饲料中添加胆汁酸对欧洲鳗鲡幼鱼肝脏脂肪代谢的影响
2023-03-10马志茹赵盼月翟少伟
马志茹 赵盼月 翟少伟
(集美大学水产学院鳗鲡现代产业技术教育部工程研究中心, 厦门 361021)
胆汁酸(Bile acids, BA)由动物肝脏中合成, 贮存在胆囊中, 摄食后排入肠道中发挥作用; 其具有亲水性的羟基及羧基, 又含有亲油性烷基的特殊的两性分子结构, 决定了其具有较强表面活性, 在促进肠道对饲料中脂类物质的消化吸收中发挥重要作用[1,2], 还可通过激活法尼酯X受体(Farnesoid X receptor, FXR)调节脂类代谢[2,3]。BA除了在脂类消化吸收代谢方面的基本作用外, 还具有提高动物免疫力, 减少细菌内毒素吸收、抑制有害菌增殖及利胆的生理功能[1]。因此, 胆汁酸作为鱼类饲料添加剂的报道逐渐增多, 且大多研究表明适量添加BA可促进鱼类生长, 提高饲料利用效率[4,5]。近来的研究表明, BA可降低投喂高脂饲料的草鱼(Ctenopharyngodon idellus)[6]、大黄鱼(Larimichthy scrocea)[7]、黑鲷(Acanthopagrus schlegelii)[8]、红鳍东方鲀(Takifugu rubripes)[9]和虹鳟(Oncorhynchus mykiss)[10]等鱼类肝脂过度蓄积, 缓解肝脂过量蓄积产生的肝功能损伤; 还能减轻投喂低蛋白质饲料草鱼[11]、高淀粉饲料大口黑鲈(Micropterus salmoides)[12,13]、豆油替代鱼油饲料大黄鱼[14]、高非淀粉多糖饲料虹鳟[15]及高植物性原料比例饲料的大菱鲆(Scophthalmus maximus)、罗非鱼(Oreochromis niloticus)和虹鳟等体内脂肪代谢紊乱[16—18]。上述研究结果显示了BA在维持鱼类肝脏脂类代谢稳态方面具有重要作用, 还鲜见其在鳗鲡(Anguillaspp.)饲料中的报道。
鳗鲡营养丰富, 素有“水中人参”的美誉, 作为我国重要的经济鱼类, 连续多年在出口创汇的单一水产品中排名第一。欧洲鳗鲡(Anguilla anguilla)曾是我国主要鳗鲡养殖品种, 但该品种属于《濒危野生动植物种国际贸易公约》CITES附录Ⅱ的濒危保护物种, 目前国内进口量较少, 仍有一定养殖规模[19]。欧洲鳗鲡全鱼脂肪含量在可养殖的主要鳗鲡品种中最高, 且肝脏功能易受脂肪代谢紊乱带来的负面影响[20,21], 开展BA添加在欧洲鳗鲡饲料中的应用研究具有重要的实践和理论意义。因此, 本实验旨在研究欧洲鳗鲡幼鱼饲料中添加不同水平BA对肝脏脂肪代谢的影响, 为揭示BA调节鱼类脂肪代谢机制提供参考。
1 材料与方法
1.1 实验设计及实验饲料
将养殖欧洲鳗鲡幼鱼的9口水泥池[初始规格(141.5±1.9) g/尾; 初始鱼重(682±23) kg/池]随机分为3个组, 分别投喂幼鳗商业饲料(对照组)及在其基础上添加500 mg/kgBA(BA1组)、基础饲料添加1000 mg/kg BA(BA2组)的饲料; 每个组3个重复, 养殖实验持续15 周。在本实验中胆汁酸添加水平根据以往胆汁酸在不同鱼类饲料中适宜添加水平及胆汁酸的推荐水平在500 mg/kg左右而综合考虑确定。
所用幼鳗商业饲料由浙江科盛饲料股份有限公司生产且未添加BA, 主要原料为白鱼粉、α-糊化淀粉、酵母粉、膨化大豆及复合预混料, 其主要营养指标(测定值): 水分6.08%、粗蛋白质47.43%、粗脂肪4.81%、灰分16.87%、钙4.16%和总磷2.85%。将BA(猪源胆汁酸产品, 有效含量50%, 厦门兴牧威动物保健品有限公司生产)按照500和1000 mg/kg水平添加在上述商业基础饲料中, 采用逐级混合均匀的方法制作实验组饲料。
1.2 饲养管理
本实验在福建锦江之曼水产科技有限公司5号养殖车间进行, 选用具有循环水设施的六边形水泥池(面积30 m2, 高1.5 m, 有效水体体积27 m3, 水体流速为0.39 m3/min, 采用液氧充气)开展实验研究,所有欧洲鳗鲡幼鱼养殖水泥池实行统一管理。在实验开始前, 每日用搅拌机(YE2132M-6, 上海皓天电机有限公司)将粉状基础饲料与水混合均匀(重量比为1∶1.2), 和成面团状饲料, 再切成2 kg左右的小块进行投喂。实验鱼日投喂2次(每天06:00和18:00), 每餐根据摄食活力、水质变化和天气变化等状况确定具体投喂量; 采用自然光照, 另在投喂时开日光灯1h。由于在实际生产中规格相近的鳗鲡经过一段时间养殖, 鱼体重因个体生长速度的不同而出现较大差异, 所以一般每隔2—3个月进行一次规格大小的分群操作, 即将规格接近的鳗鲡挑选出来再置于同一池中养殖。在公司5号养殖车间的欧洲鳗鲡幼鱼重新分群后, 挑选出规格和总重接近的9口水泥池用于本实验。正式实验开始后每日记录摄食和死鱼状况(实验期间鳗鲡成活率平均为99.8%); 实验期内控制水温26—30℃, 溶解氧10—15 mg/L, pH 6.0—6.5, 氨氮浓度8.0—8.5 mg/L,亚硝酸盐浓度0.15—0.22 mg/L; 鳗鲡其他饲养管理、水质指标等与同养殖车间内其他养鳗池保持一致。
1.3 样品采集与处理
在实验结束后, 实验鱼禁食24h, 每口池随机捞取12尾鱼, 被无水乙醇配制成2 g/L丁香酚麻醉后再无菌解剖鱼体后分离肝脏取样, 其中3尾鱼肝脏同一部位的肝小叶重要静脉周围取样, 再置于Bouin氏液中至少固定24h后再制作切片; 剩余肝脏样品按照赵盼月[21]的方法采集及预处理样品待测脂肪代谢酶水平或活性。6尾鱼肝脏样品冷冻保存待测常规营养成分。3尾鱼肝脏样品放到无菌冻存管中液氮速冻, -80℃保存后将对照组和BA1组各6个肝脏样品送至上海敏心生物科技有限公司进行脂质组学分析。在本课题组之前的研究中, BA1组与BA2组欧洲鳗鲡幼鱼生长、肠道消化酶活性和血清生化主要指标等接近[5], 两组肝脏脂肪代谢变化可能相似, 只将BA1组与对照组进行脂质组学分析。
1.4 测定指标
肝脏组织切片制备及观察肝脏组织样品在4%甲醛固定液中固定24h后于75%的酒精中浸泡, 更换数次以去除样品中苦味酸的黄色, 按照常规方法进行脱水、透明、包埋、切片及HE染色,石蜡封片, 在100倍观察肝脏组织形态和结构[21]。
肝脏常规营养成分肝脏常规营养成分测定使用常规方法, 其中采用凯氏定氮法测定粗蛋白水平; 采用乙醚索氏抽提法测定粗脂肪水平; 采用常规马弗炉高温灼烧法测定灰分水平。
肝脏脂肪代谢酶采用南京建成生物工程研究所生产的试剂盒测定肝脏脂肪酸合成酶(Fatty acid synthetase, FAS)和乙酰辅酶A羧化酶(Acetyl CoA carboxylase, ACC)水平, 肝脂酶(Hepatic lipase,HL)和脂蛋白酯酶(Lipoprotein lipase, LPL)活性。具体操作步骤详见说明书。此外, 总脂酶(Total lipase, TL)活性为HL和LPL两种酶活性之和。
肝脏脂质组学分析肝脏样本于冰上缓慢解冻后取50 mg样本, 加入1.5 mL氯仿/甲醇(2/1,v/v)溶液, 加入0.5 mL纯水, 10 μL内标[200 μg/mL,PC(11∶0/11∶0)], 涡旋1min; 3000 r/min离心10min,取尽有机相于干净的试管中, 氮气吹干。用400 μL异丙醇/甲醇(1/1,v/v)复溶; 12000 r/min, 4℃离心10min后取上清于进样瓶待进行脂质组学分析。脂质组学分析委托上海敏心生物科技有限公司采用LC- MS (Thermo, Ultimate 3000LC, Q Exactive) 仪器平台进行, 所用色谱柱及其分离条件、质谱检测参数、扫描模式、碰撞模式、数据处理及统计分析方法等均与Wang等[22]相同。
1.5 数据统计分析
欧洲鳗鲡幼鱼体成分及肝脏脂肪代谢酶等指标采用SPSS 22.0统计软件中One-way ANOVA法进行单因素方差分析。P<0.05表示差异显著。脂质组学分析使用Lipid Search软件(Thermo Fisher Scientific, USA)、SIMCA-P 13.0 (Umetrics AB, Umea,Sweden)软件及OPLS-DA模型鉴定差异显著的代谢物(VIP>1,t检验P<0.05), 用MetaboAnalyst4.0数据库寻找差异脂质代谢产物富集的代谢通路。
2 结果
2.1 欧洲鳗鲡幼鱼肝脏组织形态
如图 1所示, 不同组欧洲鳗鲡幼鱼肝脏组织中脂肪空泡数量方面存在较大差异; 对照组肝脏中脂肪空泡较多, BA组肝脏中脂肪空泡数量均较少。

图1 饲料中添加胆汁酸对欧洲鳗鲡幼鱼肝脏组织结构的影响Fig. 1 Effects of dietary bile acid supplementation onliver histology of European eel juveniles
2.2 欧洲鳗鲡幼鱼肝脏常规营养成分
由表 1可知, 与对照组相比, 添加BA显著增加了欧洲鳗鲡幼鱼肝脏粗蛋白水平(P<0.05), 显著降低粗脂肪水平(P<0.05), BA组间粗脂肪和粗蛋白水平无显著差异(P>0.05); 不同组间灰分水平接近(P>0.05)。

表1 饲料中添加胆汁酸对欧洲鳗鲡幼鱼肝脏常规营养成分的影响Tab. 1 Effects of dietary bile acid supplementation on proximate composition in liver of European eel juveniles (wet sample; %)
2.3 欧洲鳗鲡幼鱼肝脏脂肪代谢酶
由表 2可知, 与对照组相比, BA组肝脏FAS水平显著降低(P<0.05), 仅BA1组ACC水平显著降低(P<0.05), LPL、HL和TL活性均显著升高(P<0.05);BA1组和BA2组间均无显著差异(P>0.05), BA2组ACC水平与对照组无显著差异(P>0.05)。

表2 饲料中添加胆汁酸对欧洲鳗鲡幼鱼肝脏脂肪代谢酶活性或水平的影响Tab. 2 Effects of dietary bile acid supplementation on lipid metabolism enzymes activities or levels in liver of European eel juveniles
2.4 欧洲鳗鲡幼鱼肝脏脂质组学分析
正交偏最小二乘法(OPLS-DA)判别分析融合了正交信号校正的干扰信息剔除能力及偏最小二乘判别的相关信息提取能力, 是脂质代谢组学中最常用的建模方法。本实验OPLS-DA法建模得分图见图 2。可见, 本模型具有较好的稳定性和预测性,不存在过拟合现象; 对照组和BA1组代谢组产物有较好分离效果, 两组之间存在明显差异。
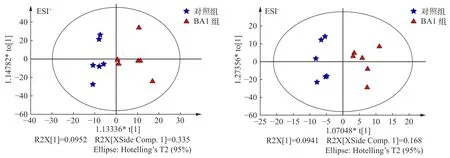
图2 正、负离子模式下对照组和BA1组OPLS-DA分析得分图Fig. 2 The OPLS-DA scores plot of control group and BA 1 group under ESI+ and ESI- mode
如图 3所示, 与对照组相比, BA1组欧洲鳗鲡幼鱼肝脏中差异脂质代谢产物富集的代谢通路按照重要程度由大到小分别是甘油磷脂代谢、甘油酯代谢、亚油酸代谢、α-亚油酸代谢及花生四烯酸代谢。

图3 BA1组与对照组相比欧洲鳗鲡幼鱼肝脏差异脂质代谢途径富集通路气泡图Fig. 3 The scatter plot of the enriched metabolic pathway for all matching significant metabolites in liver of European eel juveniles of BA1 group in comparison with control group.
由表 3可知, 两组欧洲鳗鲡幼鱼肝脏中存在显著差异的主要是甘油磷脂代谢和甘油酯代谢。从所匹配的脂质产物变化情况可以看出BA1组这两条通路的代谢增强, 涉及的脂质产物主要是磷脂酰胆碱和溶血磷脂酰胆碱。此外, 磷脂酰胆碱参与的亚油酸代谢、α-亚麻酸代谢、花生四烯酸代谢在两组中也存在一定差异, 代谢强度也有增加的趋势。
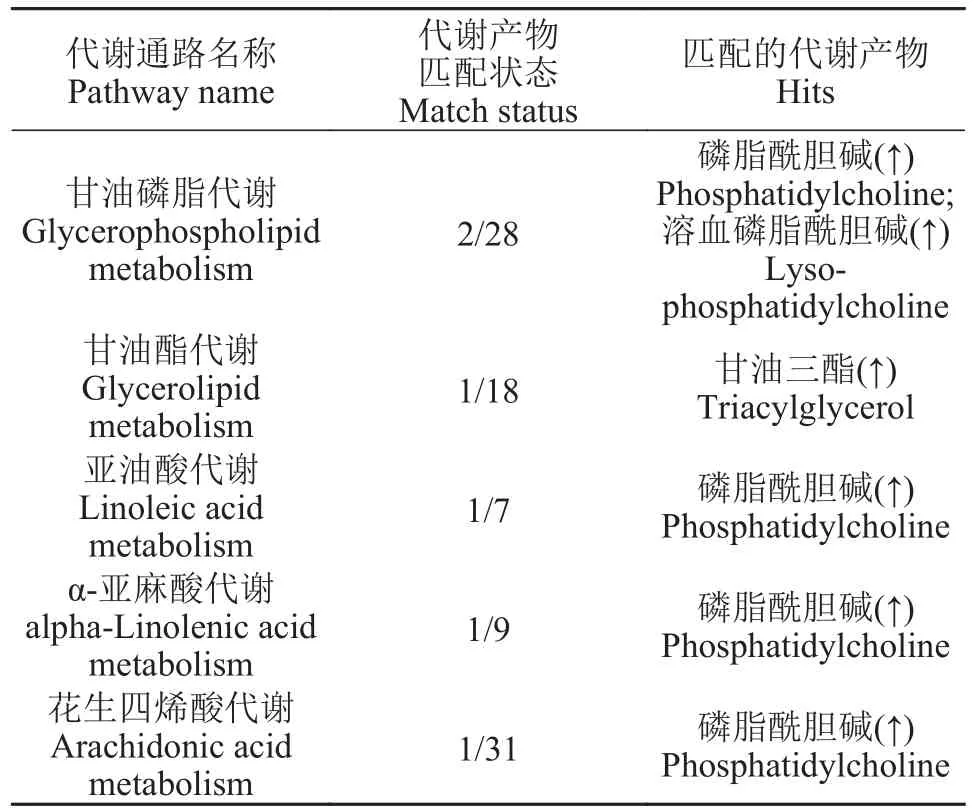
表3 BA1组与对照组相比欧洲鳗鲡幼鱼肝脏中差异代谢通路匹配的脂质代谢产物Tab. 3 The most relevant metabolic pathways and corresponding differential lipid metabolites in the liver of European eel juveniles of the BA1 group in comparison with the control group
3 讨论
3.1 肝脏脂肪蓄积
研究表明, BA除了参与食物中脂类和脂溶性维生素溶解、消化和吸收之外, 还可通过激活FXR调节肝脏脂质生成和分泌、血浆脂质清除及肠道胆固醇吸收来影响脂质代谢及自身代谢[2,3]。
在我们以往研究中发现, 在饲料中添加500 mg/kg BA可降低欧洲鳗鲡血清中甘油三酯、总胆固醇及低密度脂蛋白胆固醇水平[5]。在本实验中, BA组欧洲鳗鲡幼鱼肝脏切片的结果显示脂肪空泡数量减少, 这与在黑鲷[8]、大口黑鲈[12,13]、罗非鱼[17]、乌鳢(Channa argus)[23]、杂交石斑鱼(Epinephelus fuscoguttatus♀× E. lanceolatus♂)[24]、鳜(Siniperca chuatsi)[25]和鲤(Cyprinus carpioL.)[26]饲料中添加BA后肝脏组织切片中脂肪空泡变化的结果一致;BA组欧洲鳗鲡幼鱼肝脏脂肪水平显著降低, BA这种降低肝脏脂肪水平的效应在大黄鱼[7]、红鳍东方鲀[9]、大口黑鲈[12]、罗非鱼[17]、杂交石斑鱼[24]、鳜[25]、鲤[26]、草鱼[27,28]、大菱鲆[29]和军曹鱼(Rachycentron canadum)[30]也有类似的报道, 说明添加BA具有明显降低鱼类肝脏脂肪蓄积的作用。研究表明, 添加BA未显著提高罗非鱼[17]、鳜[25]、草鱼[27,28]和军曹鱼[30]肝脏中粗蛋白质水平, 这与本研究结果有所差异, 但在低脂肪水平下添加BA可显著提高大菱鲆肝脏脂肪水平[29], 具体原因有待查明。
3.2 肝脏脂肪代谢酶活性或水平
肝脏是鱼体脂质代谢最为活跃器官, 其相关代谢酶的变化可以反映出体内脂肪代谢状况。FAS是脂肪酸合成的关键酶之一, 催化乙酰辅酶A和丙二酸单酰辅酶A转变成脂肪酸, 可被胆固醇调节元件结合蛋白(Sterol regulatory element-binding protein, SREBP-1)调控长链脂肪酸的合成[7]。ACC是脂肪酸合成限速酶, 可催化乙酰辅酶A形成丙二酰辅酶A, 控制体内脂肪合成代谢的强弱[31]。在本实验中, 饲料中添加BA后, 欧洲鳗鲡幼鱼肝脏中的FAS和ACC水平显著降低, 表明脂肪的合成代谢受到抑制。LPL和HL是鱼类肝脏脂肪分解代谢过程中的两个关键酶, TL为二者之和。LPL可脂解血浆脂蛋白-甘油三酯而释放游离脂肪酸和单酸甘油酯,以供组织氧化供能和贮存, 间接决定从饲料中的脂类以体脂形式贮备起来或作为能源底物消耗掉[14,23,32]。HL存在于肝内皮细胞表面, 可促进低密度脂蛋白胆固醇和乳糜微粒残粒进入肝细胞, 参与高密度脂蛋白胆固醇逆转运及其残粒分解[22,23,32]。在本研究中, 欧洲鳗鲡幼鱼饲料中添加BA后, LPL、HL和TL活性均显著上升, 说明肝脏脂肪的分解代谢增强。因此, 欧洲鳗鲡幼鱼饲料中添加BA可通过抑制肝脏脂肪合成代谢, 促进脂肪分解代谢来调节脂质代谢, 从而减少脂肪蓄积。此外, 与陆生动物的研究结果类似, 在鱼类中的研究也证实FXR的激活是BA调节脂质生成的关键途径。在生理条件下,参与脂肪生成的ACC酶和脂肪酸合成的FAS酶均受小异源二聚体伴侣受体(Small heterodimer partner, SHP)调控[33], 推测BA可通过激活鱼类肝脏中FXR可通过介导 SHP来调控元件结合转录因子-1C(SREBP-1C)的表达[9,14,34]; 还可能诱导过氧化物酶体增殖物激活受体α高表达, 上调脂肪酸β氧化相关酶, 促进脂肪酸氧化[7,23,24], 从而减少脂质沉积。
3.3 肝脏脂质代谢产物
在本实验中, BA1组欧洲鳗鲡幼鱼肝脏中差异脂质代谢产物富集的代谢通路主要是甘油磷脂代谢、甘油酯代谢、亚油酸代谢、α-亚油酸代谢及花生四烯酸代谢等, 涉及的主要差异脂质代谢产物是磷脂酰胆碱和甘油三酯, 且呈上调趋势。磷脂酰胆碱是鱼体内主要的甘油磷脂, 也是脂蛋白的主要成分[35,36]; 与蛋白质、维生素并列的“第三营养素”,能够保持肝脏正常功能, 保护肝脏, 预防脂肪蓄积[37]。磷脂酰胆碱在磷脂酶A2的作用下水解为溶血磷脂酰胆碱, 同其他脂肪降解产物一起与胆汁盐混合,最终被肠粘膜细胞吸收[38]。溶血磷脂酰胆碱还可作为磷脂酰胆碱的瞬时中间体, 发挥第二信使作用[39]。此外, 甘油磷脂代谢相关的差异代谢产物, 还是胆汁的重要成分之一[40]。本实验肝脏靶向脂质代谢组学结果显示, BA1组欧洲鳗鲡幼鱼肝脏磷脂酰胆碱和溶血磷脂酰胆碱水平上调, 表明添加BA可增强使肝脏甘油磷脂代谢。甘油磷脂主要包括磷脂酰胆碱、磷脂酰乙醇氨和磷脂酰肌醇等, 是细胞膜主要的脂质成分, 在细胞增殖、分化和凋亡中发挥关键作用[35,41]。研究表明, 添加外源胆汁酸未降低红鳍东方鲀胆囊中大部分种类胆汁酸水平[9], 也未降低欧洲鳗鲡幼鱼肝脏不同种类胆汁酸水平[21]。因此, 肝脏中甘油磷脂类物质水平的升高, 可能用于胆汁酸的合成。
此外, 肝脏中磷脂酰胆碱水平上调还与亚油酸、α-亚麻酸和花生四烯酸代谢有关。亚油酸和α-亚麻酸作为鳗鲡等淡水鱼类主要的必需脂肪酸, 能够维持生物膜的正常结构和功能, 对胆固醇等类脂代谢具有重要作用[42]; 花生四烯酸在机体的营养物质代谢、白细胞功能调控和血小板激活中发挥着重要的作用[43], 还具有调节鱼体抗氧化能力、机体炎症反应及免疫功能[44]。甘油三酯是甘油酯类中最主要的中性脂质, 也是脂肪储存能量的主要形式,还可作为甘油磷脂生成的底物[45,46]。在本实验中,肝脏甘油三酯水平上调, 说明肝脏中储存的能量水平增加, 也可能用于与甘油磷脂代谢增强有关的代谢产物生成。在本实验中, 甘油磷脂代谢、甘油酯代谢、亚油酸代谢、α-亚油酸代谢及花生四烯酸代谢等分解代谢增强与BA组肝脏主要脂肪分解酶活性升高, 脂肪合成酶活性降低的结果相对应, 而生成的磷脂酰胆碱、甘油三酯等脂类物质含量上调, 可能是对肝脏保护作用及维持高强度代谢水平的能量需要的一种适应[46,47]。综上, 在饲料中添加BA对欧洲鳗鲡幼鱼肝脏中脂质代谢的影响可能主要通过调控甘油磷脂及甘油酯类, 详细作用机制还有待于进一步研究。
目前, 还未见其他水产动物的研究中使用靶向代谢组的方法, 分析BA对肝脏脂质代谢通路的影响, 本实验中显著变化的这些代谢通路仍需在转录水平、蛋白质翻译水平进一步验证。此外, 本实验中仅对对照组和BA1组开展了脂质组学分析, 主要根据本课题组以往BA添加在欧洲鳗鲡幼鱼中的实验结果[5], 推测2个BA添加水平组肝脏脂肪代谢的情况可能相近, 结合本实验中肝脏切片、肝组脂肪水平及脂肪代谢酶变化的结果可以证明原来的假设。而BA2组及更高BA添加水平对欧洲鳗鲡幼鱼肝脏脂质代谢通路影响的情况及作用机制还有待于在将来的研究中查明。
4 结论
在饲料中添加胆汁酸可通过降低欧洲鳗鲡幼鱼肝脏中脂肪合酶水平和增加脂肪分解酶活性, 上调肝脏磷脂酰胆碱和溶血磷脂酰胆碱含量, 主要增强甘油磷脂代谢和甘油酯代谢, 从而减少脂肪蓄积。本研究率先利用脂质组学技术分析胆汁酸对欧洲鳗鲡幼鱼肝脏脂肪代谢主要通路的影响, 研究结果可丰富胆汁酸调控鱼类脂肪代谢理论, 但显著变化的相关脂质代谢通路有待进一步验证, 详细的调控机制也需要在将来的研究查明。
