桂林岩溶石山青冈优势种群空间分布格局及关联分析
2023-05-30盘远方卓文花姜勇梁士楚李丽香
盘远方 卓文花 姜勇 梁士楚 李丽香


摘 要: 種群空间分布格局是研究种群特征、种间相互作用以及种群与环境关系的重要手段。青冈(Cyclobalanopsis glauca)是岩溶生态系统顶极群落的建种群,为进一步了解其种群空间分布格局特征,该研究根据样地调查规范对面积为60 m × 60 m样地中的木本植物进行空间定位且进行每木调查,并运用点格局方法中的g(r)和L12(r)函数对青冈幼苗(DBH<2 cm)、幼树(2 cm≤DBH<10 cm)和成树(10 cm≤DBH)进行空间分布特征和关联性分析。结果表明:(1)青冈种群的径级结构符合倒“J”型分布。(2)幼苗、幼树和成树在较小尺度上表现为聚集分布,而幼树和成树在中、大尺度上基本表现为随机分布。(3)不同径级个体空间关联性主要表现为幼苗与幼树在0~15 m尺度上呈正相关;幼树与成树在0~10 m尺度上呈不相关,而在10~15 m尺度上呈正相关;幼苗与成树在0~2 m尺度上呈不相关,而在2~15 m尺度上呈正相关。因此,青冈种群的分布格局及其关联性随空间尺度和发育阶段而变化,这是由种群自身的生物学特性、种内种间关系以及环境条件共同作用的结果。该研究结果为该区域植物群落形成和维持机制及资源利用研究提供了基础资料。
关键词: 青冈种群, 径级结构, 空间点格局, 空间关联性, 岩溶石山
中图分类号: Q948.15
文献标识码: A
文章编号: 1000-3142(2023)03-0527-09
Spatial distribution pattern and correlation analysis
of Cyclobalanopsis glauca dominant population
in karst hills of Guilin
PAN Yuanfang2, ZHUO Wenhua1,3, JIANG Yong1,3, LIANG Shichu1,3, LI Lixiang1,3*
( 1. Key Laboratory of Ecology of Rare and Endangered Species and Environment Protection, Ministry of Education, Guangxi Normal
University, Guilin 541006, Guangxi, China; 2. Guangxi Mangrove Research Center, Guangxi Academy of Sciences, Beihai 536000,
Guangxi, China; 3. College of Life Sciences, Guangxi Normal University, Guilin 541006, Guangxi, China )
Abstract: The spatial distribution pattern of population is an important means to describe population characteristics, interspecific interactions, and the relationship between population and environment. Cyclobalanopsis glauca is a constructive population of climax community in karst ecosystems, in order to further understand the spatial distribution pattern of its populations, according to the sample plot investigation specification, a sample plot of 60 m × 60 m was set up, and the woody plants in the plot were spatially located and investigated for each tree. The C. glauca population was divided into three different diameter classes in the study, belonging to seedling (DBH<2 cm), sapling (2 cm≤DBH<10 cm), adult tree (10 cm≤DBH). Using the point pattern analysis method, g (r) and L12 (r) functions based on Ripleys K function were used for the analysis of the spatial distribution pattern and correlation analysis of C. glauca population in different diameter classes. The results were as follows: (1)The diameter class structure of C. glauca population was in an inverted “J” type distribution. (2) On the small scale, the stage of seedlings, saplings and adult trees, the spatial distribution pattern tended to cluster distribution,and the saplings, adult trees tended to random distribution on the medium and large scales. (3) The performances of spatial correlation between different diameter classes were different, the seedlings and saplings were positively related on the scale of 0-15 m; saplings and adult trees reflected on the scale of 0-10 m were no correlation, and there was a positive correlation on the scale of 10-15 m; seedling and adult trees reflected on the scale of 0-2 m were no correlation, and there was a positive correlation on the scale of 2-15 m. Therefore, the spatial distribution pattern and correlation of C. glauca population in the karst area of Guilin change with the spatial scale and development stage, which was the result of the combined effects of the populations biological characteristics, intraspecific and interspecific relationships, and environmental conditions. The results of the study can provide basic information for the formation and maintenance mechanism of plant community and resource utilization research of karst hills.
Key words: Cyclobalanopsis glauca population, diameter class, spatial point pattern, spatial correlation, karst hills
某一特定时间组成种群所有植株个体在二维空间上的分布状况或布局称之为种群空间分布格局(Cao et al., 2003; 高福元等, 2012),通常在群落中有随机分布、聚集分布和均匀分布3种基本类型。种群空间分布格局是分析种群与环境关系的重要手段,对解释种群本身生态学过程、结构发展历史和环境变迁具有重要意义(Hubbell, 2001; 张金屯等, 2004; Zhao et al., 2010)。有研究表明,种群空间分布格局与尺度有关(Harms et al., 2001; Wiegand & Moloney, 2004),在某一尺度上可能呈随机分布或聚集分布,而在另一尺度上可能呈均匀分布(Stoyan & Penttinen, 2000)。种群空间分布格局除与尺度有关外,还受到扩散限制(dispersal limitation)和生境异质性(habitat heterogeneity)的影响,在较小的尺度上可能受种内竞争、种间竞争、扩散限制等因素影响,而在较大尺度上则可能由种群分布区环境的异质性决定(Harms et al., 2001; Lin et al., 2011)。
目前,国内外学者已在种群空间分布格局和关联方面做了不少研究。例如,Ripley(1981)利用K函数的单因素和双因素函数分析个体的空间分布格局;Call和Nilsen(2003)采用Ripleys K函数分析美国布莱克斯堡2个优势种的空间关联性;Schleicher等(2011)运用Ripleys函数研究热带草原不同土地类型对2种灌木种群生长和空间分布的影响;苏松锦等(2014)运用O-ring函数分析戴云山黄山松种群空间分布格局和关联;Zhao等(2010)采用Ripleys K函数分析常绿阔叶林优势种的空间分布格局。但是,目前文献资料显示均以大尺度研究为主,而小尺度研究较为缺乏。宋于洋等(2010)研究认为,小尺度种群空间分布格局与关联是揭示种群生态学过程的关键尺度,即小尺度范围内的种群空间分布格局和关联分析结果更能准确地从生物学背景上提供和解释植物个体周围的局部空间结构。因此,研究小尺度种群空间分布格局和关联性对深入理解种内种间的相互作用、群落组成及其形成和维持机制具有重要意义。
桂林位于广西东北部,是广西岩溶石山地貌聚集地之一,也是我国石漠化水土治理和植被恢复的重点区域(刘彦随等, 2006)。其典型的岩溶生态特征表现为岩石裸露率高、土壤贫瘠、土壤少且不连续和地表水缺乏等,其脆弱的生态系统备受全球关注(王明章, 2004; 胡芳等, 2018)。该生态系统的植被是在碳酸盐类岩石风化壳发育形成的钙质土壤中生长的,植被形成时间十分缓慢(Zhang et al., 2013; 徐艳芳等, 2016)。若该生态系统遭到破坏,其植被容易退化且难以修复。独特的岩溶地质背景和高异质性的生境类型,形成了特有的植物区系组成及结构,这对维持岩溶生态系统的稳定极为重要。但是,岩溶石山的地形、植被组成和研究手段等是限制岩溶石山野外调查的重要因素。目前,关于岩溶石山植物种群空间格局方面的研究远不如非岩溶石山植被的深入。青冈(Cyclobalanopsis glauca)是我国南方亚热带地区常绿落叶阔叶混交林的主要树种之一,是岩溶生态系统顶级群落的建群种,具有良好的抗旱性、喜钙性、耐贫瘠性和发达根系等特点,在防止岩溶石山石漠化、维持脆弱生态系统稳定、调节区域碳平衡和维持物种多样性等方面具有重要的生态学意义(Hu et al., 2007, 2017; Li et al., 2008)。鉴于此,本研究以桂林岩溶石山青冈优势种群为对象,采用空间点格局方法中的g(r)函数和L12(r)函数,拟探讨幼苗、幼树和成树的空间分布格局变化,以及幼苗、幼树和成树之间存在的关联性,以期为岩溶石山森林资源的保护、营林管理、生物多样性保护以及岩溶石山退化植被生态系统修复提供科学依据。
1 研究区域与研究方法
1.1 研究区域概况
本研究区域位于桂林市郊的演坡山(110°15′17″—110°15′19″ E、 25°19′7″—25°19′10″ N),为典型的岩溶石山地貌,海拔大多在200~400 m。该区域岩溶地貌特征显著,地貌类型以碳酸盐溶蚀所形成的峰丛洼地和峰丛谷地为主,具有成土速度缓慢、土壤钙含量高、地表土蓄水能力差及环境容纳量低等特点;属于亚热带湿润季风气候,气候温和,雨量充沛,年均降雨量为1 856.7 mm,降雨量年分配不均,降雨主要集中在4—7月,年均蒸发量为1 458.4 mm,年均气温为19 ℃,无霜期长达300 d,全年以偏北风为主,平均风速为2.7 m·s-1。土壤主要为红壤、棕壤等砂质土。典型植被以壳斗科的青冈为主,其伴生种主要有檵木(Loropetalum chinense)、干花豆(Fordia cauliflora)、扁片海桐(Pittosporum planilobum)、粗糠柴(Mallotus philippensis)、紅背山麻杆(Alchornea
trewioides)、朴树(Celtis sinensis)等。
1.2 试验设计和调查方法
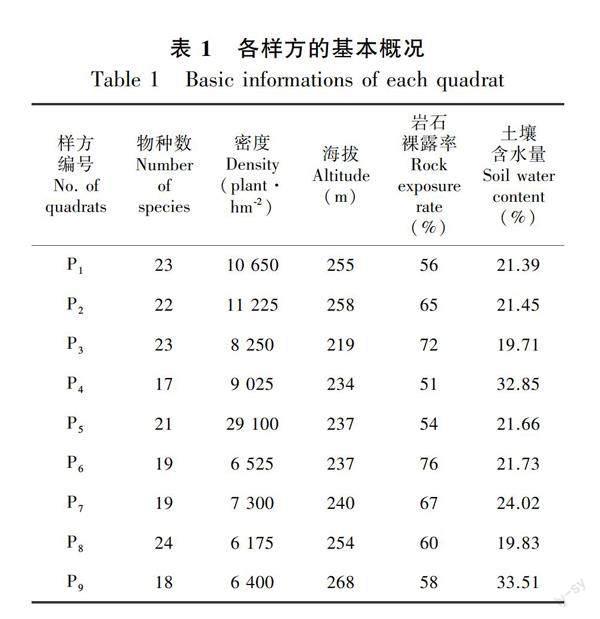
2017年7—9月,经过实地踏勘,结合青冈群落特点及生境情况,选取最具有代表性且分布较集中的青冈群落作为研究对象。但是,由于受到地形和植被的限制,即岩溶石山山体整体偏小,山体中上部位较为陡峭;在样地踏查过程中,发现山体中上部位较少有青冈的分布,主要分布着檵木、龙须藤和金樱子等藤本和小灌木,而青冈主要分布在山底至山中部位。因此,结合青冈的分布情况和地形条件确定以110°15′17″ E、25°19′7″ N 为样地原点坐标,设置一个面积为60 m × 60 m的样地。虽然整个研究样地相对偏小,但这是目前找到保存相对完好、原生性较强、生长较为集中的青冈种群样地。为了便于野外调查,采用相邻格子调查方法将整个样地分成9个20 m × 20 m的样方,各样方的基本概况如表1所示。调查时对每个20 m × 20 m的样方进行每木检尺,记录样方内胸径≥1 cm乔木物种和基径≥1 cm灌木物种的种名、胸径、基径、株高及生长状况,并测量每个植株的空间位置坐标。同时,记录每个20 m × 20 m样方的经纬度、坡向、海拔、岩石裸露率等环境特征。
1.3 径级划分
不同物种其径级的划分标准不相同,需根据物种的生物学特性和生长状况进行划分(何增丽等, 2017)。生长在相似立地条件下相同物种的龄级和径级对环境的反映规律具有一致性(任毅华等, 2021)。因此,该研究采用空间代替时间的方法,将青冈调查数据(样地中,青冈的平均胸径为8.63 cm,最大胸径为47.72 cm)划分为3个径级阶段,即幼苗的胸径DBH<2 cm;幼树的胸径为2 cm≤DBH<10 cm;成树的胸径10 cm≤DBH,以不同径级结构代替年龄结构分析青冈种群不同年龄结构的空间分布格局。虽然这样划分径级并不能非常科学准确地定义幼苗、幼树和成树,但有利于分析的简化。
1.4 数据处理和分析
1.4.1 不同径级青冈的空间格局 L(r)函数和 g(r)函数是由Ripleys K函数衍化而来(Ripiey, 1977),即以Ripleys K函数为基础,通过对Ripleys K函数进行方差校正和线性化得到L(r)函数,运用 L(r)函数来判断不同径级个体在各尺度上的空间分布格局。但是,L(r)函数为积累分布函数,在分析大尺度的空间分布格局时容易受到小尺度积累效应的影响,使得格局分析复杂化(王鑫厅等,2012,2022)。g(r)函数用环代替了Ripleys K函数中的圆,它利用两圆心之间的距离,计算任一点为圆心、r为半径的圆环区域内点的数量来进行点格局分析,计算过程中消除了累积效应(郭屹立等, 2015)。因此,本研究使用g(r)函数来描述青冈种群不同径级个体在各空间尺度上的分布格局。其中,Ripleys K函数、L(r)函数和g(r)函数公式如下:
Kr=An2∑ni=1∑nj=1Iruijwiji≠j;
Lr=K(r)/π-r; gr=dK(r)2πrdr。
式中:A表示样地面积;n为样地内个体总数;r表示空间尺度;wij为消除边界效应的权重;uij为点i到点j之间的距离;I为指示函数。当g(r)=0时,个体在r尺度上呈完全空间随机分布;g(r)<0时,为均匀分布;g(r)>0时,为聚集分布。
1.4.2 不同径级青冈的空间关联性 在分析物种间的空间关联时,Diggle(1983)在Ripleys K函数基础上引入了K12(r)函数。同样,对K12(r)函数进行方差校正和线性化得到L12(r)函数。因此,本研究利用分析物种间空间关联的方法来分析青冈种群不同径级间的空间关联。其中,K12(r)函数和L12(r)函数公式如下:
K12r=An1n2∑n1i=1∑n2j=1Ir(dij)wij(i≠j);
L12r=K12(r)/π-r。
式中:n1、n2表示两个不同径级的青冈植株个体数;i和j表示两个不同径级的植株。L12=0时,表明两个径级植株之间相互独立或无显著相关性;L12<0时,表明两个径级的植株在r尺度上呈显著负相关;L12>0时,表明两个径级的植株在r尺度上呈显著正相关。
本研究采用完全空间随机模型(complete spatial randomness, CSR),使用Monte Carlo进行100次的随机模拟,利用模拟得出的最大值和最小值来计算上下包迹线分布的估计范围,并根据其分布的区间截取95%的置信水平。若实际分布的g(r)值和L12(r)值落在包迹线内,则青冈呈随机分布或两径级之间相互独立,无空间相关性;若在包迹线以上,则为聚集分布或两径级之间呈空间正相关;若在包迹线以下,则为均匀分布或两径级之间呈空间负相关。数据分析过程在R3.6.3软件中的spatstat软件包中完成。
2 结果与分析
2.1 青冈种群及其所在群落的空间分布
如图1和表2所示,本次调查共记录到DBH≥1 cm的植株个体4 384株,隶属于29科43属50种,密度为12 177株·hm-2,平均胸径为6.12 cm,幼苗、幼树和成树的个体数分别是2 982、964、438株,分别约占总个体数的68.1%、21.9%和9.9%。其中,青冈个体2 239株,密度为6 219株·hm-2,种群平均胸径为8.63 cm,幼苗、幼树和成树的个体数分别是1 540、370、329株,分别约占青冈总个体数的68.8%、16.5%和14.7%。从径级分布趋势可以看出,样地内有大量的小径级个体储备,个体数量随径级的增大而逐渐减少,种群的径级结构更接近于倒“J”型,种群属于增长型,种群的更新状态良好。
2.2 不同径级青冈种群的空间分布格局
如图2所示,幼苗在0~15 m尺度上表现为聚集分布;幼树在0~5.5 m、8~10 m和12~15 m尺度上表现为聚集分布,在5.5~8 m和10~12 m尺度上表现为随机分布;成树在0~4 m尺度上表现为聚集分布,在4~15 m尺度上以随机分布为主。
不同径级的青冈种群在小尺度上均表现为聚集分布,除幼苗外,幼树和成树在较大尺度上主要表现为随机分布。
2.3 不同径级青冈种群的空间关联
如图3所示,幼苗与幼树在0~15 m尺度上表现为正相关;幼树与成树在0~10 m尺度上表现为不相关,在10~15 m尺度上表现为正相关;幼苗与成树在0~2 m尺度上表现为不相关,在2~15 m尺度上表现为正相关。
3 讨论
3.1 青冈种群的径级结构
种群的径级结构是指植株按径级大小的分配状况或各径级大小在种群内所占据的比例(拓锋等, 2020)。它是种群重要的结构特征之一,也是种群的繁殖特性对不同环境因子長期适应的结果,在一定程度上可以反映群落结构现状和更新策略,更是预测种群或群落动态发展趋势的有效途径(Gao et al., 2003; Miao et al., 2018)。本研究发现,青冈种群径级结构为倒“J”型,说明小径级的青冈幼苗数量储备较为丰富,其种群属于增长型,具有较强的自我更新和维持能力,若不受强烈的干扰,将来该种群数量会保持稳定,这与汪国海等(2021)的研究结果相同。青冈种群小径级个体所占的比例最高。一方面,由其繁殖特性决定,青冈成树产生的成熟种子在重力作用下大多都聚集于母株冠幅下方,而自然条件下萌发形成大量小径级个体也集中分布在母株冠幅周围,在母树的庇护下,占据着有利的资源, 有利于青冈幼苗个体与其他种群竞争过程中发挥群体效应。另一方面,由于岩溶石山土壤水分缺乏、土壤浅薄、岩石裸露、碳酸盐溶蚀和土壤贫瘠等特点(王明章,2004;胡芳等,2018),严重制约了青冈种子的萌发和幼苗的更新。有研究表明,幼苗是森林更新过程中最重要和最敏感的阶段,随着幼苗个体的生长,同种个体对光照、养分、水分及空间等资源的利用方式相同,但由于岩溶石山生境资源有限,种内种间竞争加剧,因此出现强烈的自疏效应(Das et al., 2011; Bai et al., 2012)。由于幼苗在生长过程中竞争能力较弱的幼苗个体会大量死亡,只有少量个体能够成功向幼树阶段转化,所以样地内存在大量的幼苗,而幼树和成树的数量却较少(Hu et al., 2012)。这样的种群径级结构分布能够维持种群持续健康发展,保持良好的更新潜力,这对该种群的稳定具有积极意义。
3.2 青冈种群的空间分布格局
种群的空间分布格局是种群在二维空间上布局的一个定量描述,研究其特征有助于阐明种群空间分布格局的形成机制(Comita & Hubbell, 2009)。种群的空间分布格局与尺度存在密切的关系,随着尺度的变化而变化(张华等, 2016)。本研究发现,青冈种群在小尺度上均表现为聚集分布,随尺度增加,其聚集强度逐渐减弱,在中、大尺度上以随机分布为主(幼苗除外)。这与拓锋等(2020)、张华等(2016)、任毅华等(2021)的研究结果具有一致性。一方面,小尺度上的物种分布格局主要受到种群自身的生物学特性影响(Zhao et al., 2010; Lin et al., 2011),如青冈成树成熟种子呈卵形、长卵形或椭圆形且无毛或被薄毛,主要靠重力传播,大部分成熟种子因重力作用而直接沉积在母株冠幅下方,而高度裸露的岩石形成地漏、石缝、石槽等地理屏障,从而限制了青冈成熟种子的传播距离。另一方面,岩溶石山微地形复杂、土壤总量少且斑块化,使得水分、养分资源等生态因子的再分配呈很高的空间异质性,在空间上形成更多的微生境隔离(Wiegand et al., 2004; Hu et al., 2012),这样青冈成树的成熟种子能到达适宜萌发的微生境的概率大大降低,从而导致幼苗呈聚集分布。虽然幼苗的聚集分布会加剧种内不同个体之间对光照、养分、水分和空间等资源的竞争,但这种聚集分布有利于种群发挥群体效应,能加快区域贫瘠环境的营养积累和幼苗因竞争而发生死亡后营养元素的循环,使得该环境更有利于更新幼苗的生长,从而提高植物的种间竞争和抵抗外界不良环境的能力(Ripley, 1981; Harms et al., 2001)。
随着植株个体年龄增长,植株对养分等资源的需求逐渐增大,个体之间的竞争也会逐渐增加,种内和种间的个体之间因竞争激烈而出现大量的个体死亡现象(即自疏和他疏现象),最终导致种群密度的大大降低,其聚集强度也逐渐减弱(Nathan & Muller-Landau, 2000; Comita & Hubbell, 2009)。因此,随着年龄增长,种群由聚集分布向随机分布或均匀分布转化。另外,中、大径级青冈种群之所以在较大尺度上呈随机分布,可能是因为青冈种群为优势树种且占据林冠层,而青冈成树的树冠和根系延伸到一定范围内基本定型,因种内和种间竞争加剧,自疏和他疏效应使得该种群由聚集分布向随机分布转化(Wiegand et al., 2004;汪国海等,2021)。
3.3 青冈种群的空间关联性
植物种群不同径级个体的空间关联可以揭示种群内部个体间在某一特定时间内的空间分布和功能关系,这是种群个体与周围环境长期相互作用的结果,对同一种群不同径级个体的空间关联研究有利于描述种群的现状并预测种群的动态状况(Schleicher et al., 2011; 王慧杰等,2016)。本研究结果发现,幼苗与幼树在0~15 m尺度上呈显著正相关,说明幼苗与幼树之间存在正向或相互有利的生态关系。由于幼苗、幼树对光照、水分等资源的竞争能力都比较弱,但为了生存,他们不得不采取协同策略来适应不利的生境,因此体现出一种互相帮助或互相“庇护”的互利共生关系(Comita & Hubbell, 2009)。Das等(2011)研究表明,小径级个体之间的空间关联性呈正关联,表明他们对环境资源的选择具有一致性,它们在空间分布上通常结伴出现或相互依存。另外,小径级个体之间的生长发育阶段较为接近,对环境资源的需求具有相似性且存在生态位重叠的现象,而幼树相对幼苗来说,对光照、水分、养分等资源的竞争相對较强,仍然需要借助高大的成树进行庇护(Harms et al., 2001; 宋于洋等, 2010)。因此,幼苗与幼树个体需要采取互惠的生态关系来适应资源有限的岩溶石山生境,从而提高幼苗与幼树的成活率。幼树与成树在0~10 m尺度上无显著关联性,在10~15 m尺度上呈正相关性,这主要是成树占据林分的主林层,幼树与成树占据不同的生态位空间,进而对光照等资源的竞争不激烈,能够彼此相对独立的分享生境资源,表现出种群处于一种较稳定发展状态(李明辉等, 2011;Lan et al., 2012)。幼苗与成树在0~2 m尺度上表现出无显著关联性,在2~15 m尺度上表现出正相关,这是由种子扩散限制或种间竞争引起的(Wiegand & Moloney, 2004)。幼苗与成树的关联性变化与青冈种子自身的生物学特性有关,青冈成树能够产生大量种子,主要通过重力散播,而幼苗的分布范围在一定程度上体现了成熟种子的分布情况。
4 结论
本研究结果表明径级结构呈倒“J”型分布,说明该种群处于稳定发展阶段且具有一定自我维持和更新的能力。幼苗、幼树和成树在较小尺度上表现为聚集分布,但幼树和成树在中、大尺度上基本表现为随机分布。这表明岩溶石山生境贫瘠,聚集分布有利于种群在竞争有限资源时发挥群体效应,从而提高种群的存活率。而幼苗与幼树和成树之间主要呈空间正相关,说明幼苗对环境资源的需求与幼树和成树具有相似性和互补性,但由于幼苗竞争能力弱,因此需要借助大树进行庇护才能生存。本研究结果揭示了岩溶石山脆弱生态系统青冈种群不同发育阶段的空间分布规律及其空间关联情况,为我国西南岩溶石山地区植被重建与恢复以及生态系统服务功能的评价提供了科学依据。
参考文献:
BAI XJ, QUEENBOROUGH SA, WANG XG, et al., 2012. Effects of local biotic neighbors and habitat heterogeneity on tree and shrub seedling survival in an old-growth temperate forest [J]. Oecologia, 170: 755-765.
CALL LJ, NILSEN ET, 2003. Analysis of spatial patterns and spatial association between the invasive tree-of-heaven (Ailanthus altissima) and the native black locust (Robinia pseudoacacia) [J]. Am Midl Nat, 150(1): 1-14.
CAO GX, ZHONG ZC, LIU Y,et al., 2003. The study of distribution pattern of Camellia rosthorniana population in Jinyun Mountain [J]. J Biol, 20(1): 10-12.
COMITA LS, HUBBELL SP, 2009. Local neighborhood and speciesshade tolerance influence survival in a diverse seedling bank [J]. Ecology, 90(2): 328-334.
DAS A, BATTLES J, STEPHENSON NL, 2011. The contribution of competition to tree mortality in old-growth coniferous forests [J]. For Ecol Manag, 261: 1203-1213.
DIGGLE PJ, 1983. Statistical analysis of spatial point patterns [M]. New York: Academic Press.
GAO FY, ZHAO CZ, 2012. In the process of grassland degradation the spatial pattern and spatial association of dominant species [J]. Acta Ecol Sin, 32(21): 6661-6669. [高福元, 赵成章, 2012. 甘肃臭草型退化草地优势种群空间格局及其关联性 [J]. 生态学报, 32(21): 6661-6669.]
GUO YL, WANG B, XIANG WS,et al., 2015. Spatial distribution of tree species in a tropical karst seasonal rainforest in Nonggang, Guangxi, southern China [J]. Biodivers Sci, 23(2): 183-191. [郭屹立, 王斌, 向悟生, 等, 2015. 广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析 [J]. 生物多样性, 23(2): 183-191.]
HARMS KE, CONDIT R, FOSTER HRB, 2001. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot [J]. J Ecol, 89(6): 947-959.
HE ZL, XU H, QIN XS, et al., 2017. Spatial distribution patterns and association of two Apocynaceae plants in the tropical mountain rainforests of Jianfengling, Hainan Island, China [J]. Biodivers Sci, 25(10): 1065-1074. [何增麗, 许涵, 秦新生, 等, 2017. 海南尖峰岭热带山地雨林2种夹竹桃科植物的空间分布格局与关联性 [J]. 生物多样性, 25(10): 1065-1074.]
HU F, ZENG FP, DU H, et al., 2018. Scale-dependent spatial patterns for species diversity in a karst evergreen and deciduous broad-leaved mixed forest of northwest Guangxi [J]. Acta Ecol Sin, 38(17): 6074-6083. [胡芳, 曾馥平, 杜虎, 等, 2018. 桂西北喀斯特常绿落叶阔叶混交林物种多样性分布格局的尺度效应 [J]. 生态学报, 38(17): 6074-6083.]
HU YH, SHA LQ, BLANCHET FG, et al., 2012. Dominant species and dispersal limitation regulate tree species distributions in a 20-ha plot in Xishuangbanna, southwest China [J]. Oikos, 121: 952-960.
HU G, LIANG SC, ZHANG ZH, 2007. Quantitative analysis of Cyclobalanopsis glauca community on karst hills of Guilin [J]. Chin J Ecol, 26(8): 1177-1171.
HU G, ZENG SQ, LONG SS, 2017. Structure and dynamic characteristics of Cyclobalanopsis glauca secondary forests [J]. J Cent S Univ For Technol, 37(11): 110-114.
HUBBELL SP, 2001. The unified neutral theory of biodiversity and biogeography [M]. Princeton: Princeton University Press: 127-139.
LIN YC, CHANG LW, YANG KC, et al., 2011. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation [J]. Oecologia, 165(1): 175-184.
LAN G, GETZIN S, WIEGAND T,et al., 2012. Spatial distribution and interspecific associations of tree species in a tropical seasonal rain forest of China [J]. PLoS ONE, 7(9): e46074.
LI XK, HE CX, TANG JS, 2008. Evolution and ecological processes of karst ecosystem of Guangxi [J]. Guangxi Sci, 15(1): 80-86.
LI MH, HE FH, PAN CD, 2011. Spatial distribution pattern of different strata and spatial assocations of different strata in the Schrenk Spruce Forest, northwest China [J]. Acta Ecol Sin, 31(3): 620-628. [李明辉, 何风华, 潘存德, 2011. 天山云杉天然林不同林层的空间格局和空间关联性 [J]. 生态学报, 31(3): 620-628.]
LIU YS, DENG XS, HU YC, 2006. Rocky land degradation and poverty alleviation strategy in Guangxi karst mountainous area [J]. Mt Res, 24(4): 228-233. [刘彦随, 邓旭升, 胡业翠, 2006. 广西喀斯特山区土地石漠化与扶贫开发探析 [J]. 山地学报, 24(4): 228-233.]
MIAO N, XU H, MOERMOND TC, et al., 2018. Density-dependent and distance-dependent effects in a 60-ha tropical mountain rain forest in the Jianfengling mountains, Hainan Island, China: Spatial pattern analysis [J]. For Ecol Manag, 429: 226-232.
NATHAN R, MULLER-LANDAU HC, 2000. Spatial patterns of seed dispersal, their determinants and consequences for recruitment [J]. Trends Ecol Evol, 15(7): 278-285.
RIPLEY BD, 1981. Spatial statistics [M]. New York: John Wiley & Sons: 252.
RIPLEY BD, 1977. Modelling spatial pattern [J]. J Royal Statist Soc, 39(2): 172-192.
REN YH, ZHOU YZ, HOU L, et al., 2021. Spatial distribution patterns of standing trees at different ages in Abies georgei var. smithii forests in Sejila Mountain [J]. Acta Ecol Sin, 41(13): 5417-5424. [任毅华, 周尧治, 侯磊, 等, 2021. 色季拉山急尖长苞冷杉种群不同龄级立木的空间分布格局 [J]. 生态学报, 41(13): 5417-5424.]
STOYAN D, PENTTINEN A, 2000. Recent applications of point process methods in forestry statistics [J]. Statist Sci, 15(1): 61-78.
SCHLEICHER J, WIEGAND K, WARD D, 2011. Changes of woody plant interaction and spatial distribution between rocky and sandy soil areas in a semi-arid savanna, South Africa [J]. J Arid Environ, 75(3): 270-278.
SU SJ, LIU JF, CHEN WW, 2014. Spatial variability and patterns of soil moisture physical properties in Pinus taiwanensis forest based on geostatistics and GIS [J]. Resour Sci, 36(11): 2423-2430. [苏松锦, 刘金福, 陈文伟, 等, 2014. 戴云山黃山松林土壤水分物理性质空间变异特征与格局 [J]. 资源科学, 36(11): 2423-2430.]
SONG YY,LI YY, ZHANG WH, 2010. Analysis of spatial pattern and spatial association of Haloxylon ammodendron population in different developmental stages [J]. Acta Ecol Sin, 30(16): 4317-4327. [宋于洋, 李园园, 张文辉, 2010. 梭梭种群不同发育阶段的空间格局与关联性分析 [J]. 生态学报, 30(16): 4317-4327.]
TUO F, LIU XD, LIU RH, et al., 2020. Spatial distribution patterns and association of Picea crassifolia population in Dayekou Basin of Qilian Mountains, northwestern China [J]. Chin J Plant Ecol, 44(11): 1172-1183. [拓锋, 刘贤德, 刘润红, 等, 2020. 祁连山大野口流域青海云杉种群空间格局及其关联性 [J]. 植物生态学报, 44(11): 1172-1183.]
WIEGAND T, MOLONEY KA, 2004. Rings, circles, and null-models for point analysis in ecology [J]. Oikos, 104(2): 209-229.
WANG GH, PAN Y, QIN GL, et al., 2021. Population structure and spatial distribution pattern of Kmeria septentrionalis an endangered species, in karst habitat [J]. For Res, 34(3): 81-87. [汪国海, 潘扬, 覃国乐, 等, 2021. 喀斯特生境中濒危植物单性木兰种群结构及空间分布格局研究 [J]. 林业科学研究, 34(3): 81-87.]
WANG HJ, CHANG SL, ZHANG YT, et al., 2016. Density-dependent effects in Picea schrenkiana forests in Tianshan Mountains [J]. Biodivers Sci, 24(3): 252-261. [王慧杰, 常顺利, 张毓涛, 等, 2016. 天山雪岭云杉森林群落的密度制约效应 [J]. 生物多样性, 24(3): 252-261.]
WANG MZ, 2004. A deliberation upon the karst environment of geology and its implication [J]. Guizhou Geol, 21(4): 251-253. [王明章, 2004. 论喀斯特生态地质环境及其研究意义 [J]. 贵州地质, 21(4): 251-253.]
WANG XT, WANG DJ, LI HB, et al., 2022. Cumulative effects of K-function in the research of point patterns [J]. Chin J Appl Ecol, 33(5): 1275-1282. [王鑫厅, 王殿杰, 李海兵, 等, 2022. 点格局研究过程中K-函数的累积效应 [J]. 应用生态学报, 33(5): 1275-1282.]
WANG XT, HOU YL, LIANG CZ, et al., 2012. Point pattern analysis based on different null models for detecting spatial patterns [J]. Biodivers Sci, 20(2): 151-158. [王鑫厅, 侯亚丽, 梁存柱, 等, 2012. 基于不同零模型的点格局分析 [J]. 生物多样性, 20(2): 151-158.]
XU YF, WANG KL, QI XK, et al., 2016. Spatial and temporal vegetation changes under geological settings of dolomite and limestone based on TM images [J]. Acta Ecol Sin, 36( 1):180-189. [徐艳芳, 王克林, 祁向坤, 等, 2016. 基于TM影像的白云岩与石灰岩上喀斯特植被时空变化差异研究 [J]. 生态学报, 36(1): 180-189.]
ZHAO CZ, GAO FY, WANG XP,et al., 2010. Fine-scale spatial patterns of Stellera chamaejasme population in degraded alpine grassland in upper reaches of Heihe, China [J]. Chin J Plant Ecol, 34(11): 1319-1326.
ZHANG ZH, HU G, ZHU JD, et al., 2013. Aggregated spatial distributions of species in a subtropical karst forest, southwestern China [J]. J Plant Ecol, 6(2): 131-140.
ZHANG JT, 2004. Two methods for two dimensional pattern analysis of community [J]. Acta Bot Boreal-Occident Sin, 24(8): 1448-1451. [张金屯, 2004. 群落二维格局分析的两种方法 [J]. 西北植物学报, 24(8): 1448-1451.]
ZHANG H, HUANG Q, ZHAO CY, et al., 2016. Spatial pattern analysis of individuals in different age-classes of Picea crassifolia [J]. Chin J Ecol, 35(5): 1182-1187. [张华, 黄琴, 赵传燕, 等, 2016. 青海云杉不同龄级立木的点格局分析 [J]. 生态学杂志, 35(5): 1182-1187.]
(责任编辑 蒋巧媛 邓斯丽)
收稿日期: 2022-04-30
基金项目: 国家自然科学基金(31860124); 国家级大学生创新创业训练计划项目(20181060263)。
第一作者: 盘远方(1994-),硕士,研究实习员,研究方向为植物生理生态学,(E-mail)yuanfangpan124@163.com。
通信作者: 李麗香,硕士,讲师,研究方向为植物生态学,(E-mail)lixiang_li66@126.com。
