干旱胁迫下外源ABA对甘蔗幼苗生理特性和基因表达的影响
2023-05-30农倩谢金兰林丽莫璋红王泽平宋修鹏李长宁
农倩 谢金兰 林丽 莫璋红 王泽平 宋修鹏 李长宁


关键词:甘蔗;脱落酸;干旱胁迫;生理特征;基因表达
中图分类号:S566.1 文献标识码:A
甘蔗是世界上重要的糖料作物,生长周期长、需水量大。但我国80%以上的甘蔗种植在丘陵旱坡地上,季节性干旱频繁,已成为影响甘蔗糖业可持续发展的重要因素之一。植物能通过感受刺激和信号传导,启动各种生理生化反应响应水分胁迫。干旱胁迫下,植物叶片气孔导度、叶绿素合成及电子传递会受到抑制,导致光合作用效率降低[1] 。干旱胁迫还导致植物细胞内活性氧(reactive oxygen species, ROS)大量积累,致使细胞发生膜脂过氧化作用和膜系统变性[2]。为此,植物抗氧化系统相关酶类如超氧化物歧化酶( superoxide dismutase, SOD )、过氧化物酶(peroxidase, POD)、过氧化氢酶(catalase, CAT)等的活性显著增强,非酶类抗氧剂如谷胱甘肽、类黄酮、酚类、类胡萝卜素等的含量显著提高,以清除过量ROS 积累带来的损害[3]。脯氨酸(proline,Pro)作为一种渗透调节物质,对干旱胁迫下植物细胞膜和蛋白质结构维持具有重要作用[4]。研究表明, 植物激素尤其是脱落酸(abscisic acid,ABA)在响应水分胁迫中扮演重要角色[5-7]。干旱胁迫下,植物合成大量ABA,促使叶片气孔关闭以减少水分散失,促进根系吸收水分,调节ABA响应基因并改变基因表达模式,增强植株抵抗逆境的能力[8-9]。研究显示,植物对干旱胁迫响应的信号转导基因中,约10%的基因受ABA 调控[10-11]。
干旱胁迫会干扰内源激素平衡从而阻碍植物的正常生长发育,通常情况下植物能通过自身的调节机制减轻外界胁迫带来的影响。但这种机制具有滞后性,往往不能满足植物对外界环境变化的要求而使植物受损害,特别在一些抗胁迫能力弱的作物品种上表现得尤为突出。外源植物生长调节剂能在一定程度上弥补上述调节机制的缺陷,从而提高植物耐受胁迫的能力[12-14]。研究证实,干旱胁迫下外施ABA 后,植物可溶性糖和Pro 等渗透调节物质含量上升,细胞渗透势下降,保水能力增强,有利于根系吸收水分和降低膜脂过氧化程度,保护细胞膜结构完整性[5, 7]。ABA预处理可以增强植物在应对干旱[13]、低温[14]、盐害[15]、重金属[16]胁迫条件下的抗氧化防护能力。甘蔗生长期长达12~18 个月,受季节性干旱威胁严重,若甘蔗苗期发生旱情,则会直接影响后续分蘖和伸长,导致减产。本研究拟在干旱胁迫下,分析外源ABA 对苗期甘蔗生理特性和抗氧化酶編码基因表达的影响,探讨外源ABA 提高甘蔗幼苗抗旱性的生理生化机理。
1 材料与方法
1.1 材料
甘蔗品种为‘桂糖42 号(GT42),采用桶栽土培方式,塑料桶规格为直径32 cm×高35 cm,每桶装土18.5 kg( 粘壤土, 土壤最大持水量41.6%,有机质含量18.7 g/kg,全氮含量85.6 g/kg,有效氮含量89.4 mg/kg,有效磷含量63.2 mg/kg,有效钾含量98.4 mg/kg,pH 5.8),桶底钻孔以增强透气性。单芽蔗种用甲基托布津浸种,于泥沙混合培养基质上先育苗,30 d 后,选取生长一致的甘蔗苗移栽至桶中,每桶2 株,将桶置于智能温室,按照日常管理。待植株生长至5~7 片完全叶时,设CK(正常淋水)、CK+ABA、干旱、干旱+ABA 共4 组处理。在进行干旱胁迫前连续2 d叶面喷施浓度为15 μmol/L 的ABA 溶液,程度为叶面湿透无滴水,其余2 组喷清水作对照,每个处理10 桶,3 次重复。正常淋水和干旱处理土壤含水量分别为最大持水量的60%~70% 、30%~40%,采用称重法进行控水。干旱处理开始后每隔3 d 和6 d 分别测定+1 叶的相对含水量和叶绿素荧光参数。在干旱处理后18 d,测定叶绿素荧光参数后,采集相应+1 叶样品,液氮速冻后,放入–80 ℃冰箱保存待用。
1.2 方法
1.2.1 生理生化指标测定 采用饱和含水量法测定叶片相对含水量[17];叶绿素、脯氨酸(Pro)、丙二醛(MDA)和过氧化氢(H2O2)含量分别按照孙哲等[18]、BATES 等[19]、HEALTH 等[20]和JALEEL 等[21]的方法进行测定;使用羟胺氧化法测定超氧阴离子自由基(O2–)产生速率[22];按照IRITI 等[23]的方法测定脱落酸(ABA)和生长素(IAA)含量;使用苏州科铭生物技术有限公司商品化试剂盒测定过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、抗坏血酸过氧化物酶(ascorbateperoxidase, APX)、过氧化物酶(POD)的活性,操作步骤按说明书。
1.2.2 叶绿素荧光参数测定 选取长势一致植株的+1 叶, 使用FMS-2 脉冲调制式荧光仪(hansatech, UK)测定叶绿素荧光参数。将叶片充分暗适应后,先打开测量光测定最小荧光Fo,后打开饱和脉冲光测定最大荧光Fm,再启动1000 μmol/(m2·s)的作用光,每隔20 s 测定一次光下的荧光参数,以时间为横坐标,描绘光系统II的实际光化学效率ΦPSII 启动变化曲线。
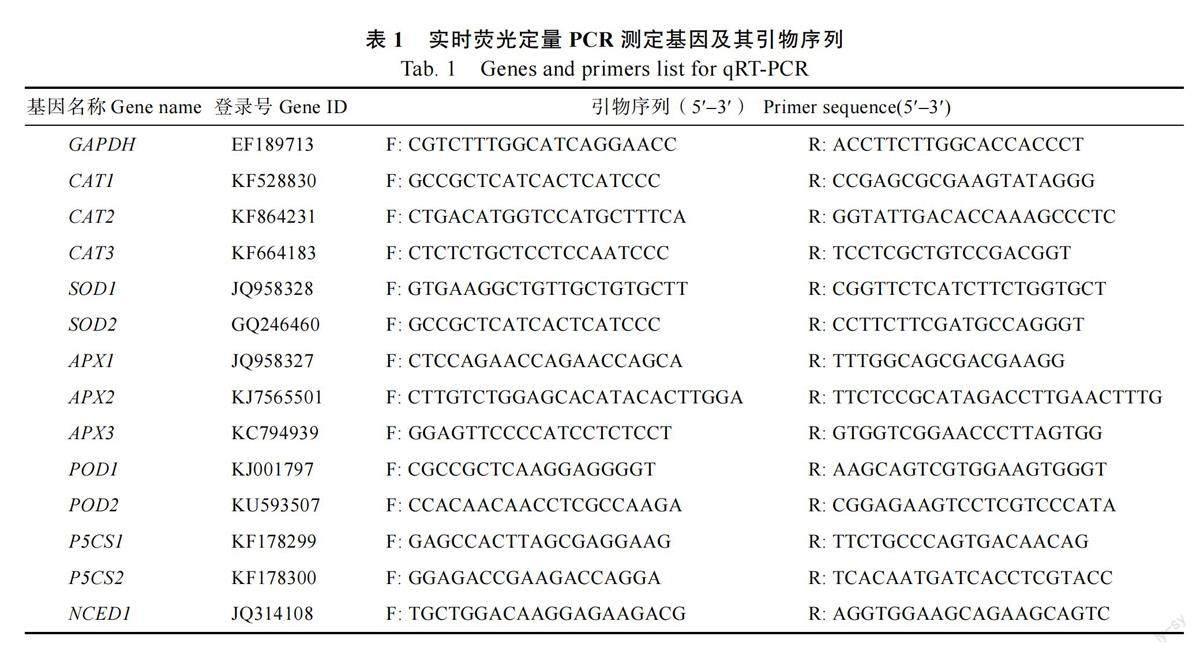
1.2.3 基因表达量测定 以甘蔗GAPDH 为内参基因,设计特异引物(表1),测定抗氧化酶CAT、SOD、APX、POD 编码基因、ABA 合成限速酶9-顺式-环氧类胡萝卜素双加氧酶(NCED)编码基因、脯氨酸合成关键酶△1-吡咯啉-5-羧酸合成酶(P5CS)编码基因相比对照的表达量变化。采用TRIzol (Invitrogen)试剂提取叶片总RNA,逆转录为cDNA,进行基因表达量测定。反应体系为:10 μL 2×All-in-One qPCR Mix (Gene Copoeia)、2 μL cDNA、4 μmol/L 的正反向引物各1、6 μL双蒸水。反应程序为:95 ℃预变性10 min;95 ℃变性10 s,60 ℃退火20 s,72 ℃延伸20 s,40 个循环。经熔解曲线检验后采用2–ΔΔCT 法进行表达量计算[24]。
1.3 数据处理
用DPS 软件以Duncans 新复极差法(P≤0.05)进行各处理平均数差异显著性检验。
2 结果与分析
2.1 叶片相对含水量(RWC)变化
干旱胁迫显著降低了甘蔗叶片相对含水量(图1)。在处理的第3、6、9、12、15、18 天,分别比对照减少2.7%、9.0%、15.1%、22.5%、26.4和31.6%,降幅随干旱时间延长而增大。干旱+ABA 处理能够提高叶片相对含水量,分别比同期干旱处理增加了1.3%、4.0%、7.1%、11.1%、11.6%和14.6%,增幅逐渐变大。正常淋水条件下喷施ABA 处理(CK+ABA)的叶片相对含水量与对照间差异不显著。
2.2 ABA和IAA含量变化
干旱、干旱+ABA 处理的ABA 含量均显著高于对照,但增量不同(图2A)。处理18 d 时,干旱处理的ABA 含量比对照增加75.2%,而干旱+ABA 处理比对照增加125.1%,干旱+ABA 处理比干旱处理增加了28.5%,差异显著。相比对照,干旱、干旱+ABA 处理均提高了甘蔗叶片的IAA含量(图2B),处理18 d 时,分别比对照提高了19.9%和18.2%,差异显著,但干旱、干旱+ABA处理间的IAA 含量差异不显著。
2.3 Pro和MDA含量变化
干旱胁迫使甘蔗叶片Pro 含量显著高于对照,而干旱+ABA 处理可进一步提高叶片Pro 含量,胁迫18 d 时干旱+ABA 处理的Pro 含量比干旱处理增加了23.2%,差异显著,而CK+ABA 处理和对照间的Pro 含量差异不显著(图2C)。干旱处理提高了甘蔗叶片MDA 含量,在处理后18 d 比对照提高了68.7%,差异显著。干旱+ABA 处理的MDA 含量比对照提高了34.1%,但比干旱处理降低20.5%,差异显著(图2D)。正常淋水条件下喷施ABA 处理(CK+ABA)的叶片MDA 含量与对照差异不显著。
2.4 叶绿素含量变化
干旱胁迫显著降低了叶绿素a 和叶绿素b 的含量,而干旱+ABA 处理能够显著缓解叶绿素的降解,胁迫18 d 时干旱+ABA 处理的叶绿素a 和叶绿素b 含量分别比干旱处理增加19.7%和25.5%。CK+ABA 处理的叶绿素含量与对照间差异不显著(图2E、图2F)。
2.5 光系统Ⅱ实际量子效率(ΦPSⅡ)变化过程
由图3 可知,干旱处理降低了叶片光系统Ⅱ实际量子效率ΦPSⅡ,破坏程度随着干旱胁迫时间延长而加重,干旱+ABA 处理能减轻这种破坏作用,使光系统Ⅱ实际量子效率ΦPSII 始终高于干旱处理。CK+ABA 处理和CK 間的光系统Ⅱ实际量子效率差异不显著。
2.6 活性氧(ROS)含量变化
干旱、干旱+ABA 处理的ROS 含量均显著高于对照(图4)。处理18 d 时,干旱处理的O2–产生速率比对照增加73.4%,而干旱+ABA 处理比对照增加44.8%,后者比前者降低16.5%,差异显著(图4A)。干旱、干旱+ABA 处理均显著提高甘蔗叶片H2O2含量,处理18 d 时,分别比对照提高82.3%和38.7%,但后者H2O2含量比前者降低了23.9%(图4B),差异显著。
2.7 抗氧化酶活性变化
干旱胁迫处理18 d 时,甘蔗叶片CAT、SOD、APX、POD 的活性分别比对照提高52.1%、23.9%、40.1%和12.2%,而干旱+ABA 处理分别比对照提高113.6%、41.4%、43.7%和38.8%,差异显著(图5)。同时期干旱+ABA 的相关抗氧化酶活性比干旱处理分别增加40.5%、14.1%、2.5%和23.7%,除APX 外,活性均比干旱处理显著增加(图5)。
2.8 基因表达量的变化
由图6 可知,处理18 d 时,正常淋水条件下喷施ABA 处理(CK+ABA)对甘蔗叶片抗氧化酶CAT、SOD、APX、POD 编码基因、NCED1 基因、P5CS 基因的表达量影响不显著。除CAT3 和APX1外,干旱胁迫提高了相关编码基因的表达量,其中NCED1、P5CS1、P5CS2、CAT1、CAT2、SOD1、SOD2、APX2、APX3、POD1 和POD2 的表达量分别是对照的3.38 倍、2.21 倍、2.32 倍、1.67 倍、1.42 倍、2.33 倍、2.17 倍、1.55 倍、1.37倍、1.56 倍和1.49 倍,差异显著。相比干旱处理,干旱+ABA 处理的NCED1、CAT2、CAT3、SOD2、APX1、APX2 和POD1 的表达量分别提高了58.3%、54.2%、57.7%、70.1%、16.9、26.5%和37.2%,差异显著。
3 讨论
干旱胁迫条件下,植物内源ABA 的积累会促进叶片气孔关闭,减少水分蒸发,提高叶片相对含水量[8-9, 25]。本研究中,随着胁迫加剧,干旱+ABA 处理甘蔗的叶片相对含水量比干旱处理显著增加,表明ABA 处理能提高干旱条件下甘蔗叶片的保水能力。研究表明,干旱胁迫会显著提高植物内源ABA 含量,而胁迫下外源ABA 处理能够进一步促进ABA 的合成和积累,使其含量处于更高水平[25-26]。本研究结果显示,干旱处理18 d时甘蔗叶片ABA 含量比对照显著增加,而干旱+ABA 处理的增幅又显著高于干旱处理,与前人研究结果一致。研究显示,9-顺式-环氧类胡萝卜素双加氧酶(9-cis-epoxy carotenoid dioxygenase,NCED)催化类胡萝卜素氧化裂解生成黄质醛,是植物内源ABA 生物合成途径最重要的限速步骤,NCED 基因表达可被非生物胁迫强烈诱导,表达量与ABA 含量有着协同效应[22]。本研究结果表明,干旱处理18 d 时NCED1 表达量为对照的3.38 倍,而干旱+ABA 又比干旱处理提高了58.3%,与ABA 含量变化一致。研究表明,ABA对植物众多生理生化途径的调控作用与其他内源激素代谢密切相关[25-27]。本研究显示,干旱胁迫下甘蔗叶片IAA 含量随着ABA 含量显著上升,但干旱+ABA 处理的IAA 含量与干旱处理间无显著差异,显示了ABA 和IAA 在响应逆境胁迫时的重要联系,但还需进一步研究才能明确这2 种激素的相互调控作用。
植物生长代谢过程中ROS 的产生不可避免,自稳态水平的ROS 在植物正常生长发育及应对环境胁迫中起着重要作用,干旱胁迫常导致植物细胞ROS 含量急剧上升[28]。本研究结果显示,干旱处理18 d 时甘蔗叶片的O2–产生速率和H2O2含量分别比对照显著增加73.4%和82.3%,表明此时植物抗氧化防护系统已无法及时清除过量的ROS 而导致其积累。过量的ROS 积累打破了植物体内的氧化还原平衡,引起膜质过氧化作用,膜脂中不饱和脂肪酸过氧化产生MDA,导致膜系统被破坏,MDA 含量高低反映了膜脂过氧化程度[25, 29]。前人研究表明,逆境脅迫下外源ABA 处理可以诱导植物抗氧化酶基因的表达,提高相关抗氧化酶的活性[30-32]。本研究结果表明,同时期干旱+ABA处理的CAT、SOD、POD 活性均比干旱处理显著提高,同时CAT2、CAT3、SOD2、APX1、APX2和POD1 基因的表达量也比干旱处理显著增加。相关抗氧化酶活性的提高,增强了植物清除过量ROS 的能力,降低膜脂过氧化程度,保护膜结构的完整性。本研究结果表明,干旱+ABA 处理的ROS 和MDA 含量虽比对照显著增加,但仍然显著低于干旱处理,证实外施ABA 处理能有效缓解干旱胁迫对膜系统的损害。研究表明,Pro 作为重要的渗透调节物质,干旱胁迫下Pro 含量增加有利于维持渗透调节,增强细胞膜系统稳定性及过量ROS 的清除[4, 33]。本研究中,干旱、干旱+ABA 处理的Pro 含量均比对照显著性增加,但干旱+ABA 处理的增量水平高于单纯干旱处理,说明ABA 处理能进一步加强干旱胁迫下Pro 的生物合成,将有助于增强叶片保水能力,提高叶片相对含水量。干旱、干旱+ABA 处理Pro 合成关键调控酶基因P5CS1 和P5CS2 的表达量比对照显著提高,与Pro 含量变化一致。但这2 个基因的表达变化在上述两处理间差异不显著,与Pro 含量变化不一致,这可能与基因的转录后修饰有关,还需进一步研究证实。
光合作用是作物生长和产量形成的重要代谢过程,干旱胁迫会导致植物叶绿体膨胀,顺序排列紊乱,基质片层模糊,基粒间连接松弛,类囊体层肿胀或解体,致使叶绿素含量下降,过量的ROS 积累还会破坏光合器官的超微结构,影响光合作用过程[34]。本研究发现,干旱处理使甘蔗叶绿素含量显著降低,而喷施外源ABA 处理能够减缓干旱条件下叶绿素的降解程度,与前人的研究结果一致[18, 35]。干旱胁迫下外源ABA 处理提高了相关抗氧化酶的活性,增强了过量ROS 的清除能力,减轻了ROS 的积累对光合作用系统造成的伤害,此外,干旱+ABA 处理Pro 含量显著提高,叶片保水能力显著增强,可能是该处理甘蔗叶绿素含量、光系统Ⅱ实际量子效率(ΦPSⅡ)始终高于干旱处理的重要原因。
