夜间增温对铅危害下小麦物质积累分配与根系生长的影响
2023-04-09寇太记程相涵王俪睿张慧智陈俊红
寇太记,程相涵,贺 娟,王俪睿,张慧智,陈俊红
(1.河南科技大学农学院,河南 洛阳 471023;2.北京市农林科学院农业信息与经济研究所 北京 100097;3.农业部华北都市农业重点实验室,北京 100097)
我国北方冬小麦生产区春冬季和夜间升温明显[1-2],而且部分农业土壤重金属铅(Pb)等超标[3-4],冬小麦生产遭受气候变暖与Pb胁迫等多重压力。绝大多数增温试验发现,气候变暖有利于我国北方冬小麦生产[5-7],并且夜间增温改变了冬小麦对养分的吸收利用状况[7-8]。Pb危害作物生长既受其含量制约[9-10],又与作物抗Pb胁迫的阈值有关[11-12],土壤Pb危害每年均会导致部分小麦中Pb含量超标[13]。寇太记等[11]发现夜间增温改变了小麦对Pb的吸收分配,气候变暖给判断土壤Pb胁迫对小麦安全生产的影响提出了新挑战。根系作为植株吸收养分和水分的重要器官,是植物响应气候变暖与土壤重金属胁迫的调节器[14],它的发育影响植株地上部生长与优质生产。阐明小麦生长与地下根系对增温与土壤Pb污染双重胁迫的响应特征,对于理解冬小麦的抗逆响应机理具有重要意义。现有文献证实,夜间增温影响小麦的根系形态分布[15]和麦田土壤呼吸速率[16]。根系活性是表征作物抗逆响应的重要指标,且密切影响小麦产量形成[5,16],但增温以及增温下Pb胁迫对小麦根系影响的研究还少见报道。因此,本试验借助多年夜间增温平台[11],通过盆栽试验研究土壤Pb胁迫下关键生育期小麦物质积累及其地下分配、根系活性对夜间增温的响应特征,为全球变暖与重金属污染风险加剧背景下认知冬小麦抗逆响应和制定安全生产调控策略提供理论依据。
1 材料与方法
1.1 研究区与增温平台系统概况
研究区属于暖温带半干旱半湿润季风气候,年均气温13.7℃,降水量650.2 mm左右,以雨养冬小麦-夏玉米连作农业为主。田间增温实验系统位于河南科技大学周山校区农场(34°38′N,112°22′E),由增温区(NW)与常温对照区(CK)两部分构成,本研究增温区采用被动式夜间增温模式,即NW于增温时段(19∶00到次日7∶00)在小麦冠层正上方20 cm处覆盖反射膜但四周无遮挡,在冬小麦全生育期(除降水时段停止)增温,其设计与控制参见文献[11]。试验土壤采自我国重要的工业基地洛阳某金属Pb矿冶炼厂附近,属铅污染土壤,选择与冶炼厂距离不同但理化性质和肥力较一致的农田耕层,土壤类型为褐土,供试土壤理化性质为:容重1.12 g·cm-3、pH 7.8、有机质9.1 g·kg-1、全氮1.28 g·kg-1、有效磷2.4 mg·kg-1、速效钾105.5 mg·kg-1、全Pb 36.8~415.7 mg·kg-1。
1.2 试验设计
田间盆栽试验采用裂区设计,增温处理为主处理,分夜间增温环境(NW)与正常环境(CK)2个水平;Pb处理为副处理,设置Pbck(对照,背景值11.4 mg·kg-1)、Pb0(36.8 mg·kg-1)、Pb1(153.4 mg·kg-1)、Pb2(415.7 mg·kg-1)4个浓度,12次重复。参照国家《土壤环境质量标准(GB15618—2018)》,Pbck、Pb0、Pb1均低于土壤Pb污染风险筛选值170 mg·kg-1(pH>7.5),而Pb2介于风险筛选值与管控值(1 000 mg·kg-1,pH>7.5)之间。试验盆钵为市售PVC大桶(高60 cm,直径50 cm),装盆前,拣去土壤中根茬、石块等侵入物,每个Pb浓度土壤均过2 cm孔径筛后装盆,每盆装干土35 kg。在NW和CK区域按南北方向挖取可摆放各48个盆钵(分4列平行摆放)的长方形土坑,将盆栽随机排列埋入土中,以盆钵上沿高出土表2 cm为宜,共96盆。每盆施纯氮2.0 g(约合135 kg·hm-2)、纯磷(P2O5)1.4 g(约合90 kg·hm-2)和纯钾(K2O)1.4 g(约合90 kg·hm-2)无机肥,其中,50%氮肥(尿素)、磷肥(过磷酸钙)、钾肥(氯化钾)一次性溶解于水后浇于各盆中,并向每盆中补水使土壤含水量为70%饱和含水量,剩余50%氮肥在返青定植后溶水施入;浇水2 d后,于2015年10月19日按每盆35株播种冬小麦(中麦175),返青期每盆定植25株,于2016年5月27日收获。在盆栽四周种植保护行10行。分别在苗期(2015年11月19日)、拔节期(2016年3月23日)、抽穗期(2016年4月25日)、蜡熟期(2016年5月21日)共4个时期采集植株样品,每处理随机选取生长均匀一致的3盆进行测定。整个生育期除特别干旱防止植株死亡进行每盆等量水(2 L)灌溉外,基本不进行人工灌水。
从冬小麦播种到收获除降水的19 d外增温累积191 d,全生育期利用温湿度记录仪(Elitech, Rc-4,浙江精创电力科技有限公司)连续监测冠层温度和耕层土壤温度,每10 min记录1次,日均土壤温度见图1,NW和CK的土壤温度变化范围分别为1.0~19.0℃和0.4~17.6℃,全生育期土壤增温幅度为0.6~1.4℃,平均增温1.08℃。
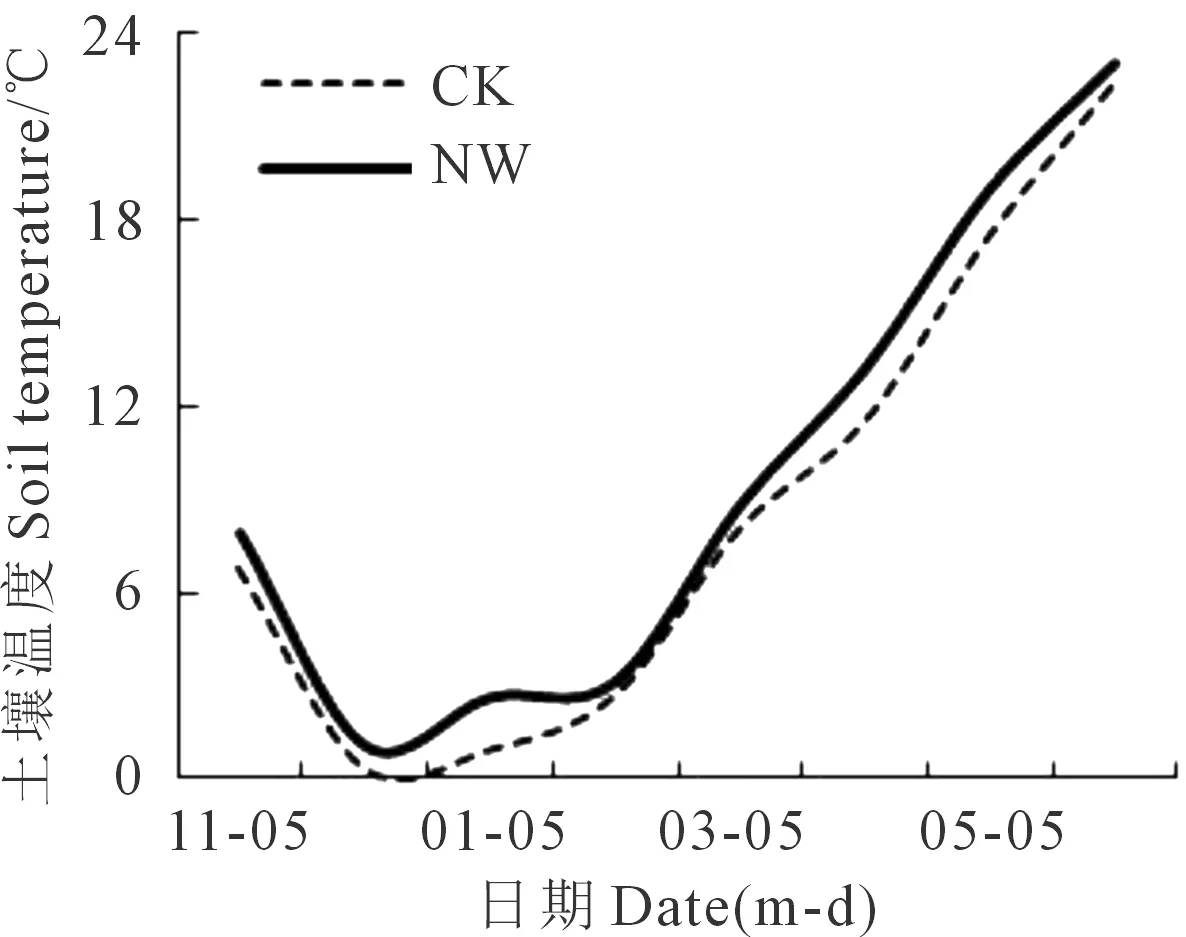
图1 2015—2016年田间被动式夜间增温(NW)和对照(CK)的土壤温度变化Fig.1 Dynamic of soil temperature by passive nighttime warming (NW)and control (CK) in field during 2015 to 2016
1.3 测定内容和方法
2016年5月30日小麦按地上和根系不同组织部位分别进行收获,取样时将整盆从土壤中取出并剪去小麦植株地上部分,将根系从盆中挖出后用水冲洗去残余泥土,收集根系样品并用蒸馏水润洗处理。然后将每株根系样品分成2份,分别测定鲜重,并计算各自占总根鲜重的比例,1份用于测定根系活力,1份烘干用于测定根系干重,并通过后者鲜重及占比折算获得根系总干重;地上部分为茎、叶、颖(包括颖壳、穗轴)、籽粒,分别称量干重;总生物量为地上部与根系生物量之和;收获指数为单株籽粒重与地上部生物量之比;根冠比为根系与地上部生物量之比。根系活力采用改良氯化三苯基四氮唑(TTC)-比色法测定[17]。干重测定则先以105℃的温度杀青30 min,随后以70℃烘48 h后称重获得。
1.4 统计分析方法
数据利用SPSS 16.0统计软件,通过ANOVA对不同温度与Pb污染因素的效应进行方差统计分析,显著水平为P<0.05,利用Excel 2003和Origin 75制图。
2 结果与分析
2.1 夜间增温与铅污染对蜡熟期小麦生物量与籽粒产量的影响
由表1知,Pbck处理下,夜间增温显著增加了蜡熟期小麦籽粒产量8.5%、总生物量12.4%,尽管显著降低了收获指数,但增强了小麦抵抗Pb污染危害的能力。正常与夜间增温环境下,小麦叶重、籽粒产量与总生物量基本上均随Pb含量的增加呈降低趋势,而收获指数则呈升高趋势。在CK环境中,Pb污染显著降低了单株小麦籽粒产量7.0%~26.4%和总生物量6.7%~19.9%,未显著影响收获指数。相比CK环境,夜间增温显著增加Pb污染处理下单株小麦茎重(10.2%~15.0%)、叶重(28.6%~33.3%)、籽粒产量(8.5%~43.2%)、总生物量(12.4%~26.6%)和收获指数(除Pbck处理),增加颖重8.8%~16.2%(仅Pb1和Pb2处理下达显著差异)。表2的因素分析也表明,增温与Pb污染因素均显著影响蜡熟期的叶重、颖重、籽粒产量和总生物量,Pb污染因素显著影响收获指数,二者对颖重、总生物量和收获指数呈显著的交互效应。
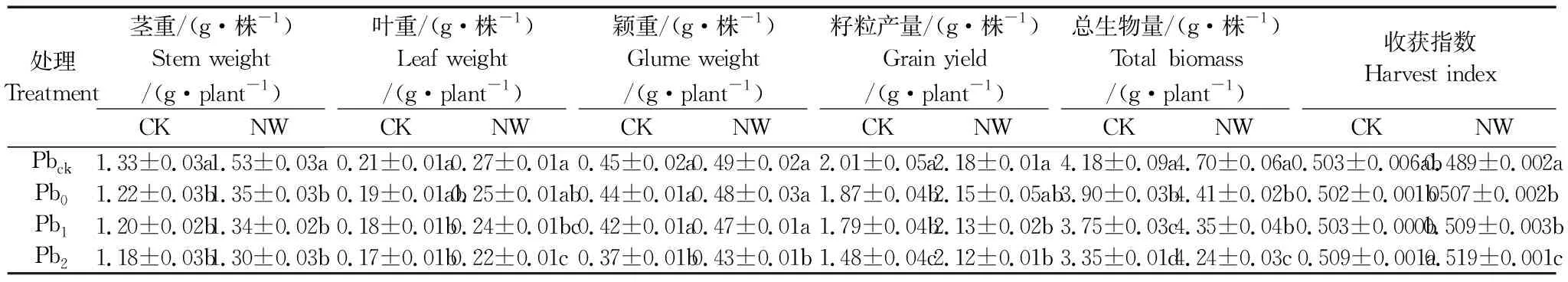
表1 夜间增温和铅污染对蜡熟期小麦总生物量、籽粒产量和收获指数的影响Table 1 Effects of nighttime warming and soil Pb pollution on total biomass, grain and harvest index of wheat in dough stage
2.2 夜间增温与铅污染对小麦根系干重的影响
各处理小麦根干重从苗期到蜡熟期均呈先增后降特征并在抽穗期达到最大值,但其对夜间增温的响应受生育期与土壤Pb污染程度制约(表2,表3)。Pb污染具有抑制根干物质积累的趋势,抑制程度随污染程度增大而加重(表3),在苗期、抽穗期、蜡熟期Pb因素显著影响根干物质积累(表2)。与Pbck相比,生长中后期(抽穗期、蜡熟期)Pb污染下根干重分别显著降低6.8%~44.5%(CK)和21.7%~41.8%(NW)。增温因素显著影响苗期、抽穗期的根干物质积累(表2),与CK环境相比,夜间增温在苗期、抽穗期分别显著增加全部处理的根干重22.2%~60.0%、5.5%~7.7%和蜡熟期Pbck处理的根干重15.8%。双因素分析表明,夜间增温、Pb污染及其二因素的交互效应对拔节期的根干物质积累均无影响,但其交互效应在蜡熟期显著影响根干物质积累(表2)。
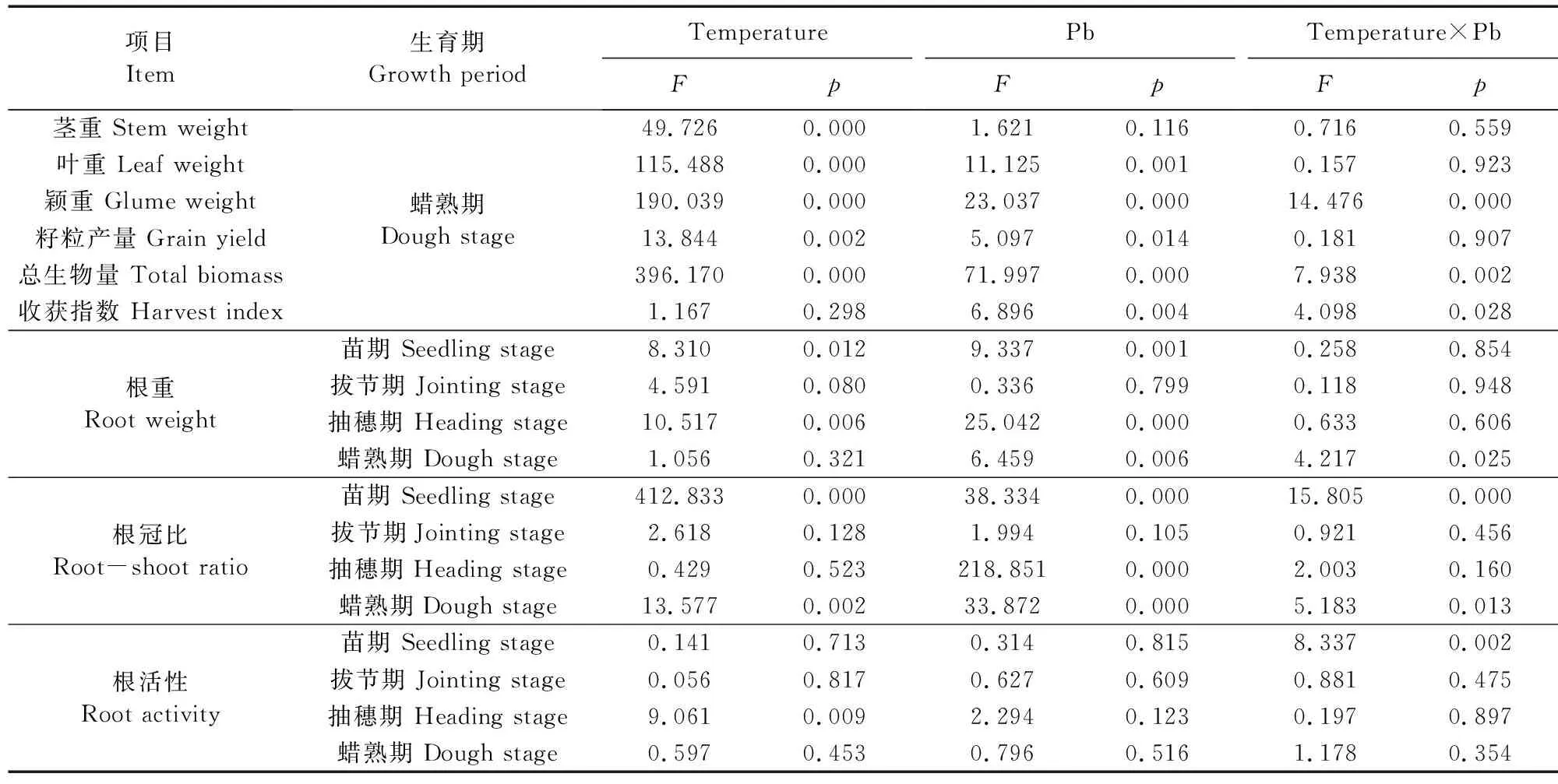
表2 夜间增温与铅污染对小麦生物量分配、根冠比和根活性的影响效应Table 2 Influencing effects of temperature and soil Pb pollution on the accumulation and allocation of biomass, the ratio of root to shoot and root activity of wheat
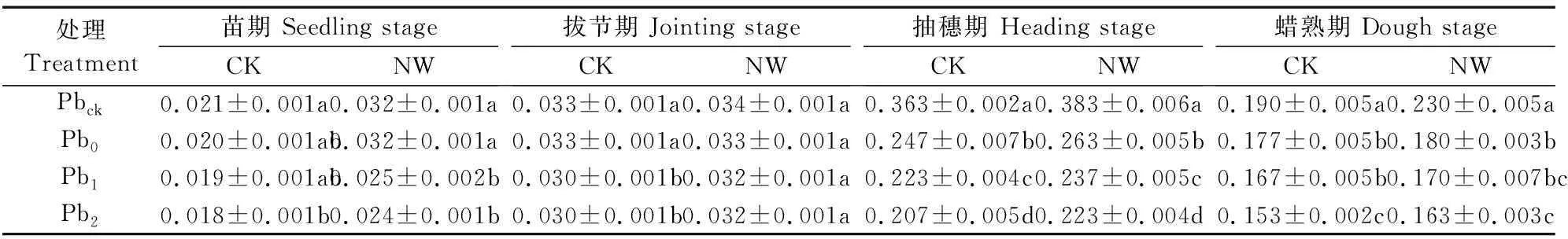
表3 夜间增温和铅污染对小麦根系干重的影响/(g·株-1)Table 3 Effects of nighttime warming and soil Pb pollution on root dry matter accumulation/(g·plant-1)
2.3 夜间增温与铅污染对小麦根冠比的影响
小麦根冠比(RSR)是反映根系与地上部干物质积累是否协调的重要指标。随生育期的推进,各处理的RSR在4个生育期基本呈现大-小-次大-最小的变化规律(表4)。Pb污染降低了苗期、抽穗期和蜡熟期小麦的RSR(蜡熟期CK环境下Pb0处理除外),尽管Pb1和Pb2处理的RSR在NW和CK环境下4个时期较Pbck和Pb0处理显著降低,但Pb1和Pb2处理间的RSR几乎无显著差异;Pb0较Pbck处理的RSR在CK环境中下均无差异,而在NW环境中则显著降低(拔节期除外)(表4)。相比CK,NW显著增加4个时期Pbck处理的RSR 5.6%~20.0%,也显著增加了苗期(5.9%~14.3%)、拔节期(12.5%~25.0%)Pb处理(Pb0,Pb1,Pb2)的RSR和抽穗期的RSR 7.1%~18.2%(Pb2处理除外,P=0.056)。增温与Pb污染在苗期和蜡熟期对RSR的影响呈显著的交互效应(表2)。
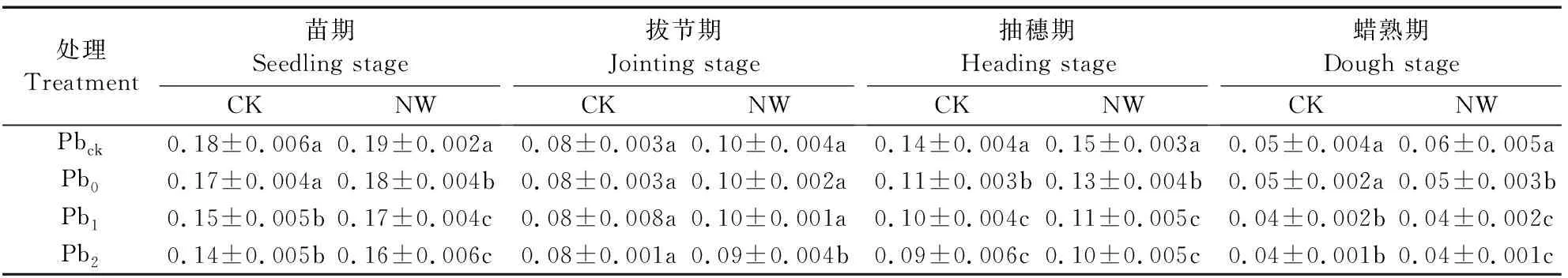
表4 夜间增温和铅污染对小麦根冠比的影响Table 4 Effects of nighttime warming and soil Pb pollution on root-shoot ratio
2.4 夜间增温与铅污染对小麦根系活性的影响
各处理的小麦根系活性均随生育期推进呈现先增后降的特征,在拔节期达到高峰后快速下降,表现为生长前期高于生长中后期,增温在蜡熟期以前具有增加小麦根系活性的趋势(图2)。随着土壤Pb含量增加,CK环境下小麦根系活性在苗期、拔节期呈先增后降变化规律并在Pb0处理达到最大,在抽穗期和蜡熟期则基本呈现逐渐降低态势,Pbck处理下达到最大;而NW环境下小麦根系活性在4个生育期均表现为先增后降的变化特征,均在Pb0处理达到最大。忽略Pb浓度差异,增温显著增加了苗期Pb1处理(8.2%)、拔节期Pb0(5.9%)和Pb2处理(18.1%)、抽穗期各Pb浓度处理(12.2%~40.2%)和显著降低蜡熟期Pbck处理(29.6%)的小麦根系活性。
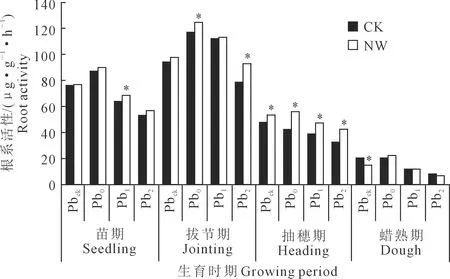
注:* 表示相同处理增温与对照间差异显著(P<0.05)。Note: * indicates differences at the 0.05 level in the same treatment between nighttime warming and no warming treatments.图2 夜间增温与铅污染处理对小麦根系活性的影响Fig.2 Effects of nighttime warming and soil Pb pollution on root activity at different growth stages
双因素分析进一步表明(表2),增温显著影响抽穗期的根系活性,增温与Pb污染因素在苗期对小麦根系活性的影响呈显著的交互效应。
3 讨 论
野外自然条件下的生态系统模拟增温试验可以定量分析和评估气候变暖对作物的影响及其机理[18]。夜间被动式增温模式因其可以模拟非对称增温[1-2],且其更接近于真实环境以及低成本而被国内外广泛应用[19]。本研究所采用的夜间被动式增温在冬小麦生育期达到了较好的增温幅度与效果(图1),且被本平台[11,20]和其他研究[21]的前期增温结果所佐证。由于温度是作物生长发育的关键环境要素之一,本研究发现夜间增温增加了小麦蜡熟期的总生物量与籽粒产量,其他模拟增温试验在成熟期也有类似发现[18]。尽管各研究存在时期差异,但均是由于温度增加促进了小麦生长与干物质的积累所致。然而,夜间增温降低了小麦的收获指数,暗示增温尽管促进了作物光合物质合成与籽粒生产,但降低了光合同化物转化为籽粒的能力,增加的光合产物更多储存在营养器官中。土壤Pb污染加剧不利于蜡熟期的叶重、颖重、籽粒产量和总生物量的提高,这是Pb胁迫对植物生长危害的结果[9]。本研究发现,随Pb含量增加小麦收获指数增加,表明作物在逆境中会优先确保种子生成以保证物种延续,而增温与Pb污染所呈现的对颖重、总生物量和收获指数影响的显著交互效应,则更进一步说明气候变暖将促进小麦碳水化合物合成,但其被更多转移到生殖器官用于确保遗传物质的生产来抵御Pb胁迫危害。
根系已成为植物应对气候变化的重要研究因子[14],是确保植物良好生长与物质形成的重要器官和基本保证[22];本研究即发现较高的小麦总生物量与籽粒产量均对应一个高的根系干物质积累,这与正常或是增温环境无关。夜间增温影响冬小麦根系干物质的积累与分配,其他研究也有类似发现[8,21,23]。尽管有研究认为根干重和次生根数不如根系活性与产量的关系密切[16],但根干重与RSR仍是反映同化产物在植物体内分配及逆境条件下植物自身调节情况的重要指标。本研究发现,RSR在4个生育期基本呈现大-小-次大-最小的变化特征,表明在苗期小麦优先促进根系生长,拔节期地上部快速生长导致RSR变小,抽穗期RSR又变大可能与营养生长转向生殖生长有关,一方面根系生长加速以满足肥水需求,另一方面地上部生长放缓,地下被动增加光合产物贮存,而蜡熟期根系逐渐衰亡[24]导致RSR下降明显。夜间增温下各时期的小麦RSR(表4)和根干重均较大,Liu等[23]认为这是植物响应增温时优先地下分配所致。尽管生育后期小麦根系逐渐衰亡[24],但夜间增温下小麦在中期积累的较大生物量与较高的RSR[16]仍导致了蜡熟期夜间增温下无Pb处理小麦的根干重相对较大。重金属Pb胁迫将对植物造成伤害[9]。本研究中随着Pb含量增加蜡熟期小麦籽粒产量与总生物量降低,由于Pb胁迫会抑制茎根生长,这是地下根系生长随Pb胁迫程度增加受到更大程度的抑制[9-10,25]所致。各时期小麦的RSR与根干重随着Pb污染程度的增加而降低也印证了这一点。夜间增温下Pb污染处理小麦的RSR和根干重(如苗期)显著增加,而且增温与Pb污染在蜡熟期对RSR和根干重的影响呈现显著交互效应,表明气候变暖使植物将更多的光合碳分配到地下促进根系生长以确保植物的抗逆生长,这有利于小麦抵御Pb胁迫。不同生育期小麦根干重积累对夜间增温与Pb的响应有所不同,其他研究者也有类似发现[8,10-11,14-15],这主要是由于小麦的适应差异造成的。
根系活力是植物旺盛生长的基础[26]。本研究中根系活力随生育进程的变化特征与前人研究一致[24]。在小麦生育前期根系旺盛生长,确保了水肥供应与物质快速积累,生长中后期根系活力随着老化进程下降是必然的趋势。本研究发现,土壤中少量Pb(36.82 mg·kg-1)会提高小麦营养生长期的根系活性,Pb含量继续增加则抑制根系活性;而生长后期则随Pb含量的增加小麦根系活性下降,这可能是由于Pb在作物主动或被动吸收其他微量元素过程中产生了竞争[9],促进了前期的根系活性,而生育后期Pb胁迫对小麦根系损害的累积效应与根系自然老化导致根系活性降低。胡智勇等[27]认为温度升高会促进根系活性,但本研究发现增温下根系活性在营养生长阶段增强而在生殖生长末段下降。这种差异应是由于观测阶段的不同和增温使作物早熟导致根系过早老化[8]所致。然而,夜间增温使少量Pb含量下小麦的根系在全生育期均呈现最高的活性,且具有增加Pb污染处理小麦根系活性的趋势,表明夜间增温通过促进作物根系的前期生长与延缓后期老化来适应Pb胁迫影响,并从增温下根干物质积累与RSR相比CK环境处理较高的结果上(表3)得以印证。而增温与Pb污染因素在苗期对小麦根系活性的影响呈显著的交互效应,暗示气候变暖可以通过提升小麦幼苗期的根系活性来抵御Pb胁迫的不利影响,这将增强小麦幼苗在Pb污染下的生存能力。寇太记等[11]前期研究已明确夜间增温能降低铅胁迫农田小麦地上部植株铅积累量,即减少粮食遭受铅污染的潜在风险。夜间增温环境中小麦根系活性增强下根系吸持重金属Pb及减缓Pb向地上部转移的机制,则有待进一步研究。
4 结 论
1)土壤中Pb污染增加会降低小麦生物量与籽粒产量,夜间增温会促进土壤Pb污染下小麦的物质积累与产量形成,提高小麦抗Pb污染危害的能力。
2)土壤Pb污染不利于小麦生物量的地下分配和根系的干物质积累,较低的Pb含量可促进夜间增温小麦环境下全生育期的根系活力,正常环境下抽穗-蜡熟期的根系活力随Pb污染加剧而降低。
3)夜间增温通过提高小麦生育早中期的根系活力、各时期的干物质积累与地下分配量,来抵御Pb污染胁迫对小麦干物质积累的不良影响,但其受Pb污染程度、作物生长时期制约。
