施氮量对高粱籽粒灌浆及淀粉累积的影响
2023-05-11媛王劲松董二伟刘秋霞武爱莲焦晓燕
王 媛王劲松董二伟刘秋霞武爱莲焦晓燕,*
1山西农业大学资源环境学院, 山西太原 030031;2高粱遗传与种质创新山西省重点实验室, 山西晋中 030600
氮是保证植株正常生长和籽粒发育的一个重要因素[1]。植物通过氮信号网络感知体内外氮状态, 协调各种代谢过程及源库关系[1-2]。适宜氮素通过影响细胞分裂素的合成提高籽粒库容量[3], 促进籽粒淀粉合成和籽粒灌浆[4-6]。但当土壤氮供应不足时, 氮加速从茎叶向籽粒转运[7], 导致叶片Rubisco酶活性降低[8], 影响冠层的同化能力[9-10], 最终影响籽粒灌浆[8]。氮素的奢侈吸收会延缓叶片源的衰老, 延长光合作用的持续时间, 增加光合产物积累[11]。然而, 亦有研究认为, 延缓叶片的衰老会延长源库生命周期,引起源库之间的竞争, 反而不利于籽粒的发育[12-13]。因此, 研究不同氮素水平下植株源库容量的差异对籽粒发育的影响, 即不同施氮水平下籽粒生长发育过程(即籽粒灌浆动态)可为促进作物籽粒灌浆和产量形成提供理论基础。
高粱[Sorghum bicolor(L.) Moench]是世界第五大作物, 具有较强的耐低氮能力[14]。高粱根系具有分泌生物硝化抑制剂的能力, 使其氮素利用效率高于其他作物[15]。虽然施氮可以有效提高高粱的穗粒数和粒重[16-17], 但不同施氮水平下高粱籽粒生长发育过程尚不清楚。在籽粒灌浆阶段, 圆锥花序轴上优先开花的小穗上籽粒生长迅速, 单粒重较高, 称为优势粒; 而开花较晚小穗的籽粒生长缓慢, 单粒重较低, 称为劣势粒[18]。优势粒和劣势粒在籽粒灌浆过程中并不同步, 最终形成的籽粒亦存在明显的差异, 探究不同穗位的籽粒形成规律, 有助于深入了解环境对作物灌浆过程及产量形成的影响[19-20]。此外, 在已有描述作物籽粒灌浆模型中, Richards模型对生物生长过程描述适用性较强, 利用该此模型可获得籽粒灌浆活跃期、平均灌浆速率及最大灌浆速率等指标来反应籽粒灌浆过程[21]。因此, 借助Richards模型分析不同氮素水平下高粱优势粒和劣势粒的灌浆规律, 有助于进一步剖析氮对高粱产量形成的影响。
淀粉是高粱籽粒加工的重要组成部分, 对出米率、适口性及酿造等方面都有重要的影响。籽粒灌浆的过程从根本上说就是籽粒淀粉的积累过程[22],其最终累积量与籽粒灌浆程度紧密相关[23-24]。淀粉累积过程中有4种关键酶参与蔗糖向淀粉的转化,其对淀粉的积累至关重要[25-28]; 其中ADP-葡萄糖焦磷酸化酶 (AGPase)作用于ADP-葡萄糖的形成,是淀粉合成过程的限速酶[29-30]。淀粉合酶, 包括可溶性淀粉合酶(SSS)和颗粒结合淀粉合酶(GBSS),促进淀粉链的延长[31-32]。淀粉分支酶(SBE)作用于淀粉链上, 切割α-1,4糖苷键连接的葡聚糖, 并将其形成的α-1,6糖苷键连接的葡聚糖片段重新结合到淀粉链上, 形成支链淀粉[33-34]。虽然决定籽粒淀粉合成的关键酶活性与淀粉形成的关系已有报道[31,35-36],但尚不清楚氮是否通过调控淀粉合成关键酶活性会影响高粱籽粒淀粉积累的研究, 为此本研究采用Richards方程对不同氮素处理下高粱穗部不同部位籽粒灌浆和淀粉积累过程进行拟合, 进一步分析不同施氮水平下籽粒中AGPase、SSS、GBSS和SBE酶活性与淀粉累积的相关联性, 以期明晰不同氮素水平对灌浆特征及籽粒淀粉累积的影响, 以期为高粱氮肥管理提供理论依据。
1 材料与方法
1.1 试验设计
试验于2019—2020年在山西农业大学东阳试验基地(37°33′21″ N, 112°40′2″ E)进行。该区属北温带大陆性气候, 海拔800 m, 年均气温9.7℃, 年均降水量440.7 mm, 连续12年生育期降水量均值为351.7 mm, 2019年(枯水年)和2020年(丰水年)生长季的总降雨量分别为214.2 mm和456.8 mm (图1)。土壤类型为石灰性褐土, 土壤质地为壤质沙土(各粒级体积百分比分别为: 小于0.002 mm黏粒占1.63%;0.020~0.002 mm 粉粒占11.29%; 2~0.02 mm 沙粒占87.09%)。2019年播前土壤含有机质16.51 g kg-1、全氮0.75 g kg-1、速效磷3.31 mg kg-1、速效钾129.08 mg kg-1。
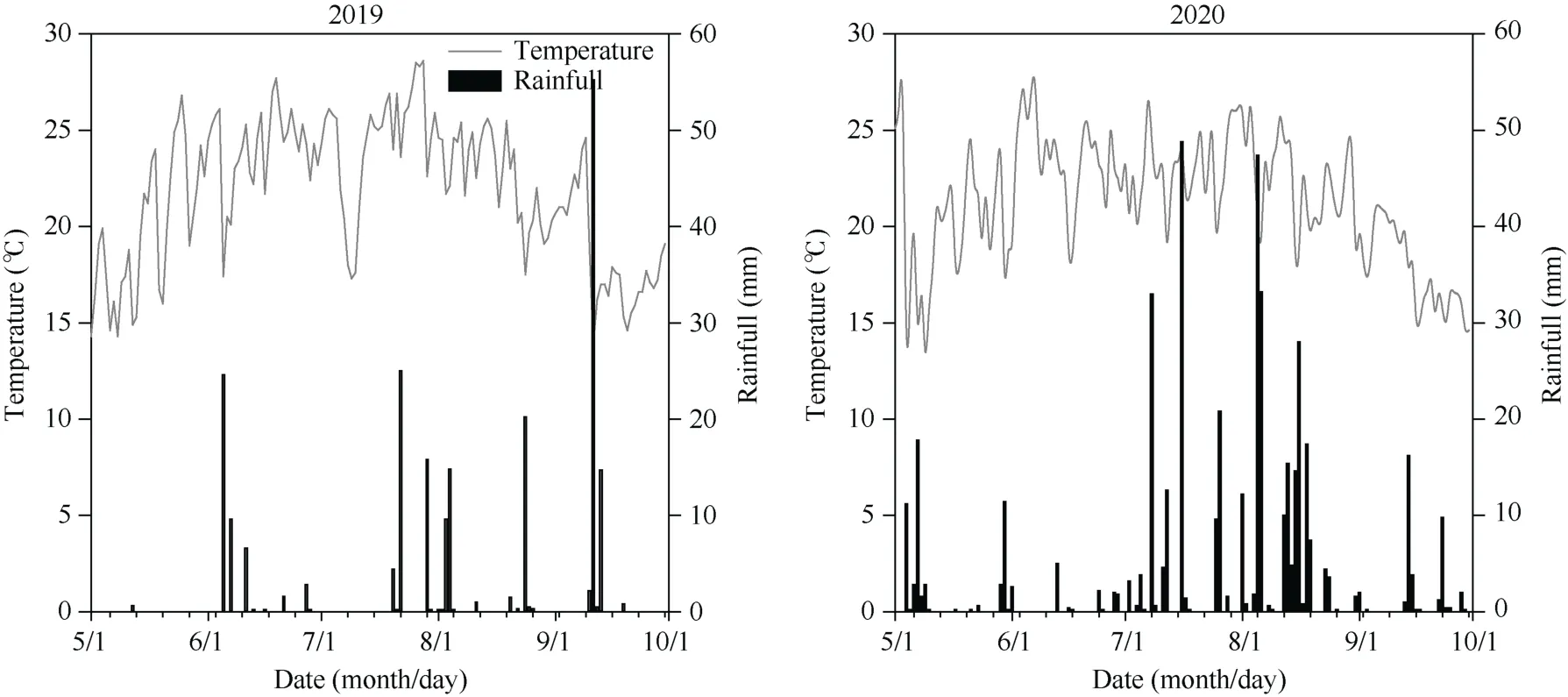
图1 2019年和2020年生长季日平均气温和降雨量Fig. 1 Average daily temperature and rainfall in 2019 and 2020 growing seasons
试验采取随机区组设计, 以当地的主推品种“汾酒粱1号”为研究对象, 设0、75、150、225、300、450 kg N hm-26个氮素水平, 各处理磷、钾施用量一致, 即施磷(P2O5) 75 kg hm-2、钾(K2O) 30 kg hm-2,氮、磷和钾分别以缓释尿素、过磷酸钙和硫酸钾全部作为基肥施入, 不再追肥。2年均在施肥前和拔节期各灌溉60 mm。小区面积48 m2(6 m × 8 m), 每个施氮水平设处理重复3次, 小区间间隔1.5 m以防止氮的运移。2019年和2020年均在5月2日播种, 分别于2019年于9月30日收获, 2020年于10月1日收获。在三叶期间苗至设计密度, 即密度为1.8×105株 hm-2。其他田间管理按高产田进行, 控制杂草、病虫害的发生, 确保除氮水平外, 没有其他因素限制生长。
1.2 样品采集
试验小区的一半设为采样区, 另一半用于收获计产和考种。各处理物候期有所差异, 去除边际效应后各小区选取同日始花, 长势一致的高粱100株进行标记。高粱整个花期约6~7 d, 穗上部先开花,始花2 d内开花的小穗发育形成的籽粒为优势粒,始花5~6 d期间开花的小穗发育形成的籽粒为劣势粒。从花期到成熟期(花后58 d)每隔7 d每小区取样9株用于采集优势粒和劣势粒, 所采籽粒在70℃烘干至恒重, 测定单粒重、单粒体积和淀粉含量。2019年同时将部分优势粒在液氮中冷冻2 min后, 贮存于-80℃超低温冰箱内用于籽粒淀粉合成关键酶酶活的测定。
1.3 测定项目及方法
1.3.1 产量测定 收获时将采集籽粒前预留的面积约为12 m2区域作为收获区, 单独计产; 同时取具有代表性植株10穗带回实验室进行拷种。
1.3.2 籽粒体积和密度测定 用95%酒精沉降法测定籽粒体积[37]。籽粒密度计算采用如下公式:
1.3.3 籽粒淀粉含量测定 将籽粒研磨过0.178 mm筛后, 采用Megazyme International Ireland Ltd.(Bray Co., 爱尔兰) 的试剂盒测定淀粉含量, 换算至单粒淀粉累积量(mg)。
1.3.4 籽粒灌浆及淀粉累积参数拟合 灌浆过程或淀粉累积采用朱庆森等[21]所描述的Richards方程[38]拟合, 方程式如下:
灌浆速率或淀粉累积速率(R)根据式(2)的导数计算:
式中,W为籽粒重或淀粉重(mg),A为最大籽粒重或最大籽粒淀粉重(mg),t为花后时间(d),B、k、N为回归系数。灌浆活跃期定义为达到籽粒最终重量或最终淀粉累积量(A)的5% (t1)~95% (t2)所用的天数, 灌浆速率中的时间(t)定义为花后第t1天至t2天, 而淀粉累积速率中的时间(t)定义为每隔7 d的花后采样天数。
1.3.5 籽粒淀粉合成相关酶提取和酶活测定 参照Nakamura等[33]的方法对籽粒中的AGPase、SSS、GBSS和SBE在4℃下进行提取, 提取液为含8 mmol L-1MgCl2, 5 mmol L-1DTT, 2 mmol L-1EDTA,12.5% (v/v)甘油和5% (w/v) PVP-40的100 mmol L-1HEPES-NaOH缓冲液(pH 7.6)。将20粒高粱籽粒在5 mL提取液中均质, 后在12,000×g离心10 min, 取上清液用于AGPase、SSS和SBE测定。沉淀部分用5 mL提取液重新混合后, 用1500×g微离心3 min,取上层混悬液用于GBSS测定。
参照Nakamura等[33]的方法测定AGPase和SBE的活性; 参照Schaffer和Petreikov[39]的方法测定SSS和GBSS的活性。所有用于酶学检测的化学物质和酶均来自Sigma化学公司(美国)。
1.4 数据分析
采用 SPSS 19.0软件进行统计分析, Origin 2019和SigmaPlot 12.5作图。
2 结果与分析
2.1 不同施氮水平对籽粒产量、籽粒形态与籽粒灌浆的影响
氮肥显著影响了高粱的籽粒产量、穗粒数(图2)。施氮75 kg hm-2显著提高了2020年高粱穗粒数和产量, 与不施氮比较分别提高30.3%和20.5%。随着施氮水平的继续增加, 穗粒数和产量基本维持不变。2019年氮肥对穗粒数提升的趋势与2020年基本一致, 施氮75 kg hm-2较不施氮提高35.1%。2019年施氮75 kg hm-2产量与不施氮虽无显著性差异, 但亦存在增加的趋势。说明与不施氮相比较, 施氮75 kg hm-2即可提高高粱籽粒产量和穗粒数。
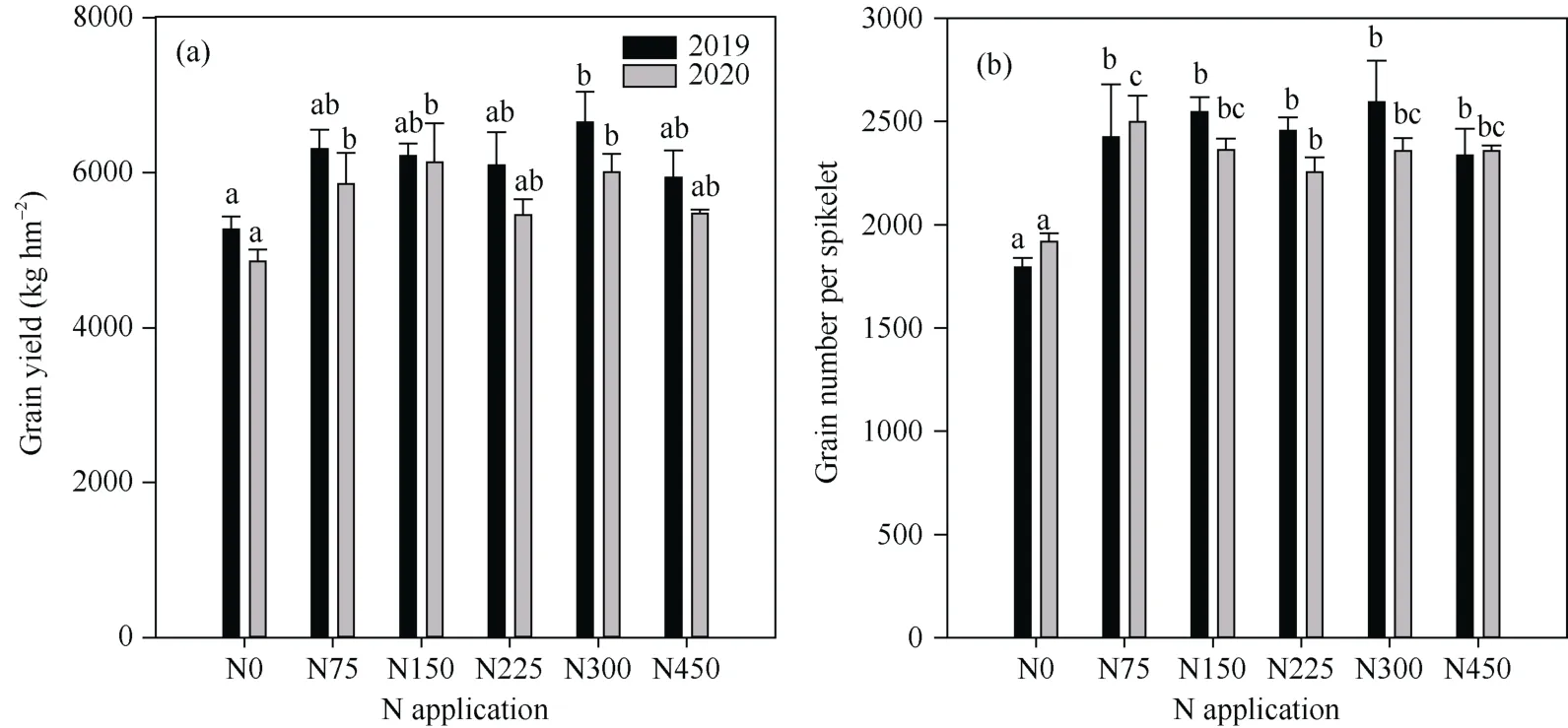
图2 不同施氮水平下的高粱产量(a)和每穗粒数(b)Fig. 2 Effect of N application on the grain yield per hectare (a) and grain number per panicle (b) of sorghum in 2019 and 2020N0: 施氮量为0 kg hm-2; N75: 施氮量为75 kg hm-2; N150: 施氮量为150 kg hm-2; N225: 施氮量为225 kg hm-2; N300: 施氮量为300 kg hm-2; N450: 施氮量为450 kg hm-2。不同字母表示同一年度不同处理间在0.05概率水平差异显著。N0, N75, N150, N225, N300, and N450 indicate the rate of 0, 75, 150, 225, 300, and 450 kg N hm-2, respectively. Bar with different letters means significant differences among treatments in the same year at the 0.05 probability level.
高粱优势粒的粒重、体积、密度和灌浆速率均显著高于劣势粒, 且具有较短的灌浆活跃期。氮素对优势粒和劣势粒的粒重、籽粒形态和灌浆特性的影响表现基本一致(表1)。与施氮75 kg hm-2比较, 缺氮(不施氮处理)或过量施氮(450 kg N hm-2)均能提高粱籽粒粒重(表1), 2019年缺氮和450 kg N hm-2处理的优势粒粒重分别提高9.14%和3.76%, 劣势粒则分别提高8.98%和5.39%。2020年高粱粒重的趋势与2019年基本一致。施氮显著提高了籽粒密度(表1), 2019年150 kg N hm-2处理的优势粒和劣势粒较不施氮处理分别提高11.25%和9.73%, 2020年则分别提高22.57%和10.25%。说明施氮有助于提高籽粒密度, 单粒重则随着施氮量的增加逐步提升, 但仍略小于不施氮。
施氮显著提高了籽粒灌浆速率(表1), 2019年施氮150 kg hm-2优势粒和劣势粒较不施氮分别提高6.16%和8.65%, 2020年分别提高11.07%和12.75%。不施氮显著延长了籽粒灌浆活跃时间(表1), 优势粒的有效灌浆期延长了3~5 d, 劣势粒延长了5~7 d。说明施氮有助于提高籽粒灌浆速率, 缩短籽粒灌浆时间。
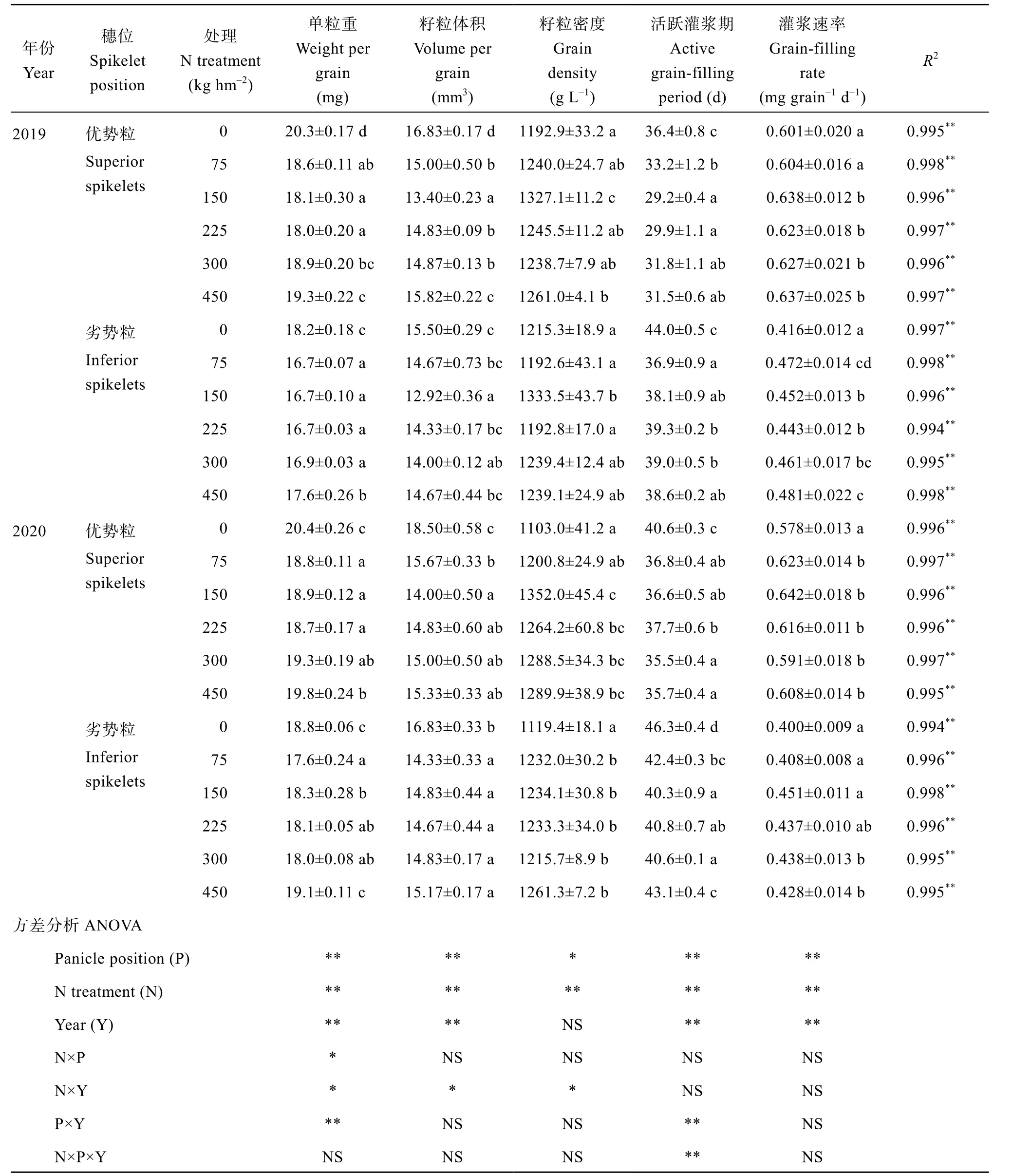
表1 不同施氮水平下高粱籽粒特点及籽粒灌浆特性Table 1 Effect of N application on the structure of sorghum grains and the characteristics of grain-filling in 2019 and 2020
2.2 不同施氮水平下的籽粒淀粉累积
施氮提高了优势粒浆灌前期的淀粉累积。但花后28 d至籽粒成熟, 不施氮处理优势粒淀粉累积量高于各施氮处理, 该时间段内不施氮处理累积的淀粉约占最终淀粉累积量的21%, 而2019年和2020年施氮处理下优势粒累积的淀粉量分别仅占最终淀粉累积量的14%~17%和12%~14% (图3-a, c)。花后35 d至籽粒成熟, 2019年不施氮处理下劣势粒淀粉累积量高于各施氮处理。2020年不施氮处理下劣势粒最终淀粉累积量次高, 略低于450 kg hm-2处理(图3-b, d)。就籽粒淀粉累积速率而言, 花后21 d至籽粒成熟, 不施氮处理优势粒高于各施氮处理(图3-e, g), 花后28 d至籽粒成熟不施氮处理劣势粒高于各施氮处理(图3-f, h), 说明缺氮促进灌浆后期籽粒淀粉累积, 同时也提高了后期淀粉累积速率,而施氮则有利于灌浆早期淀粉累积。此外, 优势粒的淀粉累积速率在2年间存在一定差异, 2020年各处理籽粒淀粉累积速率最大值的时间均早于2019年, 且N0处理籽粒淀粉累积速率峰值明显低于2019年(图3-e, g)。
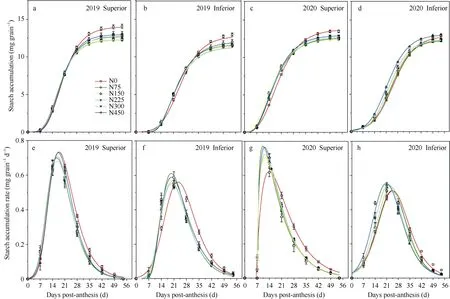
图3 不同施氮量下高粱优势粒(a, c, e, g)和劣势粒(b, d, f, h)的淀粉累积(a, b, c, d)和淀粉累积速率(e, f, g, h)Fig. 3 Effect of N application on starch accumulation (a, b, c, d) and starch accumulation rate (e, f, g, h) in superior (a, c, e, g) and inferior (b, d, f, h) spikelets of sorghum in 2019 and 2020根据Richards方程拟合淀粉累积速率。处理同图2。The starch accumulation rate was calculated according to Richards’ equation. Treatments are the same as those given in Fig. 2.
2.3 不同施氮量下籽粒淀粉合成酶活性变化
为明确氮对高粱淀粉累积过程的影响机制,2019年以优势粒为对象明确氮对淀粉合成关键酶活性的影响。施氮处理下AGPase和SSS的酶活在花后14 d时均达到最大值后迅速下降。过量施氮(450 kg N hm-2)显著提高了花后14 d和21 d的AGPase和SSS的酶活(表2)。而不施氮处理下花后14~28 d的AGPase和SSS活性较稳定(图4-a, b), 且AGPase活性在花后28 d显著高于施氮处理(表2)。不施氮下SSS活性达到峰值的时间亦延后到花后21 d (图4-b)。GBSS和SBE活性则分别在花后21 d和28 d达到最大值(图4)。过量施氮(450 kg N hm-2)显著提高了花后14 d和21 d GBSS的活性(表2), 不施氮处理则在花后35~49 d活性最高(图4-c)。说明缺氮显著提高了灌浆后期AGPase和GBSS的酶活性, 而施氮显著提高了灌浆前期AGPase、SSS和GBSS的酶活性。
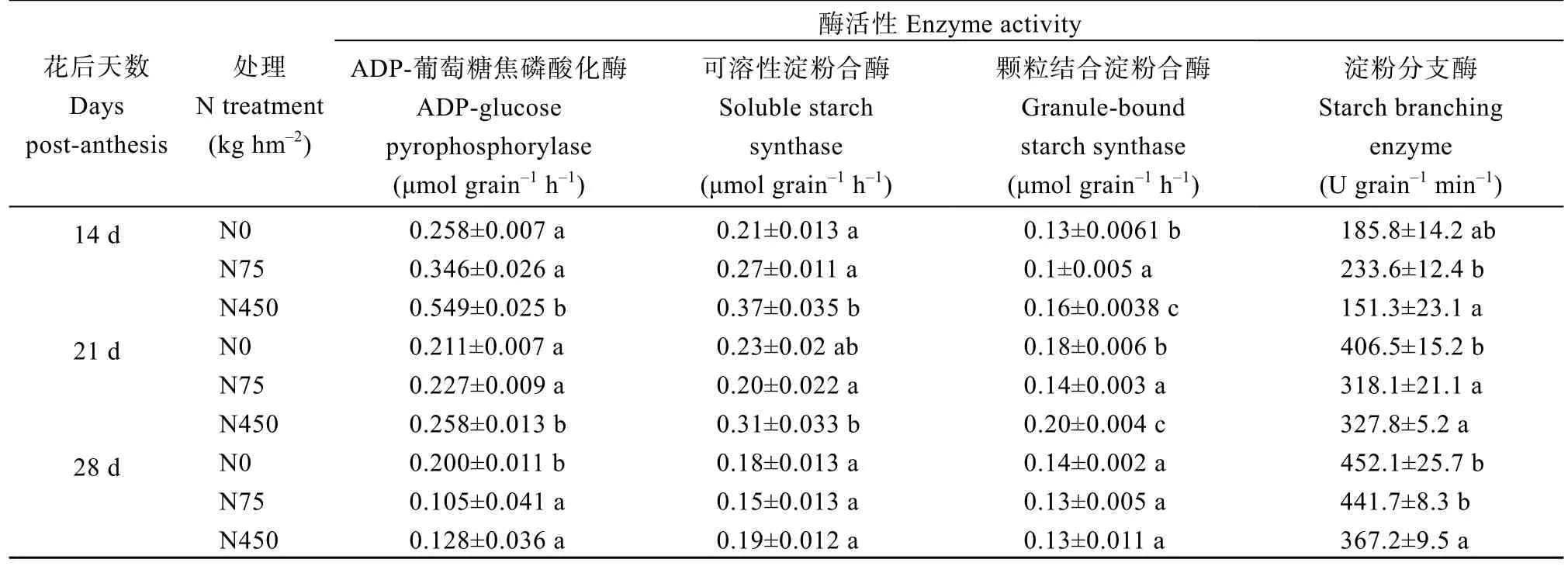
表2 花后14~28 d不同施氮量对高粱优势粒淀粉相关酶活性的影响(2019年)Table 2 Effects of N application on activity of the enzymes involved in the sucrose-to-starch conversion in the superior grains of sorghum during 14 DPA to 28 DPA in 2019
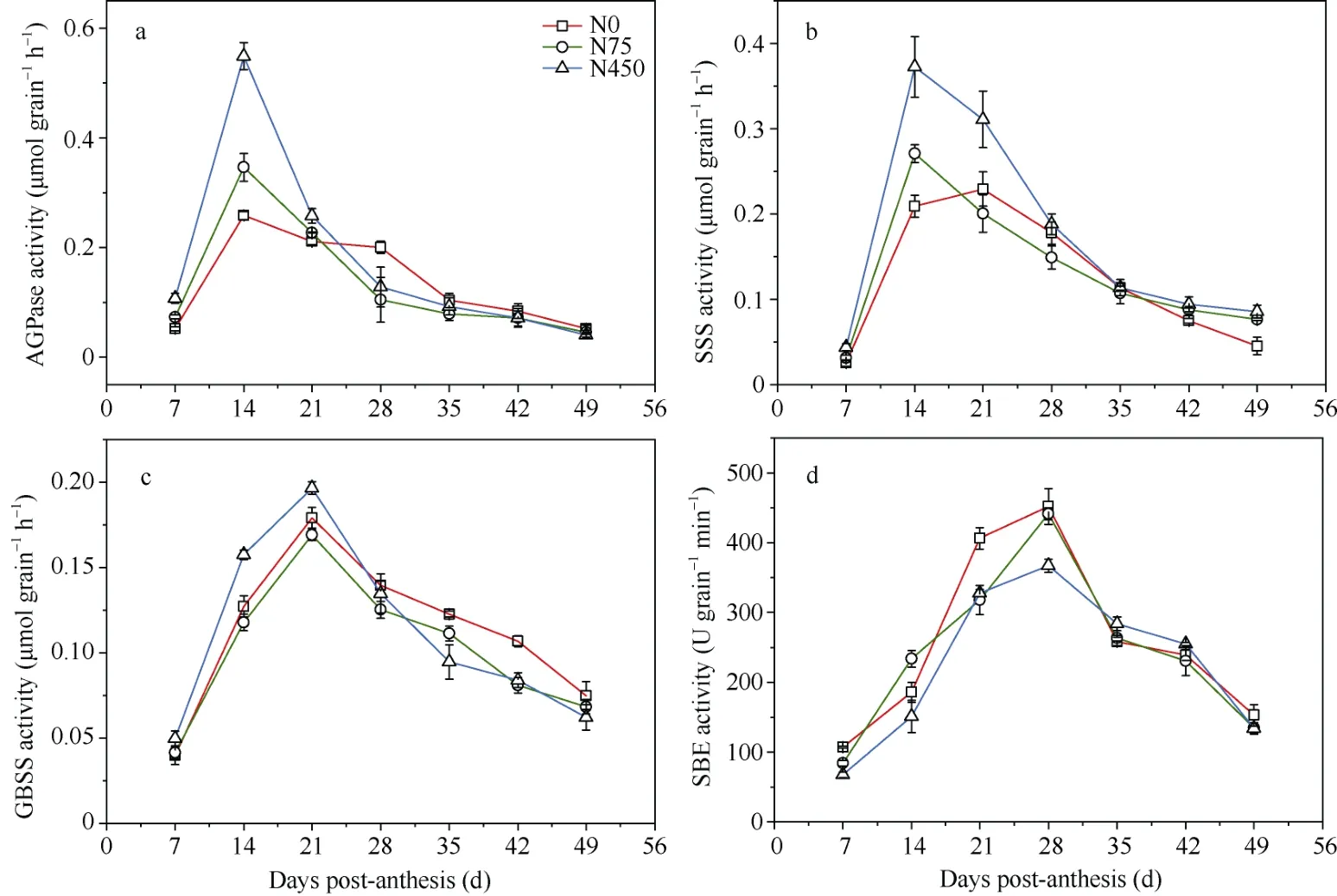
图4 不同施氮量下高粱优势粒AGPase (a)、SSS (b)、GBSS (c)和SBE (d)的酶活性(2019年)Fig. 4 Effect of N application on the activity of AGPase (a), SSS (b), GBSS (c), and SBE (d) in sorghum superior spikelets in 2019处理同图2。Treatments are the same as those given in Fig. 2.
2.4 高粱优势粒籽粒淀粉合成相关酶活性与淀粉累积速率的关系
通过分析2019年优势粒淀粉快速累积时期(即花后7、14、21、28和35 d)籽粒AGPase、SSS、GBSS和SBE的酶活性与拟合得到的当天淀粉累积速率的相关性(表3)发现, 除不施氮条件下SSS活性与淀粉累积速率无显著的相关性外, AGPase和SSS活性在其余处理下均与籽粒淀粉累积速率显著相关, 说明AGPase和SSS在高粱优势粒淀粉累积中起重要作用。
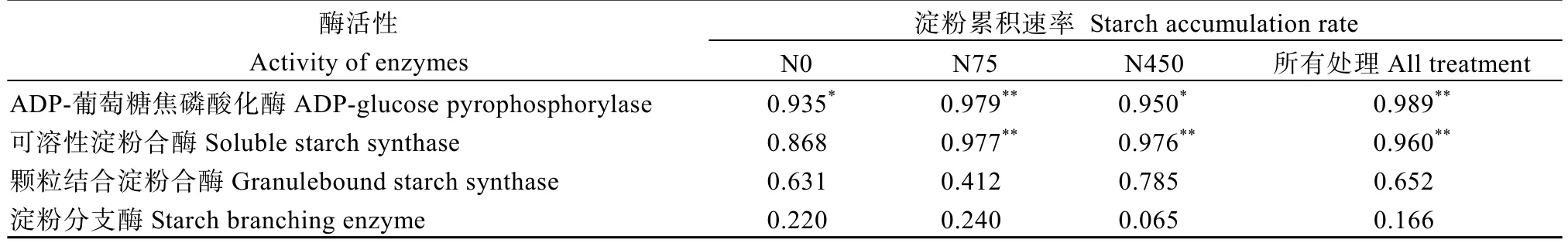
表3 高粱优势粒淀粉快速累积时期(花后7~35 d) AGPase、SSS、GBSS和SBE的酶活性与淀粉累积速率的相关性Table 3 Correlation of starch accumulation rate with the activity of enzymes involved in sucrose-to-starch conversion during the rapid starch accumulation rate period (7-35 DPA) in 2019
3 讨论
3.1 施氮量对籽粒产量构成及籽粒灌浆的影响
高粱根系能释放大量生物硝化抑制剂, 延缓土壤中的硝化过程[40], 这可能有利于高粱适应低氮胁迫, 提高氮素利用效率[4,15], 因此, 高粱达到目标产量需氮量低于玉米[42]。Schlegel等[43]在1961—2015年高粱籽粒产量对氮的响应研究发现施氮100 kg hm-2时即可达到最高产量。本研究中, 施氮75 kg hm-2的高粱产量和穗粒数最高(图2), 即达到了合理施氮量。在同一种植密度下, 作物产量的提高主要取决于穗粒数的大幅提升[44-45], 且施氮可以显著提升穗粒数[2], 这与本研究结果基本一致。
通过对水稻籽粒灌浆和籽粒淀粉累积的拟合表明, Richards方程对籽粒生长曲线的拟合适应性较强[20,46], 本试验亦得到良好的拟合效果(表1)。籽粒重由籽粒灌浆时间和籽粒灌浆速率决定[4,47], 研究表明施氮可提高籽粒灌浆速率, 缩短籽粒灌浆时间, 增加粒重[48-49]。亦有研究发现, 由于源强度的降低, 缺氮导致了小麦籽粒重的降低[50], 但Acreche和Slafer[51]的研究表明, 水稻的平均粒重随穗粒数的增加而降低, 缺氮对籽粒重的影响可能取决于氮胁迫下穗粒数的降低程度和源的降低程度二者之间的平衡[50]。本研究中, 缺氮导致穗粒数大幅降低(图2), 灌浆期延长, 灌浆速率降低(表1), 两者平衡后籽粒重显著高于所有施氮处理(表1)。此外, 关于小麦和水稻的研究发现灌浆速率的降低和灌浆期的延长, 不仅会影响籽粒产量, 进一步还会使籽粒品质下降, 尤其是碾磨品质和籽粒外观[49,52]。本研究中,缺氮使优势粒的有效灌浆期延长3~5 d, 劣势粒延长了5~7 d (表1), 高粱产量降低。高粱产区还可能发生早霜, 在缺氮环境下, 籽粒灌浆期的延长会增加籽粒成熟无法成熟的风险, 尤其是弱势粒。因此, 为抵御低温伤害带来籽粒不成熟的风险, 推荐在较为贫瘠的土壤条件下应使用早熟品种。
3.2 不同施氮处理下高粱籽粒淀粉累积与酶活性
籽粒灌浆实际上就是籽粒淀粉累积的过程。本研究中氮素对优势粒和劣势粒的粒重、籽粒形态、灌浆特性和淀粉累积过程的影响趋势基本一致(表1和图3), 且2年施氮75 kg hm-2、150 kg hm-2、225 kg hm-2和300 kg hm-2处理的籽粒淀粉积累量和累积过程比较相似, 但不施氮、施氮75 kg hm-2和施氮450 kg hm-2处理的淀粉积累存在明显的差异(图3)。加之, 年际间不同施氮水平对籽粒淀粉累积的影响亦基本一致(图3)。故为探明不同施氮处理下淀粉累积差异的机制, 本研究在2019年测定了不施氮、施氮75 kg hm-2和施氮450 kg hm-2下优势粒中AGPase、SSS、GBSS和SBE的酶活性。
与不施氮处理相比, 施氮处理籽粒淀粉累积速率在灌浆早期增加较快, 达到最大值后下降却较快(图3), Wei等[53]在玉米上也有类似的结果。氮素有助于AGPase、SSS和GBSS活性的提高[54]。大麦的研究中发现, 随着施氮量的增加, 淀粉合酶活性增加, SBE酶活性降低[55]。Yang等[48]认为与淀粉合成相关酶活性的提高有利于增强库强度。本研究中,AGPase和SSS的活性与籽粒淀粉累积速率显著相关。施用氮肥有助于灌浆早期这2种关键酶酶活的提高, 可能会增加灌浆早期高粱籽粒库强度, 从而提高灌浆早期籽粒淀粉累积速率(表3和图4)。作物籽粒灌浆是由绿叶光合能力(源强度)和同化产物储存能力(库强度)共同决定的。氮素主要在灌浆早期影响细胞分裂素的合成, 而细胞分裂素在源库关系中起着重要的调控作用[56], 可导致灌浆前期较大的库强[56-57]。细胞分裂素还可通过延缓源叶衰老和延长光合作用持续时间来增强源强度[11]。AGPase是光合产物转化为淀粉合成引物(ADP-葡萄糖)过程的关键酶[29-30]。施氮有助于灌浆早期AGPase酶活的提高,可能是由于灌浆前期, 源库强度的提升加速了光合产物从营养器官向籽粒的运输和籽粒蔗糖向淀粉的转化。过量施氮(450 kg N hm-2)则较施氮75 kg hm-2进一步强化了此过程, 诱导淀粉累积继续增加。花后0~7 d, 淀粉积累不受氮素供应的影响, 说明此阶段籽粒库强受到限制(图3), 小麦和水稻灌浆早期也存在类似现象[47-48]。
与施氮处理相比, 不施氮处理的籽粒灌浆中后期AGPase、SSS和GBSS活性下降较慢。不施氮下籽粒灌浆期延长, 单粒淀粉积累量增加, 单粒重显著高于施氮处理(图3、图4和表1)。这可能是由于缺氮导致植株生长缓慢, 植物体对碳水化合物的需求降低, 进而导致抽穗前茎秆淀粉和非结构性碳水化合物向籽粒的转移增加[54,58]。此外, 不施氮显著降低了穗粒数(图2), 穗粒数的下降可能提高了单个籽粒的碳水化合物再分配。而在单粒水平上, 这种再分配可能确保了足够的蔗糖用于淀粉的生物合成。籽粒密度的情况更为复杂, 缺氮处理的籽粒体积较大, 密度较低(表1)。这可能是由于抽穗后产生的同化物质不足以灌浆, 可能与缺氮情况下高粱源受限有关。花后土壤干旱胁迫可以调控籽粒淀粉的积累[46-47], 本研究中2019年和2020年分属枯水年和丰水年, 虽分别在施肥前和拔节期各灌溉60 mm,但仍可能导致了2020年较2019年各处理优势粒淀粉最大积累速率达到峰值的时间较早, 且2019年不施氮下优势粒淀粉积累速率峰值显著高于2020年(图3-e, g), 但年际间不同施氮水平对籽粒淀粉累积的影响亦基本一致。
4 结论
施氮提高了穗粒数、籽粒产量和灌浆速率, 缩短了灌浆持续时间。淀粉累积速率与参与籽粒淀粉合成的关键酶AGPase和SSS显著相关。过量施氮(450 kg N hm-2)灌浆前期籽粒中AGPase和SSS的活性最高, 促进了灌浆前期籽粒淀粉累积; 施氮75 kg hm-2灌浆前期籽粒中AGPase和SSS的活性和淀粉累积速率次之; 虽然缺氮降低了灌浆前期籽粒中AGPase和SSS的活性, 但在灌浆后期维持较高活性而延长了灌浆活跃期, 因而后期具有较高的单粒淀粉累积速率提升了单粒淀粉累积量和单粒重。此外,由于缺氮条件下高粱籽粒灌浆期较长, 早熟品种应在土较为贫瘠的地区种植, 以减少低温造成籽粒无法完全成熟的风险。
