甘草带对棉田多异瓢虫种群发生与控害的调控功能
2023-04-26姜岩宋冰梅陈鑫李号宾王冬梅阿克旦吾外士潘洪生
姜岩,宋冰梅,2,陈鑫,3,李号宾,王冬梅,阿克旦·吾外士,潘洪生*
(1.新疆农业科学院植物保护研究所,乌鲁木齐 830091;2.新疆大学生命科学与技术学院/ 新疆生物资源和基因工程重点实验室,乌鲁木齐 830046;3.新疆农业大学农学院/ 教育部棉花工程研究中心,乌鲁木齐 830052)
储蓄植物可以通过直接为天敌提供替代食物(如:花粉、花蜜等),或间接繁殖替代猎物来维持天敌的种群繁殖,对于天敌的多样性保护和害虫的生物防治均具有重要意义[1-3]。国内外已有大量关于储蓄植物筛选与利用的研究[4]。如在萝卜(Raphanus sativus)地里设置储蓄植物香雪球(Lobularia maritima)种植区后,在香雪球开花区诱捕到的天敌数量显著高于对照区,且能够有效控制桃蚜(Myzus persicae)、黑须污灯蛾(Spilarctia casigneta)和黄斑长跗萤叶甲(Monolepta signata)等植食性害虫对萝卜的危害[5]。Chen 等[6]发现,当在番茄(Solanum lycopersicum)田内分别使用蓖麻(Ricinus communis)上的蓖麻粉虱(Trialeurodes ricini)和大豆(Glycines max)上的豌豆修尾蚜(Megoura japonica)作为替代猎物时,可分别显著增加丽蚜小蜂(Encarsia formosa)和龟纹瓢虫(Propylea japonica)的种群数量,明显减少番茄田中烟粉虱(Bemisia tabaci)的种群数量。可见,储蓄植物可以预防性地将天敌引入农田生态系统,增加天敌种群数量,达到可持续控害的效果[7]。21 世纪初,储蓄植物系统产品便已开始在欧美国家商品化,目前在比利时、德国、法国、日本、美国和加拿大等国家均得到了广泛的推广和应用。但是由于我国地理气候的多样性以及与欧美等国家农业设施的差异性,生产中并不能完全照搬国外已报道或构建的储蓄植物系统[8]。因此,亟待研究构建针对我国不同气候环境、不同作物生境和不同农业发展阶段的储蓄植物系统[9]。
2021 年,新疆棉花播种面积为250.6 万hm2,产量为512.9 万t,分别占全国的82.76%和89.50%[10]。棉花的安全生产关系到新疆棉花种植区的农业增收、增效,关系到我国农业结构的战略性调整,是新疆乃至国家发展的战略需求[11-12]。蚜虫(Aphisspp.)是棉花上的重要害虫之一[13-14],严重影响棉花的产量与品质[15]。长期以来,人们通常应用化学杀虫剂进行蚜虫防治,致使其抗药性问题愈加严重,无法得到持续性的控制[16];因此,挖掘和利用捕食性天敌的生物控害功能,成为现阶段防治棉花蚜虫的重要途径之一[17]。多异瓢虫(Hippodamia variegata)具有发生早、数量大、食量大和食性广等特点,是新疆地区蚜虫等害虫的优势捕食性天敌[18-21]。前期调查发现,新疆棉田田埂甘草(Glycyrrhiza uralensis)带上棉黑蚜(Aphis atrata)种群数量较多,对多异瓢虫具有一定的保育作用[22]。本研究对棉田田埂甘草带及距甘草带不同距离棉田蚜虫与多异瓢虫种群动态进行系统调查,同时通过罩笼法比较有/ 无甘草带条件下蚜虫种群增长倍数与多异瓢虫控害指数的差异,以期为合理利用棉田周边甘草带来有效保育多异瓢虫、促进棉田蚜虫的生物防治奠定基础。
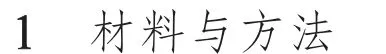

试验在国家植物保护库尔勒观测实验站(新疆巴音郭楞蒙古自治州库尔勒市和什力克乡库勒村,41.75°N,85.81°E)进行。选取田埂上有宽约1 m 甘草带(铲除田埂上除甘草以外的其他杂草,以下称“处理棉田”)和无甘草带(铲除田埂上所有杂草,以下称“对照棉田”)棉田小区各3 个,每个小区面积为800 m2(20 m×40 m),相邻2 个小区间隔20 m。棉花品种为中棉所49,采用地膜覆盖膜下滴灌栽培,膜宽150 cm,膜间距40 cm,每膜种4 行,窄行行距10 cm,宽行行距60 cm,株距10 cm。试验期间甘草带与棉田均不施用化学农药,滴灌、随水施肥等按常规措施进行统一管理。

1.2.1 甘草带及距甘草带不同距离棉花上蚜虫和多异瓢虫种群数量调查。2021 年5 月下旬至8月下旬,在所选地块采用目测法系统调查甘草带、对照棉田及距甘草带1 m、5 m、10 m、20 m 处棉花上蚜虫和多异瓢虫种群数量,每7 d 调查一次。其中:甘草带上每个小区调查5 点,每点调查面积为1 m2(1 m×1 m);棉田每个小区选取5点,苗期(6 月21 日及其以前)每点调查10 株,6月21 日后每点调查5 株。分别记录蚜虫及多异瓢虫种群数量。
1.2.2多异瓢虫对蚜虫的生物控害作用评价试验(罩笼试验)。于2021 年6 月22 日开展,所用网罩为孔径0.18 mm(80 目)的白色尼龙纱网。罩笼前首先清除选定棉株上除蚜虫外的其他害虫、天敌以及蚂蚁等,然后剔除多余的蚜虫,每株上仅保留100 头无翅成蚜。
(1)在距甘草带8 m 处,随机选取长势整齐的10 株棉花设置罩笼,每株棉花为1 组重复,共设置10 组重复,同时在距离每个罩笼约1 m 处另选10 株棉花不罩笼,相邻2 组罩笼间隔5 m以上,用于计算蚜虫种群相对增长倍数,评价多异瓢虫对蚜虫的生物控害作用。罩笼后7 d 和14 d,分别调查处理棉田和对照棉田中罩笼和非罩笼棉株上蚜虫的种群数量。
(2)在距甘草带8 m 处,随机选取长势整齐的10 株棉花设置罩笼,每株棉花为1 组重复,共设置10 组重复,同时在无甘草带田埂同样距离处的对照棉田同样设置非罩笼作为对照,相邻2组罩笼间隔5 m 以上,用于计算多异瓢虫对蚜虫的控害指数,评价多异瓢虫对蚜虫的生物控害作用。

甘草上蚜虫仅为棉黑蚜,棉花上蚜虫种群数量为棉黑蚜、棉长管蚜(Acyrthosiphon gossypii)和棉蚜(Aphis gossypii)总和,多异瓢虫种群数量为成虫和幼虫总和。甘草带上棉黑蚜和多异瓢虫种群数量均换算为每平方米虫量,棉花上则换算为每株虫量。同时,计算每次调查时对照棉田与距甘草带不同距离棉田多异瓢虫与蚜虫的益害比(NPY)。其公式为NPY=NP/NY。式中,NP与NY分别代表多异瓢虫种群密度和蚜虫种群数量。当数据方差具有齐性时,采用邓肯多重范围检验(Duncan 氏新复极差法)进行多重比较;当方差不具有齐性时,转换数据使方差具有齐性后,继续采用邓肯多重范围检验 (Duncan 氏新复极差法)进行多重比较。对盛发期距甘草带不同距离棉株上蚜虫及多异瓢虫种群数量(x)进行对数[lg(x+1)]、正 弦(sinx)、余 弦(cosx)、平 方 根转换。其中:对6 月10 日蚜虫种群数据进行余弦(cosx)转换,7 月1 日蚜虫种群数据进行正弦(sinx)转化;对6 月10 日和6 月17 日多异瓢虫种群数据进行平方根转换,7 月1 日多异瓢虫种群数据进行对数[lg(x+1)]转换。
根据罩笼试验结果,计算蚜虫种群相对增长倍数(rrg),计算公式为rrg=Nt/No。其中,Nt表示调查时间点罩笼处理或对照蚜虫数量,No表示罩笼处理或对照的起始蚜虫数量。使用天敌控害指数(INEB)来评价多异瓢虫对蚜虫的生物控制作用,计算公式为INEB=(Ac-Ao)/Ac。其中,Ac表示调查时间点罩笼处理的蚜虫数量,Ao表示同一时间点对照棉株上蚜虫数量。数据分析前对蚜虫种群密度进行对数转换[lg(x+1)],以使其满足正态分布与方差齐性。同时采用独立样本t检验比较罩笼试验中蚜虫种群相对增长倍数的差异以及棉田中多异瓢虫控害指数的差异。
所有数据均利用Microsoft Excel 2003 整理,采用SPSS 20 进行统计分析。


结果(图1)表明,5 月下旬至6 月上中旬是甘草带上棉黑蚜的盛发期,在这期间棉黑蚜种群数量波动上升,于6 月17 日达到最大值,然后骤减,7 月8 日后种群数量趋于零。甘草带上多异瓢虫在6 月盛发,其种群数量在6 月24 日达到最大值,然后逐渐下降,7 月15 日后维持在较低水平。
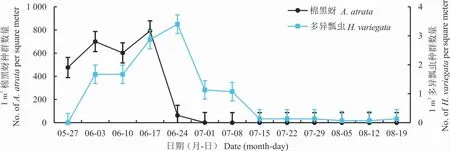
图1 甘草带上棉黑蚜和多异瓢虫的种群动态Fig.1 Population dynamics of A.atrata and H.variegata on G.uralensis strips
在对照棉田与距甘草带不同距离(1 m、5 m、10 m、20 m)的棉田中,除距甘草带1 m 和5 m 处的棉田外,蚜虫均于6 月初开始发生,然后种群数量逐渐增加,于7 月1 日达到最大值,然后逐渐降低,至8 月中旬趋近于零;而在距离甘草带1 m 处的棉田中,蚜虫种群数量在7 月22 日才达到最大值,同样于8 月中旬趋近于零。在距离甘草带1 m、5 m、10 m 的棉田中,多异瓢虫种群数量于6 月上旬开始逐渐增加,至7 月8 日达到最大值,随后逐渐下降,于8 月中旬趋近于零;在距甘草带20 m 及对照棉田中,多异瓢虫种群数量于6 月24 日达到最大值,而后种群数量逐渐下降,同样于8 月中旬趋近于零(图2)。
图2 对照棉株与距甘草带不同距离棉株上蚜虫和多异瓢虫的种群动态Fig.2 Population dynamics of aphids and H.variegata in cotton plants without G.uralensis strips and cotton plants away from G.uralensis strips at different distances
经方差分析,在蚜虫的盛发期(6 月3 日-7月1 日),6 月3 日[F(4,10)=40.504,P<0.001]和6 月24 日[F(4,10)=5.372,P=0.014]距甘草带不同距离棉株上蚜虫种群数量存在差异。多重比较结果(表1)显示:在6 月3 日距离甘草带1 m棉株上蚜虫种群数量最高,其次为距甘草带5 m处棉株,显著高于其他距离棉株;6 月24 日对照棉花上蚜虫种群数量显著高于有甘草带的棉株。
在多异瓢虫盛发期(6 月10 日-7 月8 日),除7 月1 日[方差分析结果:F(4,10)=9.057,P=0.002] 距离甘草带1 m、5 m、10 m 棉株上多异瓢虫种群数量显著大于对照棉株和距甘草带20 m棉株之外,在其余时间距甘草带不同距离棉株上的多异瓢虫种群数量与对照棉株均无显著性差异(表2)。
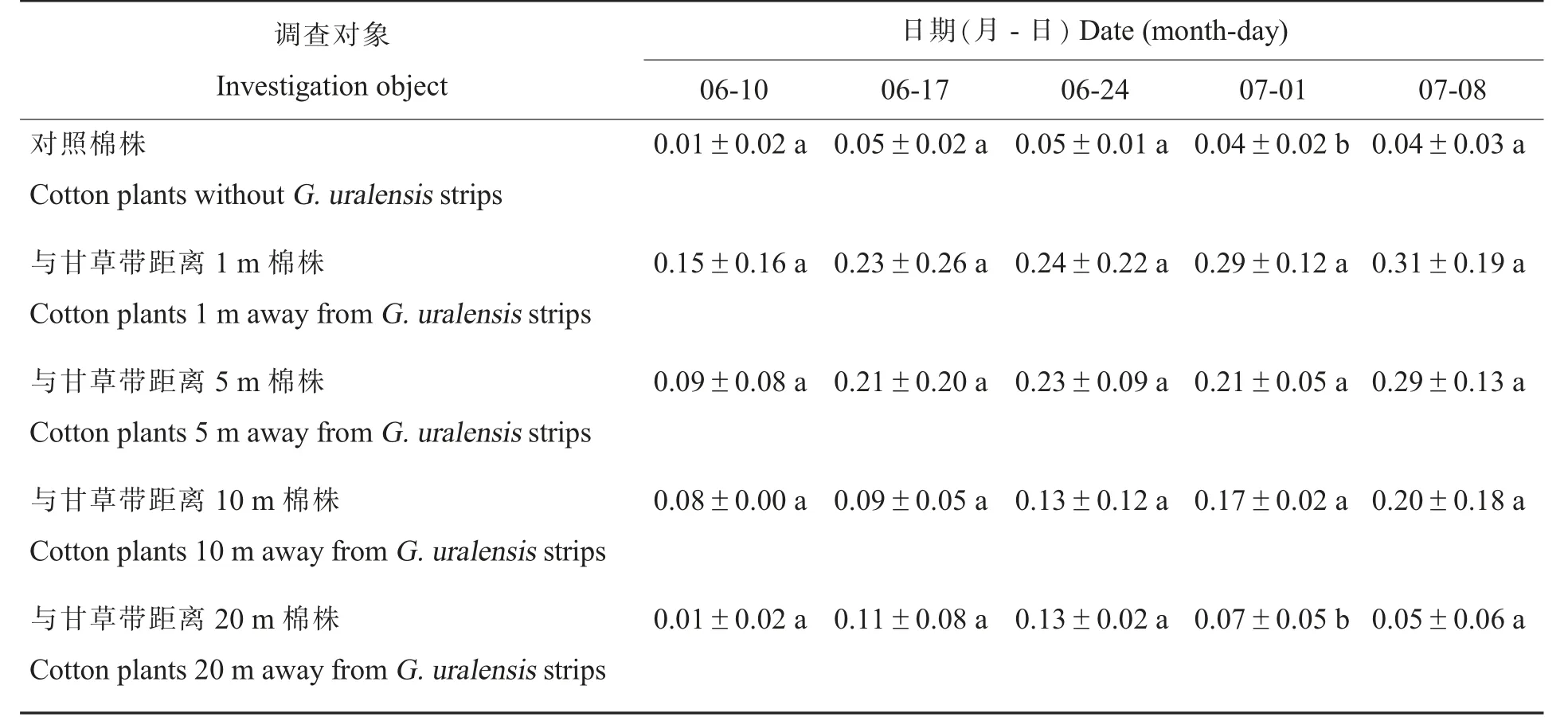
表2 盛发期距甘草带不同距离棉花植株上多异瓢虫种群数量Table 2 Population number of H.variegata in cotton plants without G.uralensis strips and cotton plants away from G.uralensis strips at different distances during peak periods

结果显示 (表3),6 月17 日、6 月24 日和7月22 日在距离甘草带1 m、5 m、10 m、20 m 的棉田中多异瓢虫与蚜虫的益害比均高于对照棉田。另外,7 月1 日和7 月15 日在距离甘草带1 m、5 m、10 m 的棉田中多异瓢虫与蚜虫的益害比均高于对照棉田。7 月29 日只有在距离甘草带1 m的棉田中多异瓢虫与蚜虫的益害比高于对照棉田。
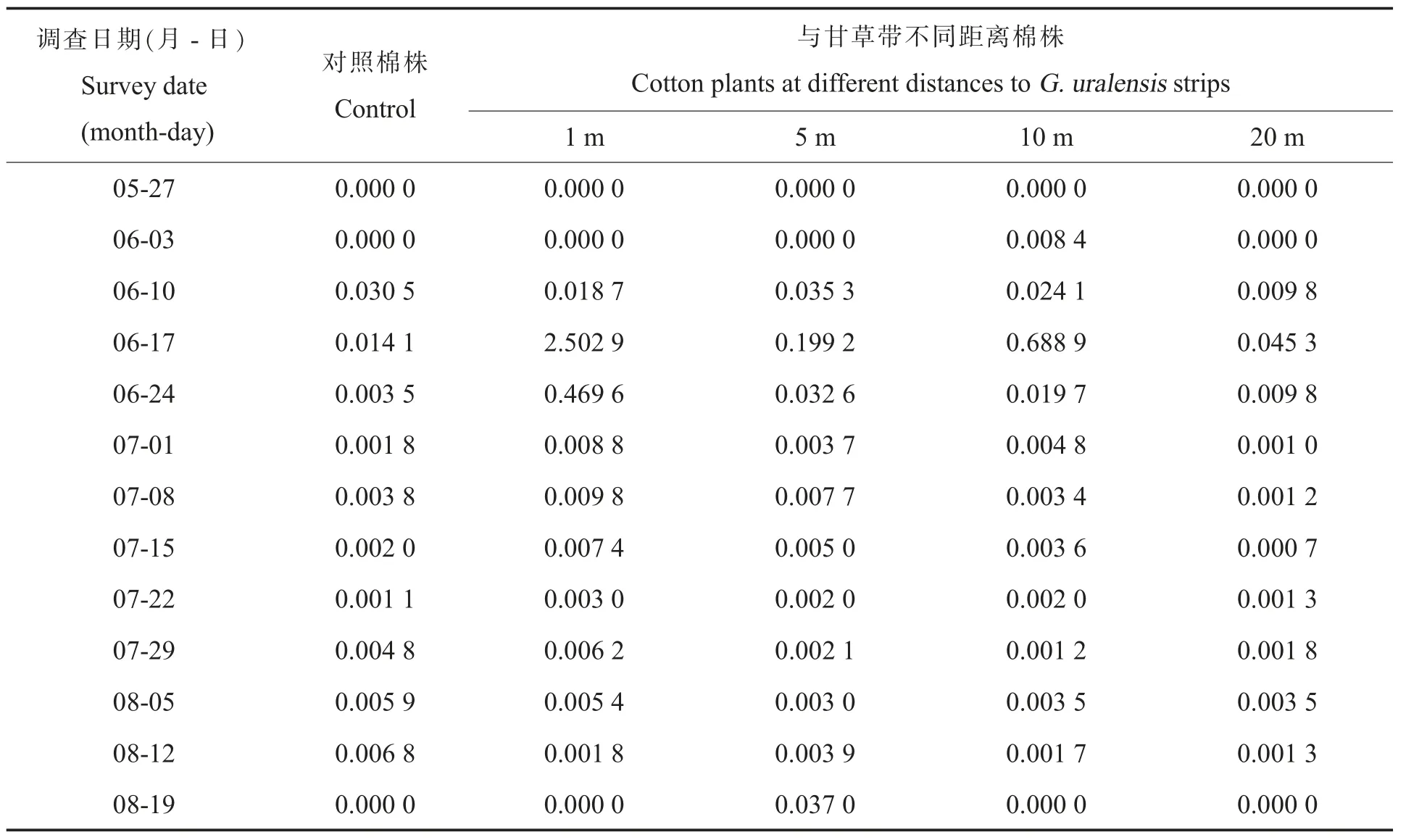
表3 对照棉株与距甘草带不同距离棉株上多异瓢虫与蚜虫的益害比Table 3 Ratios of H.variegata to aphids in cotton plants without G.uralensis strips and cotton plants away from G.uralensis strips at different distances
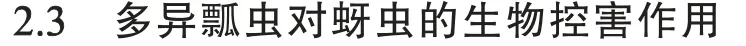
棉田中蚜虫的种群相对增长倍数统计分析结果(图3)显示,罩笼后7 d 棉株上蚜虫种群相对增长倍数为9.09±5.08,极显著(t=4.166,df=18,P<0.001)高于同一调查时间非罩笼棉株上蚜虫相对增长倍数(2.17±1.32);罩笼后14 d,罩笼内棉株上蚜虫种群相对增长倍数达19.20±11.36,极显著(t=4.168,df=18,P=0.002)高于同一调查时间非罩笼植株上蚜虫种群相对增长倍数(4.06±1.67)。
图3 罩笼和非罩笼处理后7 d 和14 d 无甘草带棉株上蚜虫种群增长倍数Fig.3 Relative growth folds of aphids on cotton plants in fields without G.uralensis strips under caged and non-caged treatments for 7 and 14 days
由图4 可知,在有甘草带的棉田中,罩笼后7 d 多异瓢虫对棉株上蚜虫的控害指数为0.72±0.28,而无甘草带棉田中为0.50±0.19,两者之间差异不显著(t=2.039,df=18,P=0.0564)。罩笼后14 d,有甘草带棉田中多异瓢虫对蚜虫的控害指数为0.76±0.21,而无甘草带棉田的控害指数为0.51±0.18,两者之间具有极显著差异 (t=2.882,df=18,P=0.01)。
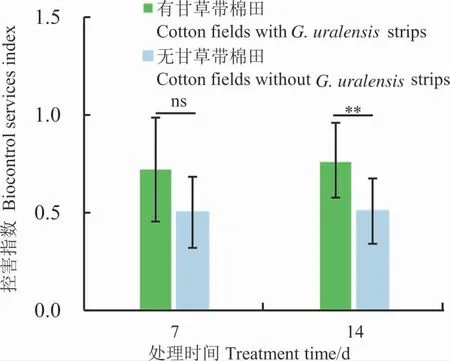
图4 有无甘草带棉株上多异瓢虫对蚜虫的控害指数Fig.4 Biocontrol services index of H.variegata to aphids in cotton plants with or without G.uralensis strips

前人研究表明碱蓬(Suaeda glauca)带[20]、苦豆子(Sophora alopecuroides)带[23]、苜蓿(Medicago sativa)带[24-25]或其他杂草带[13,26-29]能够将天敌昆虫引入棉田,对棉田内多异瓢虫及其他捕食性天敌的发生具有重要的调控作用。
甘草出苗较早[30],5 月下旬至6 月中旬已经进入初花期。由本研究的种群动态调查结果可知,此时甘草上棉黑蚜种群数量较大,为多异瓢虫提供了丰富的猎物资源,多异瓢虫在此阶段主要聚集在甘草带上,建立并维持了一定的种群数量。6 月中旬后,由于温度升高较快,甘草带上棉黑蚜种群数量下降,而此时棉田内蚜虫种群数量逐渐上升,田埂甘草上数量相对较多的多异瓢虫在此阶段迁移至邻近棉田,可实现对棉田内蚜虫的生物防控;7 月上旬,温度持续升高,棉田内蚜虫数量也随之减少,多异瓢虫迁移至其他作物上,在棉田种群数量逐渐下降。与其他植物相比,选用甘草作为新疆棉田储蓄植物具有以下特殊优点:(1)甘草是新疆棉田周边土著物种[31-32],已与棉田形成相对稳定的生态系统[33];(2)甘草能够很好地适应新疆干旱、低温等特殊的气候环境[34-35];(3)甘草生长快、耐盐碱、适应性强、无需严格选择播种地[36],且无需特殊照料、可循环播种等优点有利于进一步节省生产成本[2]。但是另一方面,甘草无法在棉花整个生长发育期间维持多异瓢虫的种群数量,需要选择其他方式或其他储蓄植物进行害虫的持续防控。例如:可以在甘草无法行使储蓄植物功能的时期,选择天敌友好型化学药剂于对天敌生活影响较小的时间进行化学防治[7,18];也可以根据气候变化和植物生长阶段,选用一种或多种储蓄植物[37],继续保育多异瓢虫和其他种类天敌昆虫,以维持天敌昆虫对蚜虫的持续防控[7,38]。
同时,本团队发现无论是在甘草带上还是在棉田内,相较于蚜虫,多异瓢虫的发生具有一定的滞后性[27],即多异瓢虫在一定程度上跟随蚜虫发生[29,37]。这与新疆棉田多种天敌种群的发生动态一致[13,20,23,25],这种天敌对猎物种群动态的跟随性和滞后性可能是由植物在受到植食性昆虫危害后释放出的多种挥发性物质介导的[39]。这些挥发性物质不仅是植物进行间接防御的媒介,也是天敌昆虫定位猎物[40]和转移寄主的直接线索[39,41]。为了更深层次利用储蓄植物——甘草,后续须明确棉黑蚜危害后甘草植株释放挥发性物质的种类和功能,加深对多异瓢虫定位猎物线索的认识,为这些挥发性物质的田间应用奠定理论基础。此外,可将其中可以吸引多异瓢虫的挥发性物质进行合理配比,研发多异瓢虫引诱剂应用于田间,人为实现棉田内多异瓢虫和蚜虫种群发生时间和空间上的重合,达到对棉田内蚜虫种群进行有效控制的目的[42-43]。
本研究发现,距棉田不同距离的甘草带均可在一定程度上增加益害比,提高棉田中多异瓢虫种群数量及对蚜虫的防控效果。其中,距甘草带5 m 处棉田的平均益害比为0.022 5(表3),这与郭佩佩等[44]的研究结果相接近,即当益害比为0.025 0 (1∶40)时,可以兼顾多异瓢虫对豆蚜(Aphis craccivora)防控的时效性和持续性。但是,2 项研究中涉及的蚜虫种类不同,多异瓢虫对2种蚜虫的捕食量也不同[45-47]。因此,田埂甘草带在棉田能发挥控蚜作用的最远距离有待进一步研究。此外,6 月17 日在距离甘草带1 m 处棉田益害比最大为2.502 9,这进一步说明甘草带能够有效保育多异瓢虫,是多异瓢虫后期转移到棉田进行蚜虫生物防治的重要源库。以上2 点提示,储蓄植物系统布局(包括:密度、面积、距离)、稳定性[5,48-49]及刈割时机等,除需室内理论数据支持外,还需要田间应用实践来检验确定。同时,也要考虑到储蓄植物的安全性及天敌昆虫对于替代猎物的偏好性和适应性[43,50],避免储蓄植物本身及其病虫害对目标作物和捕食性天敌产生二次伤害,影响生物防控效果[4]。
罩笼试验是评价天敌生物控害最为简单、直观且最适用于田间的一种方法[51]。本研究罩笼试验结果表明,多异瓢虫能够有效抑制棉田内蚜虫的种群增长(图3),并且棉田田埂甘草带能够在一定程度上提高多异瓢虫对蚜虫的控害指数,提高多异瓢虫对蚜虫的防控效果。其中,罩笼后7 d时,有无甘草带棉田多异瓢虫对蚜虫控害指数差异较大,罩笼后14 d 时,罩笼处理的控害指数显著大于非罩笼处理(图4)。这说明甘草带对于棉田内多异瓢虫种群的保育作用具有持效性,可在较长时间起到保育作用,但这种作用起效较慢,须提前将多异瓢虫释放或“推”进棉田,才能保证将蚜虫种群数量控制在经济阈值以下[42]。但是,对于甘草保育多异瓢虫的持续时间、多异瓢虫进入棉田的最佳蚜虫密度均须根据不同年份、不同气候环境及其他田间实际情况等进一步深入探究。

本研究通过田间系统调查及罩笼评估研究发现,田埂甘草带能够有效地保育多异瓢虫,增加邻近棉田多异瓢虫种群数量,并提高益害比,对多异瓢虫有效控制棉田蚜虫具有重要的调控功能,是新疆地区棉田优质的储蓄植物。
