浙江4个主要茶树群体种资源表型性状及遗传多样性分析
2023-03-16郑旭霞黄海涛毛宇骁
丁 一,郑旭霞,黄海涛,毛宇骁,赵 芸
(杭州市农业科学研究院,浙江 杭州 310024)
茶树[Camelliasinensis(L.) O. Kuntze]为山茶科(Theaceae)、山茶属(CamelliaLinn.)、茶组(Section Thea)多年生木本植物,因其异花授粉、种植历史悠久等特性,而具有遗传信息携带量大、群体种资源丰富的特点[1-3]。浙江是中国最大的产茶省份之一,有优越的茶叶种植环境和悠久的种茶历史,其茶树品种资源含大量种植历史悠久的有性群体种材料,其中不乏大量优质或特异资源,鸠坑、木禾、天台群体等为代表的一批名茶优茶群体种享誉世界[4-5]。
在浙江丰富的群体种资源中,天台群体,原产于天台山主峰华顶,天台制茶香气浓郁持久、滋味甘醇爽口、汤色嫩绿明亮、芽叶成朵匀齐,饮后口颊留芳、清心健脾[6];木禾群体,持嫩性强,芽叶生育力较强,制成的珠茶具有花香,为浙江省省级茶树良种[7];鸠坑群体,又名鸠坑大叶种,原产于杭州市淳安县鸠坑乡塘联村,为首批国家级茶树良种之一[8];龙山群体,原产于浙江省上虞市覆厄山,芽叶生育力较强,持嫩性中等,具有抗寒性强、结实性强等特点。
基于茶树自然群体长期的栽培历史及异化杂交的特性,茶树群体种不同于无性系品种,具有丰富的遗传多样性,是茶树新品种选育的良好基础。收集浙江范围内主要种植的这些茶树群体种资源,研究其遗传多样性是未来培育茶树新品种,丰富茶类结构和茶产品的基础。
表型性状鉴定和描述是种质资源研究与利用的有效方法和途径,通过明确不同群体材料表型性状上的特点和多样性,对于定向开发专用型茶树资源是一种很好的手段;但茶树是异花授粉植物,杂合性高,尤其是其中的群体种材料遗传背景较为复杂,单一的表型性状调查不能较为全面地解释茶树群体种的多样性分布,目前各种分子标记已用于茶树种质资源的鉴定筛选和分类研究,并在育种中辅助选配亲本增强遗传多样性以提高育种效率[9-12]。
本研究以天台、木禾、鸠坑及龙山群体为研究对象,对成熟叶、花朵等器官表型性状进行调查和统计分析,同时利用SSR这一重要分子标记进行遗传多样性鉴定,为浙江茶树群体种资源的保存与挖掘和利用提供研究基础。
1 材料与方法
1.1 材料
于2019年春季至2021年春季,分别于种植茶园调查4个群体茶树资源的成叶、花朵性状,供试材料形态特征的描述和生物学特性的调查方法参考《茶树种质资源描述规范和数据标准》,叶相关性状重复观测30次,花相关性状重复观测10次[13]。调查共涉及16个项目,具体调查内容包括萼片数、完全开放花冠大小、花瓣数、最大花瓣质地、花柱长度、柱头裂位、雌雄蕊相对高度、叶长、叶宽、叶正面颜色、叶正面隆起性、叶形、叶身形态、叶基形态、叶缘形态及叶片着生状态,具体调查方法,参照文献[13]进行。群体种鲜叶材料各100株,采集新鲜嫩叶用液氮快速冷却,带回实验室放入-85 ℃冰箱备用。样品采集茶园详见表1。

表1 样品采集茶园地理信息
1.2 表型性状调查与鉴定
表型性状鉴定方法参照《茶树种质资源描述规范和数据标准》[13]。取单株材料,参照每份材料的质量性状重复观察10次,数值性状重复观测5次。详细编码处理见表2。

表2 形态性状的选取及编码
1.3 DNA提取与质量控制
采用柱式法植物基因组DNA提取试剂盒(TaKaRa MiniBEST Plant Genomic DNA Extraction Kit)提取茶树的基因组DNA,用2%琼脂糖电泳和分光光度计检测所提样本的DNA质量和浓度,以保证提取的DNA光吸收值D260/D280=1.80~2.00、样品浓度>30 μg·μL-1、单个样品总量>3μg。
1.4 SSR引物选取及扩增条带特异性分析
随机在已报到的SSR引物中选择平均分布于各染色体的标记引物,对部分样品DNA进行PCR扩增,根据结果挑选多态性高、条带清晰的3对SSR引物作为实验标记引物,详见表3。所有引物由上海百赛生物技术有限公司合成。

表3 筛选所得SSR引物[2]
PCR反应体系参照Reaction Mix 5.0 μL(含DNA Polymerase、2×PCR Buffer、MgCl2和dNTP)、模板 DNA 2.0 μL、Primer-F、Primer-R (10 ng·μL-1)各 0.2 μL,加 ddH2O 至 10 μL。PCR循环参数参照94 ℃预变性 4 min;94 ℃变性30 s,最适退火温度退火30 s,72 ℃延伸30 s,共计35个循环;72 ℃延伸10 min;4 ℃保存(根据各个引物合成信息,退火温度有所调整)。
扩增产物加入2 μL上样缓冲液(6×Loading Buffer)混匀,进行2%的琼脂糖凝胶电泳,采用人工读带的方法,将电泳图上的目的片段范围内清晰的条带,按照分子量大小,从大到小依次记录条带具体大小,单一条带为纯合基因型,两条条带为杂合基因型,建立起原始基因型数据矩阵。
1.5 数据处理
利用Excel 2016和sigmaplot 13软件进行基础数据处理及表型性状变异分析、关联分析及全组分分析;按照供试样本的基因型数据,将其转换成PowerMarker v3.25 软件要求的数据格式,按Neil[14-15]的方法进行遗传距离和遗传相似性分析。
2 结果与分析
2.1 茶树群体种种质资源表型分析
2.1.1 四个群体茶树种质资源表型性状对来自龙山、天台、鸠坑及木禾4个茶树群体的共330份材料的16个主要表型性状分析表明:品种间的差异较大,不同表型性状在不同品种间呈现出不同程度的多样性(表4)。其中与茶花相关的表型性状中,龙山群体萼片数显著高于其他群体,均值为5.20±0.52,且花瓣质地上天台群体显著薄于其他群体(1.90±0.55),接近中厚;鸠坑群体的花冠直径显著低于其他3个群体,均值在(3.29±0.31)cm,且花柱长度上鸠坑群体明显小于其他群体,为(1.09±0.14)cm;天台群体的花瓣数最多(7.70±1.60),其次是龙山群体(7.25±1.16),木禾和鸠坑群体的花瓣数较少;柱头裂位天台>龙山>木禾>鸠坑,依次从中部到上部;雌雄蕊相对高度在4个群体中均无显著性差异,均表现为雌高雄低。

表4 四个茶树群体主要表型性状统计
与叶相关的表型性状中,叶片着生状态在4个群体中均处于上斜至稍上斜状态间,斜度上龙山>木禾>天台>鸠坑群体;天台群体的叶宽显著大于其他3个群体,龙山和木禾群体叶长叶宽均小于其他群体;天台群体叶正面颜色最深,鸠坑群体叶正面颜色最浅,4个群体的叶正面颜色均在深绿到黄绿的范围内;天台群体的叶身形态显著高于其他群体,趋于稍背卷,其余群体都接近于平;鸠坑群体叶形最偏圆形,天台最偏椭圆形,其余两个群体在二者之间;木禾群体的叶基形态显著低于其他群体,更接近楔形;龙山和木禾群体的叶缘形态的波状程度显著低于鸠坑和天台群体。
调查的4个群体中表型性状参数变异系数在4.46%~44.90%,平均为25.28%,其中叶片着生状态变异系数最大,达41.43%,其他变异系数40.00%以上的性状还有柱头裂位(41.31%)。变异系数在30.00%~40.00%和20.00%~30.00%的分别有5项和1项;10.00%~20.00%的有7项,10.00%以下的只有萼片数,变异系数为6.62%,具体见图1。
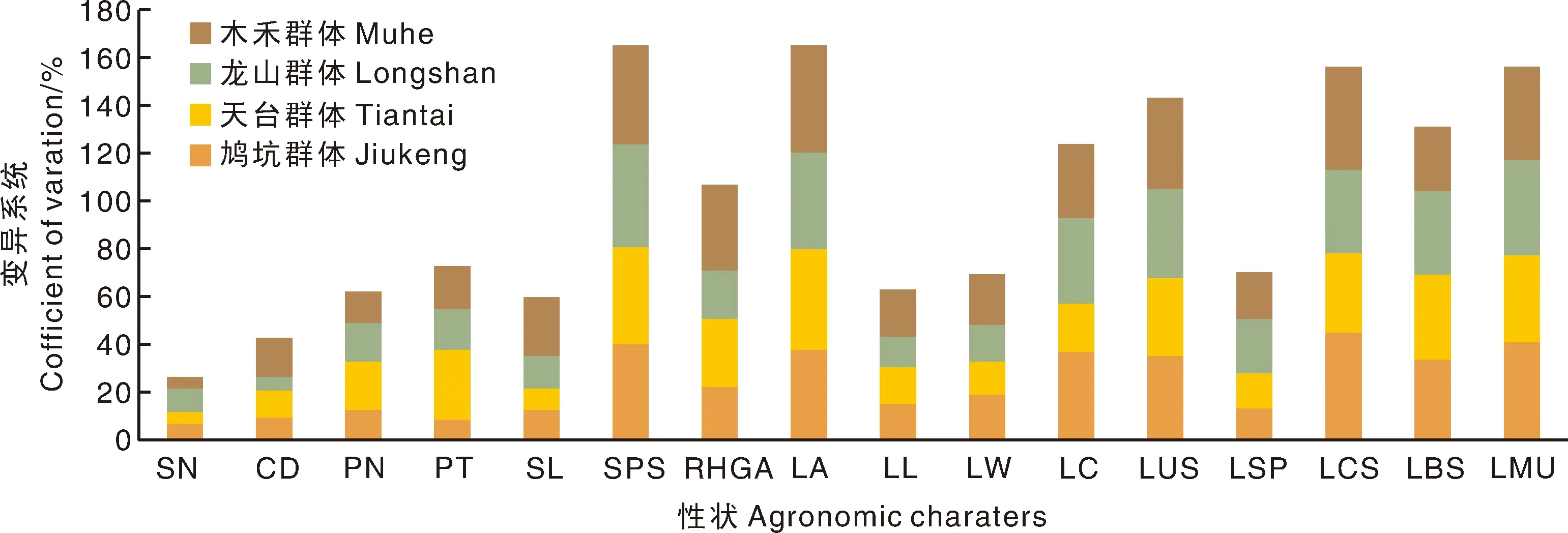
图1 四个茶树群体表型性状变异系数比较Fig.1 Comparison of variation coefficients of agronomic characters among the four tea populations
不同群体的变异系数表现出明显的差异;鸠坑群体变异最大的是叶身形态,天台群体和木禾群体变异系数最大的是叶片着生状态,龙山群体变异系数最大的是柱头裂位;除龙山群体外,其余3个群体变异系数最小的均为萼片数,龙山群体则为花冠大小。
龙山群体较之其他群体遗传多样性更为丰富的表型性状为萼片数、花柱裂位和叶形;鸠坑群体比其他群体更为丰富的性状为叶正面颜色、叶身形态和叶缘形态;天台群体中则为花瓣数、花瓣质地和叶基形态;木禾群体中为花冠直径、花柱长度、雌雄蕊相对高度、叶片着生状态、叶长、叶宽和叶正面隆起性。
2.1.2 四个茶树群体表型性状的相关性分析
单独分析每个群体可以发现,各表型性状的相关性有较大的多样性。对鸠坑群体中各表型性状的相关性分析显示(图2),花柱长度与雌雄蕊相对高度、花瓣质地及花冠直径均呈显著正相关,且花瓣质地及花冠直径呈显著正相关;叶宽和叶长呈显著正相关,叶形与叶身形态和叶宽负相关,叶片着生状态与叶长及叶宽正相关。茶花相关性状及叶相关性状之间相关性较小。

蓝色—龙山群体;黄色—鸠坑群体;绿色—木禾群体;红色—天台群体;橙色—两个群体;紫色—三个群体。Blue-Longshan; Yellow-Jiukeng; Green-Muhe; Red-Tiantai; Orange-two populations; Purple-three populations.图2 四个茶树群体主要表型性状关联分析Fig.2 Analysis of the correlation of the main agronomic characters of the four tea populations
对天台群体中各表型性状的相关性分析显示(图2),花柱长度与雌雄蕊相对高度呈显著正相关,花冠直径与花瓣质地正相关;叶宽和叶长呈显著正相关,叶正面隆起性与叶宽及叶形负相关。较之其他群体,比较特别的是天台群体的花与叶相关表型性状联系更加紧密,而花相关表型性状之间的联系较为薄弱。对龙山群体中各表型性状的相关性分析显示(图2),花相关表型性状相关性较之其他群体较低,其中花瓣数与萼片数负相关;叶相关表型性状中叶宽与叶长、叶身形态与叶正面隆起性正相关,叶基形态与叶长负相关。对木禾群体中各表型性状的相关性分析显示(图2),花柱长度与雌雄蕊相对高度正相关。
单独分析每个群体可以发现,各表型性状的相关性有较大的多样性,尤其是花相关表型性状和叶相关表型性状之间的关联在4个群体中的分布完全不同。因此,针对具体的表型性状可以选取不同的品种组合来丰富特异的性状关联。
对4个茶树群体中各表型性状的相关性分析显示(表5、图3),花柱长度与雌雄蕊相对高度显著正相关;叶长与叶宽、叶身形态及叶片着生状态均显著正相关,而叶宽还与叶正面隆起性、叶身形态、叶基形态及叶片着生状态显著正相关,与叶形负相关;叶形与叶宽、叶正面隆起性、叶身形态及叶基形态显著负相关;总的来说,较之单独分析单个群体可以发现,叶相关表型性状及花相关表型性状的关联度均显著增加,且花叶表型性状之间的联系也更为紧密,其中花柱长度与叶缘形态显著正相关,柱头裂位与叶形显著负相关,叶长与花瓣数显著正相关。

表5 四个茶树群体主要表型性状关联分析
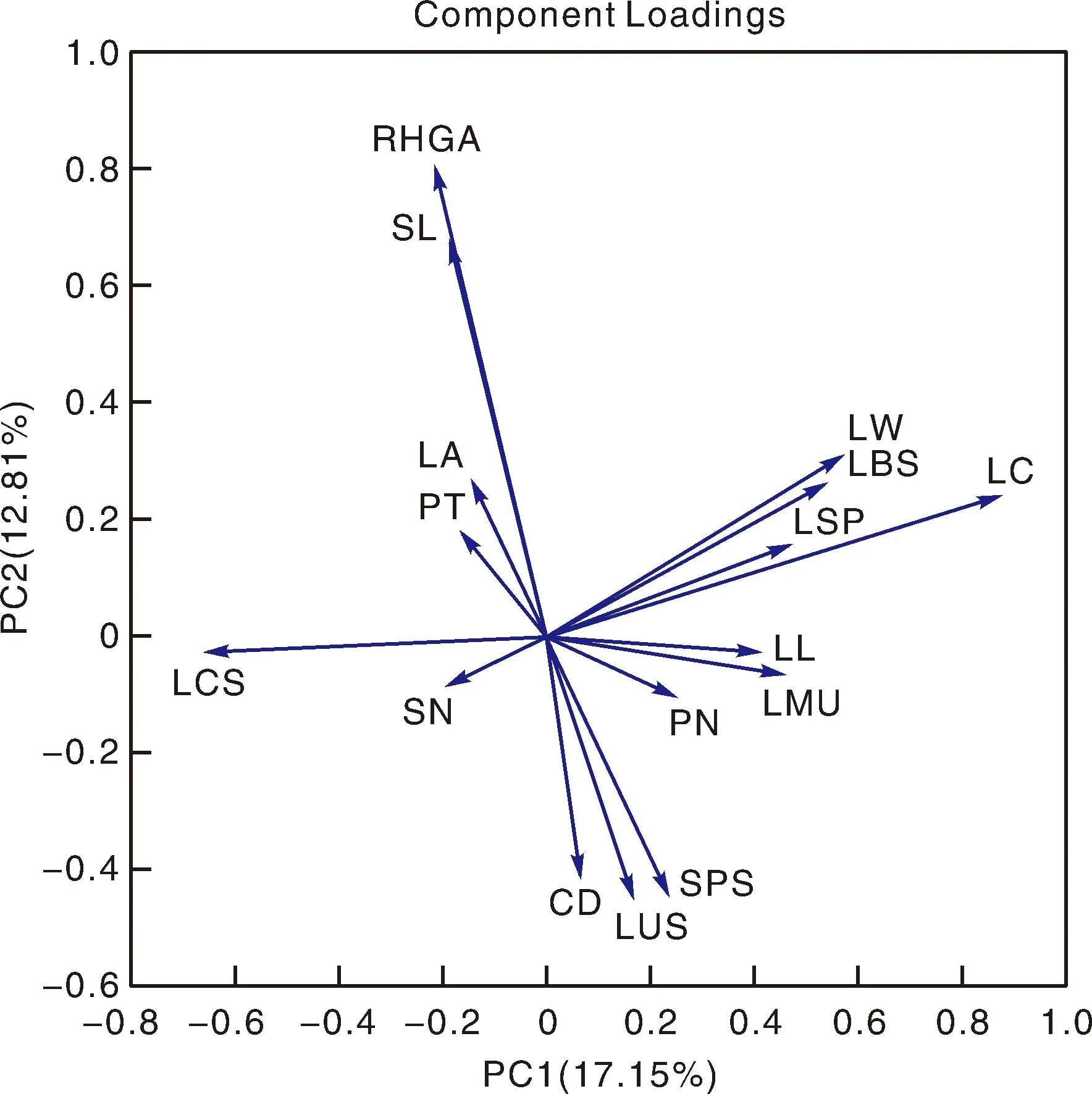
各特征值关联性分析以表示不同参数之间的相关性,相同方向的变量表示正向关系,相反表示负向关系。Correlation analysis of various eigenvalues indicated the correlation between different parameters. Variables in the same direction showed a positive relationship and the opposite showed a negative relationship.图3 四个茶树群体所有表型性状特征数据的主成分分析Fig.3 Principal component analysis of all agronomic characters data of four tea populations
2.2 茶树种质资源基因型变异分析
筛选了在4个群体中具有差异性的3对SSR引物进行群体扩增,结果表明,3对SSR引物均能在供试材料中检测到多态性,其中引物1在天台群体中检测到较强的多态性,而在鸠坑及木禾群体中较为稳定;引物4在天台、龙山及木禾群体中均检测到较强的多态性,引物6在鸠坑和天台群体中检测到较强的多态性;总的来说天台群体展现出的遗传多样性明显高于其他群体(表6)。

表6 四个茶树群体SSR分子标记多态性
3 讨论
丰富优质的遗传多样性种质资源是定向育种的基础,掌握不同亲本的差异性有利于提高育种效率,明确育种方向。本研究中茶树群体种种质资源的主要表型性状差异较大,遗传变异性丰富,但不同性状的遗传稳定性有所差异,龙山群体中相比其他群体遗传多样性更为丰富的表型性状为萼片数、花柱裂位和叶形;鸠坑群体中比之其他群体遗传多样性更为丰富的为叶正面颜色、叶身形态和叶缘形态;天台群体中则为花瓣数、花瓣质地和叶基形态;木禾群体中为花冠直径、花柱长度、雌雄蕊相对高度、叶片着生状态、叶长、叶宽和叶正面隆起性。群体种材料因其丰富的遗传多样性常被作为选种繁育的亲本材料,以上结果在以不同性状为育种目标的材料选择上可以有一定的参考价值。
以茶花为研究对象,表型性状遗传多样性较为丰富的群体为天台群体,以叶为研究对象,鸠坑群体是更为理想的材料,开发花叶两用材料可选择总表型性状遗传丰富度最高的木禾群体。结合该结果,分别开发可用于优化叶性状或花性状的茶树新资源时可选择性地选取不同亲本。
对4个群体的茶树种质资源基因型变异分析结果表明,4个群体都具有分子水平上的遗传多样性,且在大多数引物水平下,天台群体展现出的遗传多样性明显高于其他群体。由于4个群体分别各采样于两个茶园,地理环境有差异,分别对4个群体共8个地点的数据进行全组分分析发现(图4),龙山群体在各特征值下分布较为松散,其他群体都保持有一定的聚合性,说明地理环境对茶树群体种的影响在不同品种间有所差异,龙山群体显著受到地理环境的影响,其他群体则相对稳定。

图4 不同茶园的4个茶树群体材料的相关性分析Fig.4 Correlation analysis of the four tea populations in different gardens
综上所述,茶树群体种资源在长期的自然选择和人工栽培条件下,从分子水平到表型性状都展现出较强的多样性、稳定性和变异性,龙山群体受到地理环境影响较大,作为育种材料稳定性较弱;以茶花为研究对象,表型性状遗传多样性较为丰富的群体为天台群体,且其分子水平上展现出较为丰富的遗传多样性;以叶为研究对象,鸠坑群体是更为理想的材料;花叶兼用则木禾群体是较优的材料。利用不同茶树群体种的表型性状特异丰富度及遗传信息多样性,作为专用研究对象用于浙江茶树群体种资源的保存与挖掘,有望选育出具有优异品质的茶树新品种。
