黔中天龙山典型喀斯特次生林地上生物量与环境因子的关系
2023-05-30叶天木容丽王梦洁李璇杨文松王琪
叶天木 容丽 王梦洁 李璇 杨文松 王琪


摘 要: 为探究喀斯特次生林地上生物量与环境因子的关系,该文以黔中普定縣喀斯特天龙山典型次生林样地为研究对象,采取单物种及不同径级组地上生物量模型计算优势种及群落生物量,采用空间分布图描述环境因子与群落地上生物量空间分布状况,利用相关性检验(Pearson)、一般线性模型(GLM)以及冗余分析(RDA)讨论群落、生活型、物种地上生物量与环境因子的关系。结果表明:(1)喀斯特次生林群落地上生物量总量为106.94 t·hm-2,优势种地上生物量占整个样地的91.77%,其中常绿植物高于落叶,窄叶柯(Lithocarpus confinis)与化香树(Platycarya strobilacea)地上生物量在群落中占比最高,分别为34.23%和34.37%。(2)岩石裸露率空间分布呈现明显的上下梯度差异,上坡显著大于下坡,坡度与土壤厚度空间分布不连续,无明显规律。(3)群落地上生物量与土壤厚度呈显著正相关,二者空间分布趋于一致,土壤厚度是群落地上生物量的主要影响因子,岩石裸露率与坡度对群落地上生物量的影响较低。(4)对于不同生活型,岩石裸露率对地上生物量的影响程度最高,土壤厚度与坡度对常绿植物的影响大于落叶。(5)对于不同物种,环境因子与地上生物量的相关性复杂,大部分物种与土壤厚度呈正相关,化香树、刺异叶花椒(Zanthoxylum dimorphophyllum var. spinifolium)分别与岩石裸露率、坡度呈正相关。综上认为,该研究区次生林植被恢复缓慢,土壤厚度是群落地上生物量的主要影响因子,但对于不同生活型和物种而言,地上生物量对环境的响应会受到生活型差异、物种生境偏好及种间关系等生物因素的影响。
关键词: 喀斯特, 地上生物量, 土壤厚度, 岩石裸露率, 坡度
中图分类号: Q948
文献标识码: A
文章编号: 1000-3142(2023)03-0484-10
Relationship between aboveground biomass and environmental
factors of typical karst secondary forest in
Tianlong Mountain, central Guizhou
YE Tianmu1,2, RONG Li1,2*, WANG Mengjie1,2, LI Xuan1,2, YANG Wensong1,2, WANG Qi1,2
( 1. College of Geography and Environmental Sciences, Guizhou Normal University (Karst Research Institute), Guiyang 550001, China;
2. Puding Karst Ecological Observation and Research Station, Chinese Academy of Sciences, Anshun 562100, Guizhou, China )
Abstract: In order to explore the relationship between the aboveground biomass of karst secondary forest land and environmental factors, we took the typical secondary forest sample of karst Tianlong Mountain in Puding County, Central Guizhou as the research object, adopted the aboveground biomass model of single species and different diameter groups to calculate the biomass of dominant species and community, used the spatial distribution map to describe the spatial distribution of environmental factors and aboveground biomass of community, and used the correlation test (Pearson) General linear model (GLM) and redundancy analysis (RDA) to discuss the relationship between community, life form, species aboveground biomass and environmental factors. The results were as follows: (1) The total aboveground biomass of karst secondary forest group was 106.94 t·hm-2, and the aboveground biomass of dominant species accounted for 91.77% of the whole sample plot. The aboveground biomass of evergreen plants was higher than that of deciduous plants. The aboveground biomass of Lithocarpus confinis and Platycarya strobilacea accounted for the highest proportion in the community, 34.23% and 34.37% respectively. (2) The spatial distribution of rock exposure rate showed obvious upper and lower gradient difference, the upper slope was significantly greater than the lower slope, and the spatial distribution of slope and soil thickness was discontinuous and had no obvious law. (3) There was a significant positive correlation between aboveground biomass and soil thickness, and their spatial distribution tended to be consistent. Soil thickness was the main influencing factor of aboveground biomass of the community, and the influence of rock exposure rate and slope on aboveground biomass of the community was low. (4) For different life forms, the influence of rock exposure rate on aboveground biomass was the highest, and the influence of soil thickness and slope on evergreen plants was greater than that of defoliation. (5) For different species, the correlation between environmental factors and aboveground biomass was complex. Most species were positively correlated with soil thickness. Platycarya strobilacea and Zanthoxylum dimorphophyllum var. spinifolium were positively correlated with rock exposure rate and slope respectively. In conclusion, the vegetation restoration of secondary forest in the study area is slow, and soil thickness is the main influencing factor of aboveground biomass of the community. However, for different life forms and species, the response of aboveground biomass to the environment will be affected by biological factors such as life form differences, species habitat preferences and interspecific relationships.
Key words: karst, aboveground biomass, soil thickness, rock exposure rate, slope
生物量是指生物個体或者群落在一段时间积累的有机质总量(付威波等,2014),是生态系统获取能量的重要指标,植物生物量作为生态系统中生产力的基础,在植被恢复的评价过程中有着重要意义(Nagaraja et al., 2005;郭娜和刘剑秋,2011)。森林是陆地生态系统的主体,森林生物量不仅能揭示森林生态系统能量平衡、养分循环和生产力等功能过程的变化规律(Poorter et al., 2012),还可以反映出生态系统功能的强弱(赵士洞和汪业勖,2001)。目前,关于森林生物量已有大量研究,基于收获法、分层切割法、皆伐法及遥感等方法对不同区域进行估算(范文义等,2011;李尚益等,2018;刘立斌等,2020), 对个体、种群、群落、区域到生物圈等多个尺度进行分析(巨文珍和农胜奇,2011)。此外,植物生物量不仅与气候、地形、土壤养分等非生物因素存在显著关系(Krankina et al., 2005;杨远盛等,2015;邓瑞明,2018),还受到林分类型、物种特性、种间竞争等生物因素的影响(Chen et al., 2004;杨远盛等,2015;李尚益等,2018)。但是,由于森林类型以及区域差异,影响植物生物量的因素并不统一(Ashton & Hall, 1992),因此,通过分析不同区域森林群落生物量与环境关系,有利于补充生态功能研究的完整性。同时,关于两者关系多为大尺度讨论(王晓莉等,2014;沙威等,2016;刘莉等,2017),Burke(2001)研究表明,小尺度上的植物群落发育更多由非地带性因素主导,因此,从小尺度上探讨植物生物量与环境关系对理解植被功能维持机制具有重要意义。
位于贵州中部的普定县(简称黔中)是喀斯特石漠化的典型地区,土层浅薄且不连续,土壤持水能力差,石缝、石沟等小生境复杂多样(刘长成等,2009),在此立地条件上生长的植物多具有耐旱性、石生性等特征,明显不同于非喀斯特地区植被。目前,对喀斯特森林生物量的研究主要集中于森林地上生物量的估算(刘之洲等,2017;刘立斌等,2020),而关于地上生物量与环境关系的研究相对较少且侧重于单个因素的影响,如土壤厚度、地形、火烧等(张喜等,2011;马士彬,2019;刘媛,2020),喀斯特地区立地环境复杂,仅考虑单一因素不足以揭示喀斯特森林生态功能系统的维持机制。
本文以普定天龙山喀斯特典型次生林为研究对象,选取坡度、岩石裸露率、土壤厚度作为环境因子,从群落、生活型、物种3个角度分析植物地上生物量与环境因子的关系,拟探讨以下问题:(1)3种环境因子对群落地上生物量的影响程度;(2)对于不同生活型及不同物种,植物地上生物量对环境因子的响应程度有何变化。
1 研究区与研究方法
1.1 研究区和样地概况
样地位于贵州省中部的普定县,贵州高原长江水系与珠江水系的分水岭处,海拔为1 460 m。属于中亚热带季风温暖湿润气候,季风交替十分明显,全年气候温和,辐射能量低,年均气温为15.1 ℃,平均日照时数为1 164.9 h,无霜期长,可达到301 d,雨量充沛,年均降水为1 378.2 mm,降水主要集中在5—9月。该研究区是典型的喀斯特次生林,植被以壳斗科(Fagaceae)、樟科(Lauraceae)、胡桃科(Juglandaceae)、海桐科(Pittosporaceae)、虎耳草科(Saxifragaceae)、鼠李科(Rhamnaceae)、桦木科(Betulaceae)、芸香科(Rutaceae)、榆科(Ulmaceae)为主。
1.2 样地调查和样品采集
2012年在贵州省普定县(105°45′ 06.65″ E、26°22′ 07.06″ N)天龙山南坡设置面积为2 hm2的永久监测样地,样地划分为200个10 m×10 m的小样方。本研究于2020年对样方内DBH≥1 cm的木本植物进行复查,调查群落中的植物种类、数量、高度、胸径、冠幅等,同时记录样方的坡度、岩石裸露率,并在每个10 m×10 m小样方内的对角线上使用带刻度的钢钎随机测5次土壤厚度,取其平均值(刘媛,2020)。
1.3 计算和统计方法
1.3.1 重要值计算 重要值表征物种的优势程度(李博,2020),其方法如下:
重要值=(相对密度+相对频度+相对显著度)/3;相对频度=(某物种的频度/全部物种的频度总和)×100%;相对显著度=(某物种个体胸面积和/全部物种胸面积总和)×100%;相对密度=(某物种植物的密度/全部物种的总密度)×100%。
1.3.2 地上生物量估算 本研究中地上生物量计算公式使用刘长成及本课题组在相同样地根据植物的干重、胸径、高度、冠幅的回归关系所拟合的物种以及径级组地上生物量模型(刘长成等,2009),优势种地上生物量采用8个单独模型(表1),其余物种则采用不同径级的回归模型,最后求和得出群落地上生物量。
1.3.3 统计方法 根据地上生物量估算模型计算每个小样方的地上生物量,利用Origin 2021软件制作群落地上生物量、土壤深度、岩石裸露率、坡度的空间分布图,在R语言中使用Corrplot制作土壤厚度、岩石裸露率、坡度、地上生物量的相关性热图。通过IBM SPSS Statistics软件中的一般线性模型,分析环境因子对群落和不同生活型地上生物量的解释度,采用Canoc4.5软件中的冗余分析(RDA)分析优势种地上生物量与环境因子之间的关系,实心箭头表示优势种地上生物量,空心箭头表示环境因子,相关性等于矢量之间角度的余弦,两个矢量之间的夹角小于90°说明两者之间呈正相关;大于90°且小于180°说明两者之间呈负相关;两者之间的夹角越接近90°,则相关性越弱(龙建等,2012)。
2 结果与分析
2.1 天龙山次生林地上生物量特征
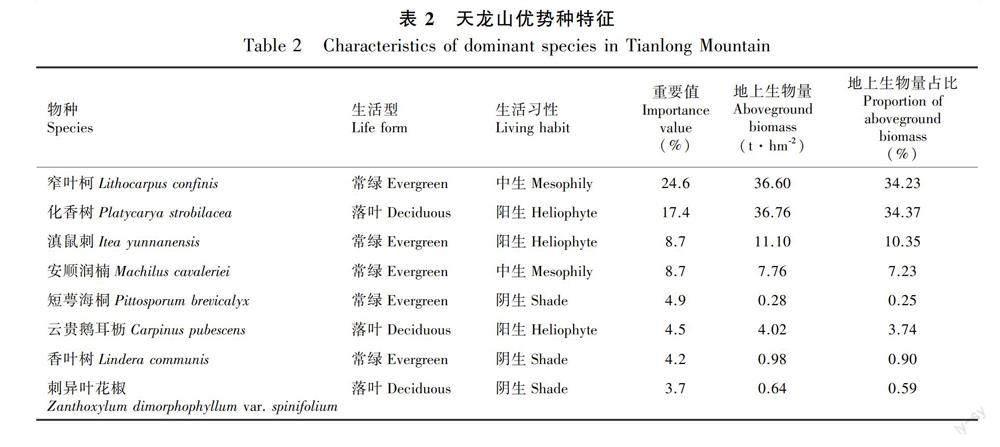
喀斯特天龙山典型次生林主要物种有窄叶柯(Lithocarpus confinis)、化香树(Platycarya strobi-lacea)、滇鼠刺(Itea yunnanensis)、安顺润楠(Machilus cavaleriei)、短萼海桐(Pittosporum brevicalyx)、云贵鹅耳枥(Carpinus pubescens)、香叶树(Lindera communis)、刺异叶花椒(Zanthoxylum dimorphophyllum var. spinifolium)、朴树(Celtis sinensis)、倒卵叶旌节花(Stachyurus obovatus)、珊瑚冬青(Ilex corallina)、多脉猫乳(Rhamnella martini)、异叶鼠李(Rhamnus heterophylla)、白蜡树(Fraxinus chinensis)、猴樟(Cinnamomum bodinieri)等。根据植物群落实际情况,将重要值排前8的物种作为优势种。
天龙山次生林群落地上生物量为106.94 t·hm-2,优势种地上生物量占群落的91.77%(表2),其中落叶植物占41.42 t·hm-2,常绿植物占56.72 t·hm-2。窄叶柯单个物种占群落地上生物量的34.23%,重要值为24.6%,是天龙山次生林中的建群种;化香树地上生物量占比最高,为34.37%。
2.2 地上生物量与环境因子空间分布特征
天龙山样地土壤厚度均值为15.59 cm,最高值为35.40 cm,最低值为5.80 cm,变异系数为68.03%(表3),根据岩溶地区的坡度地划分,该样地为陡坡(李瑞玲,2004),其变异系数较低(34.88%)。岩石裸露率平均值为44.69%,且空间分布呈明显的上下坡梯度差异(图1),岩石裸露率较高的区域基本分布于样地上坡,坡度整体呈不连续性,而群落地上生物量空间分布与土壤厚度趋于一致。
2.3 地上生物量与环境因子之间的关系
从图2可以看出,通過相关性分析发现,群落地上生物量与土壤厚度呈显著正相关(P<0.01),岩石裸露率与坡度呈显著正相关(P<0.01),土壤厚度与坡度(P<0.01)、岩石裸露率(P<0.05)呈负相关。
从表4可以看出,土壤厚度对群落地上生物量的解释度最高(R2=0.505),而岩石裸露率和坡度则较低。对于不同生活型,岩石裸露率对常绿与落叶植物的解释度最高(R2=0.386,R2=412),坡度与土壤厚度对常绿植物的影响大于落叶植物,而岩石裸露率对常绿植物的影响略小于落叶植物。
从图3可以看出,3个环境因子与8个优势种呈现不同的相关性,窄叶柯、刺异叶花椒、云贵鹅耳枥、短萼海桐与土壤厚度呈正相关。化香树与岩石裸露率呈正相关,刺异叶花椒与坡度呈正相关。
3 讨论
3.1 优势种对群落地上生物量起决定作用
天龙山次生林地上生物量总量为106.94 t·hm-2,显著低于多年前同一气候带下的云南哀牢山木果石栎林(348.7 t·hm-2)(邱学钟,1984)以及茂兰原生林(168.02 t·hm-2)(朱守谦等,1995),喀斯特天龙山次生林在五六十年代受过间伐,虽然经过较长时间恢复,但其地上生物量仍然较低,说明该特殊环境背景下,植被的恢复速度较慢。
喀斯特土壤贫瘠,立地条件差,环境过滤出的优势种能较好适应喀斯特的环境胁迫,在竞争中能保持优势,在同样条件下能大量生长并积累营养元素(陈萍,2015;王磊等,2020),优势种占群落地上生物量的91.77%, 对群落地上生物量起决定
性作用,与选择效应假说一致(Slik et al., 2010)。本研究中的常绿植物地上生物量大于落叶植物,可能由于常绿植物更适合南方湿润喀斯特环境,黔中地区日照率较低,仅为26.3%(倪健等,2017),使得偏好光照资源的落叶植物在竞争过程中呈劣势(Givnish, 2001;车俭等,2020)。
窄叶柯与化香树的地上生物量分别占群落地上生物量的34.23%和34.37%,属于喀斯特次生林群落中的主要乔木树种,耐旱、耐贫瘠能力强,物种个体数量多且分布广泛,生态幅宽(李婷婷,2021)。窄叶柯在天龙山扮演着建群种和主要生产者的角色,且窄叶柯是中生性植物,在未来的群落演替中仍有较大的成长空间。化香树的重要值虽低于窄叶柯,但其地上生物量却略高于窄叶柯,可能因为本文中地上生物量计算模型是根据植物的干重与胸径、高度与冠幅的多种回归关系所拟合(刘长成等,2009),而化香树的植株干重相对较高,即在个体大小相同的情况下,化香树地上生物量高于窄叶柯。
3.2 群落地上生物量与土壤厚度空间分布趋于一致
从空间分布情况来看,岩石裸露率有着明显的上下梯度差异,可能是上坡受到雨水冲刷较多,使得样地上方基岩大量裸露。坡度与土壤厚度的空间分布不连续,土壤厚度与微地形关系密切,体现了喀斯特生境高度异质性。Sheng等(2018)研究表明,环境因子会影响植物的空间分布状况。土壤厚度是影响群落物种多样性、植物生物量的重要因子(刘媛,2020),本文中群落地上生物量呈不连续分布,地上生物量较高的区域土壤较厚,二者空间分布趋于一致,反映出植物地上生物量对土壤厚度的高度依赖性,与大部分学者的相关研究结果相符(王志强,2007;李程程等,2012),这可能因为更深的土壤厚度具有较高的土壤质量,土壤微生物的数量更丰富(周玮等,2018)有关,同时,较厚的土壤能为植物根系提供更多的生存空间,并且能提升土壤对水分与矿物养分的保持能力(Belcher et al., 1995;李程程等,2012),从而影响植物的生长发育,使得其拥有较高的生物量。
3.3 不同生态类群、物种对环境响应方式不同
通过一般线性模型与Pearson相关性检验分析,结果显示土壤厚度对群落地上生物量的解释度最高,与前人的研究结果一致(梁千慧,2016;刘媛,2020),说明土壤厚度是群落地上生物量的主要驱动力。坡度和岩石裸露率对群落地上生物量的解释度较低,这主要可能与喀斯特植物的生态适应性有关,具有攀缘特性的物种(如藤黄檀等)在坡度较高的环境中仍能生长(魏兴琥等,2014)。此外,植物为适应喀斯特环境形成了独特的养分吸收策略(Huang et al., 2011),如化香树,其根系发达,可在岩石缝隙中扎根生长,从中吸收养分和水分(徐良等,1997)。岩石裸露率、坡度与土壤厚度呈负相关是一种普遍的现象,土壤的抗流失能力一般随坡度的增高而降低,而基岩裸露也会造成土壤黏性降低,引起土壤流失(Heimsath et al., 1997;张伟等,2007)。因此,坡度和岩石裸露率可能是通过影响土壤厚度来间接作用于群落地上生物量。
同一生活型是指植物对环境有相似的要求与适应能力(高贤明和陈灵芝,1998;杨秀清等,2017)。在本文中,岩石裸露率对地上生物量的解释度最高,说明以岩石裸露率为主导的生境分化是天龙山次生林常绿与落叶植物地上生物量空间分布差异以及共存的重要因素之一,而土壤厚度对常绿植物的解释度大于落叶植物, 这可能是由不同生活型植物的生态习性差异所致,常绿植物更倾向于对土壤肥力和水分的选择,而落叶植物通过规律性落叶来降低自身消耗,使其能在土壤资源稀缺的环境中生存,并且落叶植物对光照有更高需求(车俭等,2020),岩石裸露较高的区域,林木相对稀少,林窗以及空旷地较大,落叶植物为获取更多光照资源而生长在岩石裸露率高的区域。谢玉彬等(2012)发现地形因子对常绿植物的影响大于对落叶植物,本文中坡度对常绿植物的解释度大于对落叶植物的,即常绿植物由于生活习性,更容易受到坡度的限制。样地中坡度虽呈不连续分布,但其变化程度不大,加剧资源异质程度的能力有限,导致坡度对不同生活型植物地上生物量的解释度较低。
对于不同物种,植物地上生物量与环境因子间相关性更为复杂,喀斯特植物有着独特的环境偏好(胡刚等,2017),不同树种占有特定的资源和空间以实现物種间的共存(Nakashizuka, 2001)。窄叶柯、云贵鹅耳枥、短萼海桐、刺异叶花椒的地上生物量与土壤厚度呈正相关,说明以上物种在选择环境时,更加倾向于土壤资源相对丰富的区域,以此吸收更多土壤养分供给自身的生长需求,而各物种对环境因子的相关程度不同,可能是由植物的生活习性与资源利用能力差异所致,如刺异叶花椒的优势度相对较低,对资源的利用能力不及其他优势种,且喜阴的习性更倾向于林窗较小的环境,土壤较厚的区域植被相对茂盛,使刺异叶花椒与土壤厚度表现出较强的相关性。滇鼠刺对土壤资源的要求不高,为保证其在群落中的优势地位,其生境的选择与其他优势物种趋异。香叶树个体较小,根系相对较浅,岩石裸露率与坡度较高的环境会限制根系水平生长,且香叶树对水分的利用能力较强(邓晓琪等,2015),使其对土壤厚度的需求降低,其地上生物量与土壤厚度呈不相关。化香树的地上生物量与岩石裸露率呈正相关,可能是由于岩石裸露率较高的区域水土亏缺,植被相对稀疏,林冠空隙较大,而化香树有着较强的岩溶环境适应能力(徐良等,1997),为获得更多光照资源,倾向于岩石裸露率较高的区域大量生长。杨文松等(2022)发现化香树与窄叶柯、安顺润楠呈互斥的生态关系,本研究中岩石裸露率与窄叶柯、安顺润楠地上生物量均呈负相关,说明植物的环境偏好、种间关系共同影响着植物地上生物量对环境的响应方式。云贵鹅耳枥虽与化香树同为阳生性落叶树种,但与环境间的关系同化香树有明显差异,可能是由于云贵鹅耳枥作为群落演替中的先锋种,群落发育早期在土壤厚度较高的区域占据森林的上层资源,且自身竞争策略导致该物种只有在森林上层遭到破坏后,才会重新生长以补充上层(梁士楚,1992),使得在水平空间上的变化较小,表现出与化香树不同的环境响应方式,这说明植物生活史对策同样会影响地上生物量与环境的关系。
综上所述,当研究对象从群落依次变为物种时,植物地上生物量对环境因子的响应方式愈加复杂,这是由于受到不同生活型的环境适应性、物种生活史、种间关系以及生境偏好等生物因素的影响。但在本文中,尚未具体分析生物因素对植物地上生物量的作用强度,今后研究将进一步讨论生物与非生物因素对喀斯特生态功能的共同影响机制。
4 结论
本文通过探讨喀斯特典型次生林植物地上生物量与环境因子之间的关系,得出以下结论:(1)天龙山次生林群落地上生物量为106.94 t·hm-2,植被恢复缓慢,显著低于多年前同一气候带下的非喀斯特森林以及区域喀斯特原生林,土壤厚度是群落地上生物量的主要影响因子;(2)岩石裸露率的分异是造成常绿植物与落叶植物空间分布差异及共存的重要因素,常绿树种相比落叶植物更容易受到土壤厚度与坡度的限制;(3)对于不同物种而言,生物因素的干扰,如种间关系、植物生活史对策、物种生境偏好等,使得物种地上生物量与环境的关系较群落、不同生活型间更为复杂。
参考文献:
ASHTON PS, HALL P, 1992. Comparisons of structure among mixed dipterocarp forests of north-western Borneo [J]. J Ecol, 80(3): 459-481.
BELCHER JW, KEEDY PA, TWOLAN SL, 1995. Root and shoot competition intensity along a soil depth gradient [J]. J Ecol, 84(4): 673-682.
BURKE A, 2001. Classification and ordination of plant communities of the Naukluft Mountains, Namibia [J]. J Veg Sci, 12(1): 53-60.
CHE J, ZHENG J, JIANG Y, et al., 2020. Separation of phylogeny and ecological behaviors between evergreen and deciduous woody angiosperms in the subtropical forest dynamics plots of China [J]. J Plant Ecol, 44(10): 1007-1014. [车俭, 郑洁, 蒋娅, 等, 2020. 中国亚热带森林动态监测样地常绿和落叶木本被子植物谱系结构及生态习性差异 [J]. 植物生态学报, 44(10): 1007-1014.]
CHEN P, 2015. Research on nutrient distribution characteristics of secondary forest ecosystems in plateau karst of Itea yunnanensis [D]. Guiyang: Guizhou University. [陈萍, 2015. 高原型喀斯特云南鼠刺次生林生态系统营养元素分配特征研究 [D]. 貴阳: 贵州大学.]
CHEN JQ, SONG B, RUDNICKI M, et al., 2004. Spatial relationship of biomass and species distribution in an old-growth Pseudotsuga-Tsuga forest [J]. For Sci, 50(3): 364-375.
DENG XQ, RONG L, DENG XX, 2015. Study of different plant water use in karst areas [J]. Wireless Internet Technol, (1): 151-152. [邓晓琪, 容丽, 邓晓霞, 2015. 喀斯特区植物水分利用研究 [J]. 无线互联科技, (1): 151-152.]
DENG RM, 2018. Relationship between the biomass characteristics and soil nutrients among different forest types in Daiyun Mountain [J]. For Constr, (6): 29-32. [邓瑞明, 2018. 戴云山不同类型森林生物量差异特征及其与土壤养分的关系 [J]. 林业建设, (6): 29-32.]
FAN YW, LI MZ, YANG JM, 2011. Forest biomass estimation models of remote sensing in Changbai Mountain forests [J]. For Sci, 47(10): 16-20. [范文义, 李明泽, 杨金明, 2011. 长白山林区森林生物量遥感估测模型 [J]. 林业科学, 47(10): 16-20.]
FU WB, PENG WX, SONG TQ, et al., 2014. Biomass and its allocation characteristics of Eucalyptus urophylla × E. grandis plantations at different stand ages [J]. Acta Ecol Sin, 34(18): 5234-5241. [付威波, 彭晚霞, 宋同清, 等, 2014. 不同林龄尾巨桉人工林的生物量及其分配特征 [J]. 生态学报, 34(18): 5234-5241.]
GAO XM, CHEN LZ, 1998. The revision of plant life-form system and an analysis of the life-form spectrum of forst plants in the warm an analysis of the life-form temperate zone of China [J]. Chin Bull Bot, 40(6): 70-76. [高贤明, 陈灵芝, 1998. 植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析 [J]. 植物学报, 40(6): 70-76.]
GUO N, LIU JQ, 2011. A review of research on plant biomass [J]. Subtrop Plant Sci, 40(2): 83-88. [郭娜, 刘剑秋, 2011. 植物生物量研究概述(综述) [J]. 亚热带植物科学, 40(2): 83-88.]
GIVNISH TJ, 2001. Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox [J]. Silv Fenn, 36(3): 703-743.
HEIMSATH AM, DIRTRICH WE, NISHIIZUMIi K, et al., 1997. The soil production function and landscape equilibrium [J]. Nature, 388: 358-361.
HUANG YQ, LI XK, ZHANG ZF, et al., 2011. Seasonal changes in Cyclobalanopsi glauca transpiration and canopy stomatal conductance and their dependence on subtemanean water and climatic factors in rocky karst terainm [J]. J Hydrol, 402(1/2): 135-143.
HU G, ZHANG ZH, CHENG AY, et al., 2017. Characterizing and analyzing stand spatial structure of a northern subtropical karst secondary forest in tianlong mountain of central Guizhou Province, China [J]. Earth Environ, 45(1): 25-31. [胡刚, 张忠华, 程安云, 等, 2017. 黔中天龙山喀斯特次生林林分空间结构的量化与分析 [J]. 地球与环境, 45(1): 25-31.]
JU WZ, NONG SQ, 2011. Research advance of forest biomass [J]. J SW For Univ, 31(2): 78-83. [巨文珍, 农胜奇, 2011. 森林生物量研究进展 [J]. 西南林业大学学报, 31(2): 78-83.]
KRANKINA ON, HOUGHTON RA, HARMON ME, et al., 2005. Effects of climate, disturbance, and species on forest biomass across Russia [J]. Can J For Res, 35(9): 2281-2293.
LI CC, CHENG X, YANG SC, 2012. Study on the soil thickness factor of plant growth in karst mountains — Take the Guizhou Xiangbao Mountain as an example [J]. J Guizhou Norm Coll, 28(9): 38-41. [李程程, 程星, 楊士超, 2012. 岩溶山区植物生长的土壤厚度因素研究——以贵州相宝山为例 [J]. 贵州师范学院学报, 28(9): 38-41.]
LI RL, 2004. Natural background and spatial differentiation of land rocky desertification in karst areas of Guizhou [D]. Guiyang: Institute of Geochemistry, Chinese Academy of Sciences. [李瑞玲, 2004. 贵州岩溶地区土地石漠化形成的自然背景及其空间地域分异 [D]. 贵阳: 中国科学院地球化学研究所.]
LI TT, 2021. Study on the dynamics of vegetation community in the karst mountains in Central Guiyang [D]. Guizhou: Guizhou Normal University. [李婷婷, 2021. 黔中喀斯特山地次生林的群落动态研究 [D]. 贵阳: 贵州师范大学.]
LI SY, FANG X, CHEN JL, et al., 2018. Effects of different degrees of anthropogenic disturbance on biomass and spatial distribution in Subtropical forests in Central Southern China [J]. Acta Ecol Sin, 38(17): 6111-6124. [李尚益, 方晰, 陈金磊, 等, 2018. 人为干扰对中亚热带森林生物量及其空间分布格局的影响 [J]. 生态学报, 38(17): 6111-6124.]
LIANG QH, 2016. Growth and physiological response of ryegrass to karst soil depth under different water conditions [D]. Chongqing: Southwest University. [梁千慧, 2016. 不同水分条件下黑麦草对喀斯特土壤深度的生长与生理响应 [D]. 重庆: 西南大学.]
LIANG SC, 1992. A preliminary study on the structure and dynamics of pubescent hornbeam population in karst mountain of Guiyang [J]. Chin J Plant Ecol, 16(2): 108-117. [梁士楚, 1992. 贵阳喀斯特山地云贵鹅耳枥种群结构和动态初探 [J]. 植物生态学与地植物学学报, 16(2): 108-117.]
LIU LB, ZHOU YC, CHENG AY, et al., 2020. Aboveground biomass estimate of a karst forest in central Guizhou Province, southwestern China based on direct harvest method [J]. Acta Ecol Sin, 40(13): 4455-4461. [刘立斌, 周运超, 程安云, 等, 2020. 利用皆伐法估算黔中喀斯特森林地上生物量 [J]. 生态学报, 40(13): 4455-4461.]
LIU L, HAN M, LIU YB, et al., 2017. Spatial distribution of wetland vegetation biomass and its influencing factors in the Yellow River Delta Nature Reserve [J]. Acta Ecol Sin, 37(13): 4346-4355. [刘莉, 韩美, 刘玉斌, 等, 2017. 黄河三角洲自然保护区湿地植被生物量空间分布及其影响因素 [J]. 生态学报, 37(13): 4346-4355.]
LIU Y, 2020. Effects of soil depth on plant community structure and biomass in karst region [D]. Chongqing: Southwest University. [刘媛, 2020. 喀斯特土壤厚度对植物群落结构及生物量的影响 [D]. 重庆: 西南大学.]
LIU CC, WEI YF, LIU YG, et al., 2009. Biomass of canopy and shrub layers of karst forests in Puding, Guizhou, China [J]. Chin J Plant Ecol, 33(4): 698-705. [刘长成, 魏雅芬, 刘玉国, 等, 2009. 贵州普定喀斯特次生林乔灌层地上生物量 [J]. 植物生态学报, 33(4): 698-705.]
LIU ZZ, NING C, YAN WD, et al., 2017. Three kinds of pine forest biomass and carbon storage research in karst landform [J]. J Centr S Univ For Technol, 37(10): 105-111. [刘之洲, 宁晨, 闫文德, 等, 2017. 喀斯特地区3种针叶林林分生物量及碳储量研究 [J]. 中南林业科技大学学报, 37(10): 105-111.]
LONG J, LIAO HK, LI J, et al., 2012. Relationships between soil and rocky desertification in typical karst mountain area based on redundancy analysis [J]. Chin J Environ Sci, 33(6): 2131-2138. [龙健, 廖洪凯, 李娟, 等, 2012. 基于冗余分析的典型喀斯特山区土壤-石漠化关系研究 [J]. 环境科学, 33(6): 2131-2138.]
MA SB, 2019. Estimation of secondary forest age and the above-ground biomass of the tree layer in areas with complex terrain [D].Guiyang: Guizhou Normal University. [马士彬, 2019. 复杂地形区次生林林龄与乔木层地上生物量遥感估算研究 [D]. 贵阳: 贵州师范大学.]
NAKASHIZUKA T, 2001. Species coexistence in temperate, mixed deciduous forests [J]. Trends Ecol Evol, 16(4): 205-210.
NAGARAJA BC, SONASHEKAR RK, RAJ MB, 2005. Tree species diversity and composition in logged and unlogged rainforest of Kudremukh National Park, South India [J]. J Environ Biol, 26(4): 627-634.
NI J, WANG SJ, LIU LB, et al., 2017. Establishment and monitoring of biological plots at puding karst ecosystem research station [J]. Earth Environ, 45(1): 106-113. [倪健, 王世杰, 劉立斌, 等, 2017. 普定喀斯特生态系统观测研究站的生物样地建设与监测工作 [J]. 地球与环境, 45(1): 106-113.]
POORTER H, NIKLAS KJ, REICH PB, et al., 2012. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control [J]. New Phytol, 193(1) : 30-50.
QIU XZ, XIE SC, JING GF, et al., 1984. A preliminary study on the biomass of Lithocarpus xylocarpus forest in Xujiaba area, Ailao Mts, Yunnan [J]. Acta Bot Yunnan, 6(1): 85-92. [邱学忠, 谢寿昌, 荆桂芬, 等, 1984. 云南哀牢山徐家坝地区木果石栎林生物量的初步研究 [J]. 云南植物研究, 6(1): 85-92.]
SHENG MY, XIONG KN, WANG LJ, et al., 2018. Response of soil physical and chemical properties to rocky desertification succession in South China Karst [J]. Carbon Evapor, 33(1): 15-28.
SHA W, DONG SK, LIU SL, et al., 2016. Spatial patterns of plant community biomass and species diversity in Aerjin Mountain Nature Reserve and their influencing factors [J]. Chin J Ecol, 35(2): 330-337. [沙威, 董世魁, 刘世梁, 等, 2016. 阿尔金山自然保护区植物群落生物量和物种多样性的空间格局及其影响因素 [J]. 生态学杂志, 35(2): 330-337.]
SLIK J, AIBA S, BREARLEY FQ, et al., 2010. Environmental correlates of tree biomass, basal area, wood specific gravity and stem density gradients in Borneos tropical forests [J]. Glob Ecol Biogeogr, 19(1): 50-60.
WANG L, CUI M, LIU YG, et al., 2020. Relationship between stand growth and impact factors in karst area [J]. J Zhejiang A F Univ, 37(6): 1036-1044. [王磊, 崔明, 刘玉国, 等, 2020. 岩溶区林分生长与影响因子的关系 [J]. 浙江农林大学学报, 37(6): 1036-1044.]
WANG XL, CHANG Y, CHEN HW, et al., 2014. Spatial pattern of forest biomass and its influencing factors in Daxinganling, Heilongjiang Province [J]. J Appl Ecol, 25(4): 974-982. [王晓莉, 常禹, 陈宏伟, 等, 2014. 黑龙江省大兴安岭森林生物量空间格局及其影响因素 [J].应用生态学报, 25(4): 974-982.]
WANG ZQ, LIU BY, HAI CX, 2007. Effects of soil depth on vegetation cover and above ground biomass in east part of Inner Mongolia [J]. J Soil Water Conserv, 21(4): 164-167. [王志强, 刘宝元, 海春兴, 2007. 土壤厚度对天然草地植被盖度和生物量的影响 [J]. 水土保持学报, 21(4): 164-167.]
WEI XH, XU XZ, LEI LL, et al., 2014. Study on the features of vegetation in different landforms of peak forest plain in the North Guangdong [J]. Carsol Sin, 33(4): 441-448. [魏兴琥, 徐喜珍, 雷俐, 等, 2014. 粤北岩溶峰林不同地貌部位植被特征研究 [J].中国岩溶, 33(4): 441-448.]
XIE YB, MA ZP, YANG QS, et al., 2012. Coexistence mechanisms of evergreen and deciduous trees based on topographic factors in Tiantong region, Zhejiang Province, eastern China [J]. Biodivers Sci, 20(2) : 159-167. [謝玉彬, 马遵平, 杨庆松, 等, 2012. 基于地形因子的天童地区常绿树种和落叶树种共存机制研究 [J]. 生物多样性, 20(2): 159-167.]
XU L, HE XR, FANG ZJ, et al., 1997. Study on the community characteristics of the worlds only walnut single leaf incense tree [J]. J NW For Univ, 12(4): 1-6. [徐良, 何新荣, 房志坚, 等, 1997. 世界唯一胡桃目单叶化香树植物群落学特点的研究 [J]. 西北林学院学报, 12(4): 1-6.]
YANG HK, CHENG SZ, 1991. Study on biomass of the karst forest community in Maolan, Guizhou Province [J]. Acta Ecol Sin, 11 (4): 307-312. [杨汉奎, 程仕泽, 1991. 贵州茂兰喀斯特森林群落生物量研究 [J]. 生态学报, 11(4): 307-312.]
YANG XQ, SHI C, WANG XG, et al., 2017. Correlation between different life-form species and habitat in secondary Picea forest [J]. Chin J Ecol, 36(6): 1481-1487. [杨秀清, 史婵, 王旭刚, 等, 2017. 关帝山云杉次生林不同生活型物种与生境相关性 [J]. 生态学杂志, 36(6): 1481-1487.]
YANG WS, RONG L, YE TM, et al., 2022. Spacial distribution and correlation of dominant species of karst secondary forest in central Guizhou, China [J]. Chin J Appl Ecol, 33(5): 1215-1222. [杨文松, 容丽, 叶天木, 等, 2022. 黔中喀斯特次生林优势物种空间分布格局及关联性 [J]. 应用生态学报, 33(5): 1215-1222.]
YANG YS, ZHANG XX, YU HY, et al., 2015. The spatial distribution of Chinas forest biomass and its influencing factors [J]. J SW For Univ, 35(6): 45-52. [杨远盛, 张晓霞, 于海艳, 等, 2015.中国森林生物量的空间分布及其影响因素 [J]. 西南林业大学学报, 35(6): 45-52.]
YIN L, 2012. Spatial differentiation of soil thickness in a watershed of highland areas in karst region [D]. Changsha: Hunan University. [尹亮, 2012. 西南岩溶高原石漠化地区小流域土壤厚度空间分异规律研究 [D]. 长沙: 湖南大学.]
ZHAO SD, WANG YX, 2001. Forest and carbon cycle [J]. Impact Sci Soc, (3): 38-41. [赵士洞, 汪业勖, 2001. 森林与碳循环 [J]. 科学对社会的影响, (3): 38-41.]
ZHANG W, CHEN HS, WANG KL, et al., 2007. Effects of planting pattern and bare rock ratio on spatial distribution of soil nutrients in karst depression area [J]. Chin J App Ecol, 18(7): 1459-1463. [张伟, 陈洪松, 王克林, 等, 2007. 种植方式和裸岩率对喀斯特洼地土壤养分空间分异特征的影响 [J]. 应用生态学报, 18(7): 1459-1463.]
ZHANG X, CUI YC, ZHU J, et al., 2011. Influence of fire on stands of Pinus massoniana in a karst mountain area of central Guizhou Province [J]. Acta Ecol Sin, 31(21): 6442-6450. [张喜, 崔迎春, 朱军, 等, 2011. 火烧对黔中喀斯特山地马尾松林分的影响 [J]. 生态学报, 31(21):6442-6450.]
ZHOU W, YAN M, SU CH, et al., 2018. Study on soil microbial quantity and biomass developed from different carbonate-rock and soil thickness:A case study of Huaxi District in Guiyang [J]. Carsol Sin, 2018, 37(2): 168-174. [周玮, 严敏, 苏春花, 等, 2018. 不同碳酸盐岩和土层厚度下土壤微生物数量及生物量的研究——以贵阳市花溪区为例 [J]. 中国岩溶, 37(2): 168-174.]
ZHU SQ, WEI LM, CHEN ZR, et al., 1995. A preliminary study on biomass components of karst forest in Maolan of Guizhou Province, China [J]. 19(4): 358-367. [朱守谦, 魏鲁明, 陈正仁, 等, 1995. 茂兰喀斯特森林生物量构成初步研究 [J]. 植物生态学报, 19(4): 358-367.]
(责任编辑 李 莉 王登惠)
收稿日期: 2022-04-25
基金项目: 国家“十三五”重点研发计划课题(2016YFCO502607); 贵州省科技计划重大专项([2017]5411); 贵州省世界一流学科建设计划项目([2019]125)。
第一作者: 葉天木(1997-) ,硕士研究生,主要从事植物地理学研究,(E-mail) gyytm1233@163.com。
通信作者: 容丽,博士,教授,研究方向为植物生态、岩溶环境生态等,(E-mail) ronglit@126.com。
