马铃薯抗晚疫病种质资源中抗病基因的筛选
2023-03-02李晓川王朝海王宗明宋治豪
李晓川, 王朝海, 周 平, 马 维, 陈 军, 陆 燚, 吴 显, 王宗明,吴 瑞, 宋治豪, 马 杰, 付 毅
(毕节市农业科学研究所, 贵州 毕节 551700)
0 引言
【研究意义】我国是马铃薯生产第一大国,而贵州所处的乌蒙山区是我国马铃薯的主产区,但同时由于气候冷凉潮湿,特别适合马铃薯晚疫病病原菌生长,易造成晚疫病大规模爆发。晚疫病由致病疫霉菌引起,是马铃薯生产的第一大病害[1-2]。晚疫病病原菌具有高度变异性,因此,研究新的晚疫病抗性基因对马铃薯晚疫病抗性育种具有重要意义。【前人研究进展】目前,已克隆获得多个晚疫病抗性R(Resistance)基因,均是NBS-LRR类基因[3-15]。其中,来源于马铃薯野生种Solanumdemissum(2n=6x=72)的R1位于第V号染色体上[3];R2和R2-like位于第IV号染色体上,在LRR2和LRR3间相差一段氨基酸序列[4];R3a和R3b位于第XI号染色体上,有73%的氨基酸序列一致性[5-6];R8位于第IX号染色体上[7]。来源于马铃薯野生种S.bulbocastanum(2n=2x=24)的Rpi-blb1/RB、Rpi-blb2和Rpi-blb3,被认为是具有广谱晚疫病抗性的基因,Rpi-blb1/RB定位于第Ⅷ号染色体上[8-9],Rpi-blb2位于第Ⅵ号染色体上与番茄Mi-1基因位点相同的区域[10],Rpi-blb3定位于第Ⅳ号染色体上[4],以及来源于S.venturii、S.americanum和S.stoloniferum的R基因,大多数R基因在不同马铃薯野生种中是同源基因[4,11-15]。最初晚疫病抗性基因的克隆主要通过图位克隆的方式,但随着发现R基因的结构上均为NBS-LRR类基因,利用不同的技术手段富集并挖掘抗晚疫病种质资源基因组或转录组内包含NBS-LRR保守序列的基因,进而筛选晚疫病抗性基因[13-15]的研究较多。【研究切入点】利用田间多点鉴定及室内接种晚疫病病原菌鉴定得到的野生种S.demissum品系cs47,通过转录组测序对晚疫病抗感材料的差异表达基因进行筛选,并根据已克隆晚疫病抗病基因在结构上的保守性,筛选差异表达基因中的马铃薯晚疫病抗病基因。【拟解决的关键问题】利用晚疫病抗病种质资源,通过转录组测序技术筛选晚疫病抗病基因,为马铃薯晚疫病抗性种质创新与利用提供理论依据。
1 材料与方法
1.1 材料
广谱晚疫病抗性材料野生种S.demissum品系cs47,通过室内接种晚疫病病原菌鉴定获得,在2018—2020年于马铃薯晚疫病高发重灾区的贵州省毕节七星关区和威宁彝族回族苗族自治县6个地点进行田间晚疫病鉴定,发现其在晚疫病大规模发病的早期、中期和晚期,叶片虽有晚疫病侵染发病的菌斑,但菌斑均未扩大,说明其拥有持续且较强的抗性。卡它丁(Katahdin)、大西洋(Atlantic)和底西瑞(Desiree)经过多年种植鉴定且查询欧洲马铃薯栽培种数据库(http://www.europotato.org/quick_search.php),3个品种均为易感晚疫病品种。
1.2 方法
1.2.1 病叶取样 分别取cs47、卡它丁、大西洋和底西瑞经晚疫病侵染的多片叶片,按单品种混合。样品经液氮速冻后保存备用。
1.2.2 转录组测序及分析 样品送安诺优达生物科技公司进行转录组文库构建和测序。经Illumina NovaSeq PE150高通量平台进行转录组测序,原始数据过滤掉低质量Reads(测序片段)和不合格序列后,完整Reads使用Bowtie2(--bowtie2-mismatch-rate)与马铃薯参考基因组(SolTub_3.0)进行比对。基于上述比对后,对各样本中比对到参考基因组的各unigene(条非重复序列)上的Reads进行定量,并进行FPKM(fragments per kilobase of exon model per million mapped reads)值的转换,从而获取各条unigene的表达水平[16-17]。利用Corrplot进行相关性分析。基于定量得到各unigene的FPKM值对样品间差异表达基因进行比较,利用DEGSeq进行cs47和卡他丁、cs47和底西瑞及cs47和大西洋的基因差异表达分析[18],并选取|log2Ratio|≥1(即基因间表达水平相差大于2倍)和Q<0.05的基因作为显著差异表达基因,并进一步选取基因间表达水平相差大于10倍的基因作为显著差异的表达基因。对获得的显著差异表达基因利用GO(Gene Ontology,基因本体论分析)数据库从生物过程(Biological process)、分子功能(Molecular function)和细胞成分(Cellular component)3个类别进行基因功能注释[19]。
1.2.3 序列比对及系统进化分析 基因的氨基酸序列利用Clustal Omega在线服务器(http://www.ebi.ac.uk/Tools/msa/clustalo/)进行多序列比对[20]。利用MEGA 6的Neighbor-Joining参数经过计算1 000次生成聚类树[21]。
2 结果与分析
2.1 转录组测序
将晚疫病抗病材料cs47和3个易感晚疫病品种进行转录组测序,过滤低质量Reads和不合格序列后,样品测得54 410 876~59 397 314个Reads。将完整Reads与马铃薯参考基因组进行比对,分别有4 602 576~40 166 116个Reads比对到参考基因组上。获取各条unigene的表达水平,利用DEGSeq分别进行3组样品间基因差异表达分析, cs47与卡它丁相比分别有6 464个和6 316个基因表达水平显著上升和显著下降(图1A);cs47与底西瑞相比分别有6 547个和6 264个基因表达水平显著上升和显著下降(图1B);cs47与大西洋相比分别有6 084个和6 264个基因表达水平显著上升和显著下降(图1C)。经计算,cs47与3个样品间的相关性系数(r)分别为0.79、0.71和0.74,而3个样品间的相关性系数分别为0.83、0.90和0.82,相关性系数越高,说明样本间的表达模式越接近。因此,cs47与易感晚疫病品种的表达模式差异大于易感晚疫病品种间的表达模式差异。
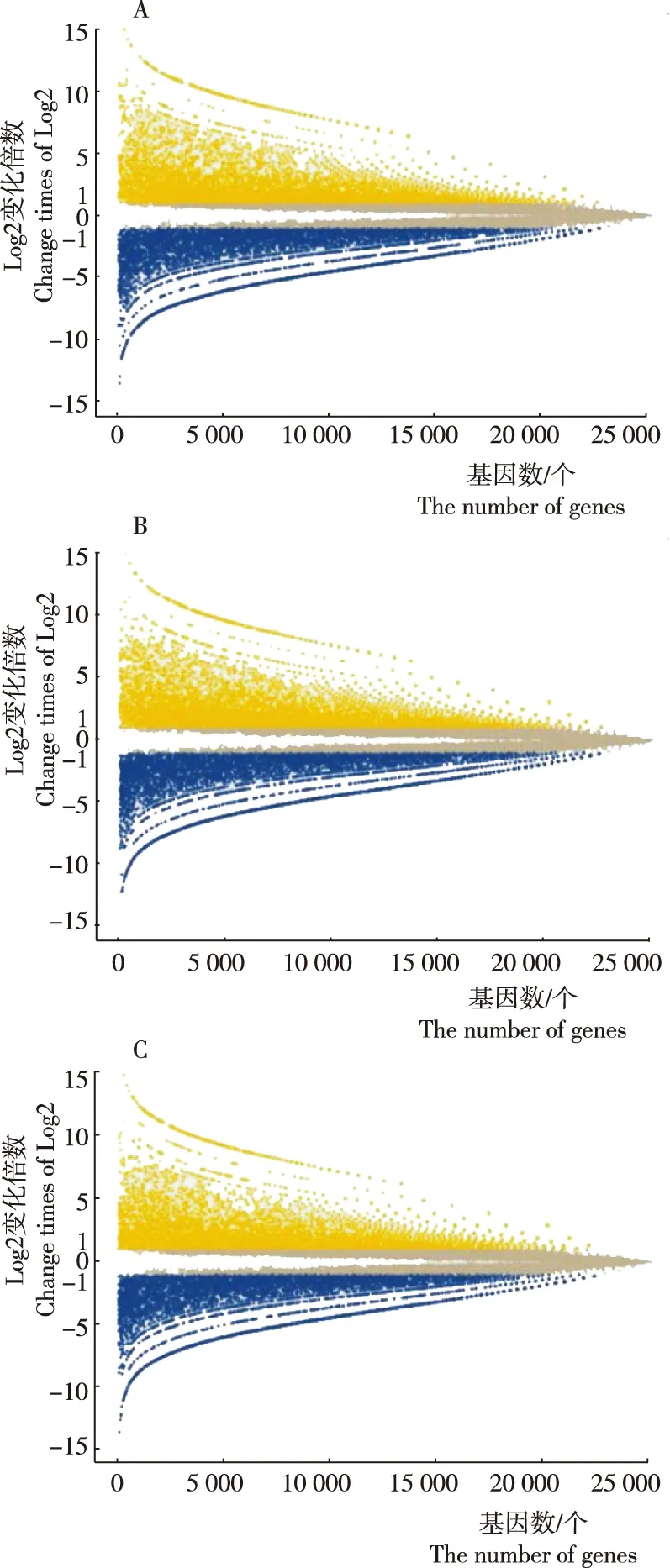
注:A为cs47与卡它丁对比,B为cs47与底西瑞对比,C为cs47与大西洋对比。其中黄点和蓝点分别代表基因表达显著上升和显著下降的基因。
2.2 差异表达基因的GO分析
cs47与卡它丁间的差异表达基因,在生物过程类别中涉及到免疫系统(immune system process)的分别有270个和223个表达显著上升和显著下降的基因,涉应激反应的(response to stimulus)分别有1 579个和1 616个表达显著上升和显著下降的基因(图2A)。cs47与底西瑞间的差异表达基因,在生物过程类别中涉及到免疫系统的分别有283个和210个表达显著上升和显著下降的基因,涉应激反应的分别有1 599个和1 619个表达显著上升和显著下降的基因(图2B)。cs47与大西洋间的差异表达基因,在生物过程类别中涉及免疫系统的分别有270个和193个表达显著上升和显著下降的基因,涉应激反应的分别有1 540个和1 540个表达显著上升和显著下降的基因(图2C)。

图2 3组对比显著差异性表达基因的GO基因功能注释
2.3 候选晚疫病抗性基因
在样品间基因表达水平相差10倍的基因中,有3个基因属于cc-NBS-LRR类,分别为PGSC0003DMP400045185、PGSC0003DMP400015105和PGSC0003DMP400015107,而当前已克隆马铃薯的抗晚疫病基因均属于cc-NBS-LRR类基因,将其氨基酸序列与已克隆的马铃薯抗晚疫病基因进行序列比对,分别由911个、1 243个和1 072个氨基酸组成。已克隆R基因和候选R基因的蛋白序列内均含有CC、NBS和LRR结构域区域(图3)。并对候选R基因和已克隆的R基因进行系统进化分析显示,PGSC0003DMP400045185和R1-A基因的亲缘关系较近,同时与R8以及Rpi-blb2基因的亲缘关系较近。PGSC0003DMP400015105和PGSC0003DMP400015107的氨基酸序列比较类似,与Rpi-pta1、Rpi-blb1/RB和Rpi-sto1基因的亲缘关系较近(图4)。

注:类似性质的氨基酸为同颜色字符;R 基因的CC结构域区域标注为黄色、NBS结构域区域标注为蓝色、LRR结构域区域标注为灰色。
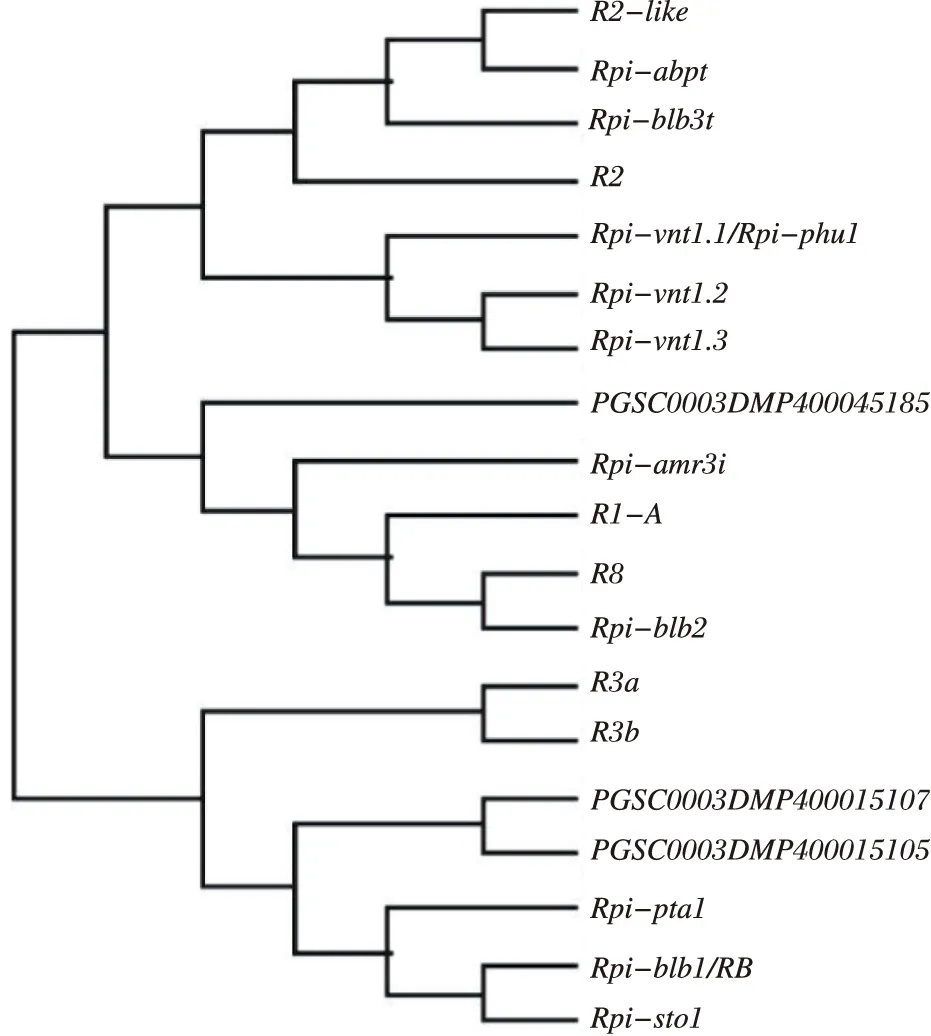
图4 3个晚疫病抗性基因与已克隆晚疫病抗性基因的系统进化树
3 讨论
通过转录组测序技术是分析马铃薯抗晚疫病种质资源中抗病基因的重要手段。在本研究中,通过分析抗晚疫病种质资源cs47与3个易感晚疫病品种的转录组差异表达基因发现,在不同材料中分别有6 084~6 547个和6 264~6 316个基因表达水平显著上升和显著下降。在不同材料中,涉及免疫系统的分别有270~283个和193~223个表达显著上升和显著下降的基因,涉及到涉应激反应的分别有1 540~1 599个和1 540~1 619个表达显著上升和显著下降的基因。
在更加显著差异表达的基因中,得到3个cc-NBS-LRR类基因,分别为PGSC0003DMP400045185、PGSC0003DMP400015105和PGSC0003DMP400015107。PGSC0003DMP400045185和R1-A的亲缘关系较近,同时又与R8以及Rpi-blb2基因的亲缘关系较近。Rpi-blb2克隆自马铃薯野生种S.bulbocastanum,是具有广谱晚疫病抗性的基因[10]。PGSC0003DMP400015105和PGSC0003DMP400015107与Rpi-pta1、Rpi-blb1/RB和Rpi-sto1基因的亲缘关系较近。Rpi-blb1/RB克隆自马铃薯野生种S.bulbocastanum,是第1个克隆得到的具有广谱晚疫病抗性的基因[8-9],Rpi-sto1和Rpi-pta1克隆自马铃薯野生种S.stoloniferum,是Rpi-blb1/RB的同源基因[11, 13]。
4 结论
通过转录组测序技术,利用马铃薯抗病种质资源广谱晚疫病抗性材料野生种S.demissum品系cs47,筛选得到PGSC0003DMP400045185、PGSC0003DMP400015105和PGSC0003DMP400015107共3个潜在的晚疫病抗性基因。转录组测序技术有助于分离晚疫病抗病基因,为晚疫病抗性育种提供理论基础。
