上述二年生苗的表型值与一年生苗相比,均有一定程度的增大,其中叶长增长最多,平均增大125.25%。
2.2.3 F1群体二年生苗表型变异分析 由表8可见,F1杂交群体二年生苗中,叶表型的总变异系数平均值为13.22%,而花表型的總变异系数平均值为11.18%。叶表型性状的总变异系数为5.17%~23.42%,其中变异最大的是叶形指数,最小为叶长,且叶长在所有表型性状中变异系数最小,说明象牙参属植物叶长表现最稳定,可将其作为杂交来源鉴定的主要依据之一。花表型性状的总变异系数为6.31%~20.49%,变异最大的是唇瓣宽,最小的为兜状花冠裂片长。
2.2.4 F1群体表型典则判别分析 结果(表9)显示,2个典则变量间存在极显著相关(P<0.01),且2个典则函数特征值占总变异的100%,能较好地体现F1群体的分类特征。
标准化后的典则判别函数系数见表10。可知,对第1典则变量贡献率最大的是叶形指数,对第2典则变量贡献率最大的是叶长,其余表型对典则变量也有不同程度的貢献。
典则判别函数散点图见图1。可以看出,3个杂交组合F1表型在两个函数坐标系内可以区分出来,但是各表型区内组群间分化不明显,其中“早花×粉叶”F1表型主要分布在第二、第三象限,“早花×绵枣”F1表型主要分布在第二象限,“早花×长柄”F1表型主要分布在第一、第四象限。这可能与父母本是同属不同种有关。
3讨论与结论
植物表型的差异是其内部遗传物质与环境互作在外部形态上的反映。本研究以早花象牙参为母本、3个同属不同种象牙参为父本组配杂交组合“早花×粉叶”“早花×绵枣”“早花×长柄”,并以其F1代为研究对象,分析其F1表型的遗传多样性及变异规律。结果表明:杂交组合“早花×长柄”的F1后代遗传了母本和父本的优良性状,块根较大、叶片较宽、花朵变异丰富,可考虑作为选育观赏和药用象牙参的优良组合:而“早花×粉叶”和“早花×绵枣”的F1代表型与其父本更接近,表现为块根较小、叶片较窄。有研究发现油茶F1代叶表型的变异幅度极大,最大值和最小值之比为2.42~9.08,变异系数为2.21%~37.50%,遗传传递力大于100%。本研究中,不同杂交组合F1代群体均表现出丰富的遗传变异:各杂交群体一年生苗表型性状的变异系数为4.47%~54.12%,二年生苗表型性状的变异系数为0.27%~30.54%:二年生苗各表型性状较一年生苗均有一定程度的增大,其中叶长增加最多,平均增大125.25%。综合来看,叶长与其它表型相比变异系数最小,表现最为稳定,可将其作为判别杂交亲本来源的可靠指标。本研究中F1种群的花色变异丰富,表现为数量性状遗传特征,这与‘香水柠檬×白花柠檬F1代花色表型分离及‘白姜花×金姜花F1花色遗传分析结果相似。表明可以依据部分性状表现对植物种质资源杂交后代进行早期间接评价选择,为高海拔地区象牙参属植物育种提供参考依据。
本研究中,以早花象牙参为母本进行种间杂交的结实率较高,说明其远缘杂交亲和性较高,这与对古老月季的研究结果一致。对叶表型和根表型的研究表明,3个杂交组合F1群体间和群体内均存在显著和极显著差异,且不同杂交组合间叶表型的变异比根表型大;花表型的研究表明,3个杂交组合F1间花朵大小无明显差异,但花朵结构、花色以及开花率存在明显区别,均表现为“早花×长柄”F1的多样性最高,而“早花×绵枣”F1代二年生苗暂时未开花。表明象牙参属不同种之间可以进行远缘杂交,但“早花×绵枣”F1代二年生苗未开花是否为生长年限不足或是基因交流受阻,还需进一步观察和深入研究。
我国象牙参属植物资源较丰富,但目前国内极少对其进行开发利用,更缺乏遗传多样性和遗传育种等相关研究。本研究通过探索象牙参属种间杂交后代的表型多样性和相关分析,可为今后象牙参属植物种质资源的创新和利用提供技术基础,还可为其它姜科植物种质资源的创新提供一定参考。
