铁皮石斛DobHLH96基因克隆及表达分析
2023-02-15张红瑞于白音刘博婷刘羽佳
张红瑞, 理 雅,, 于白音,, 刘博婷, 陈 洁, 刘羽佳
[1.河南农业大学农学院,河南郑州 450046; 2.韶关学院英东生物与农业学院,广东韶关 512005;3.广东省粵北食药资源利用与保护重点实验室(韶关学院),广东韶关 512005]
植物在应对生物、非生物胁迫的过程中逐渐进化形成了由转录因子参与调控的极为复杂而高效的分子应答机制[1],转录因子是植物参与胁迫应答过程中具有重要调控作用的DNA结合蛋白,被激活后能够调控下游相关基因或其他转录因子表达,从而提高植物的抗逆能力[2]。bHLH转录因子(transcription factors,TFs)是植物中仅次于MYB TFs的第二大转录因子基因家族,由约60个氨基酸组成2个高度保守的结构域,N端的碱性区域能与顺式作用元件E-box(5′-CANNTC-3′)结合;螺旋区域通过2个α-螺旋的相互作用,形成同源/异源二聚体,从而与靶基因启动子的相关部位结合,调控下游基因表达[3]。已有研究发现,植物中的bHLH TFs参与植株的生长发育、逆境胁迫、调节信号转导与合成代谢等多种重要的生理代谢过程[4]。例如,在拟南芥(Arabidopsisthaliana)中异源过表达卷柏(Selaginellatamariscina)SlbHLHopt基因能够显著提高植株在缺水胁迫、盐胁迫下的种子萌发率、绿色子叶出苗率[5];拟南芥中2个bHLH TFs(AtLP1和AtLP2)可以调控植株叶片的长宽率及表皮毛形态[6];另有研究表明,拟南芥bHLH TFs能与蓝光受体隐花色素互作来调控开花时间,抑制AtbHLH113基因的表达,从而推迟植株开花、提高花青素含量[7-8];丹参(Salviamiltiorrhiza)中的SmbHLH92基因能够负调控植株毛状根中有关酚酸、丹参酮生物合成关键酶的基因转录水平[9]。由此可见,bHLH TFs在植株生长发育和次生代谢物质的合成过程中具有重要作用。另外,bHLH TFs能够参与植物响应非生物胁迫、病虫害等逆境胁迫过程,如百脉根(Lotusjaponicus)同源过表达LjbHLH7基因能够提高植株的抗虫性[10];同源过表达GmbHLH3基因的大豆(Glycinemax)毛状根复合植株和异源转基因拟南芥对Cl-/盐胁迫的耐受性增强[11];在拟南芥中异源过表达葡萄(Vitisvinifera)VvbHLH1基因能够增强转基因拟南芥的耐旱、耐盐能力[12];异源过表达甜菜(Betavulgaris)BvbHLH93基因的拟南芥能够通过增强抗氧化酶活性、减少活性氧(ROS)的产生来提高转基因植株对盐胁迫的耐受性[13];异源过表达苹果(Maluspumila)MdbHLH130基因能够提高转基因拟南芥的发芽率和存活率,增强其抗氧化性,缓解脱落酸(ABA)诱导的气孔关闭及叶片失水[14];过表达水稻(Oryzasativa)bHLH TF(OsPTF1)能提高缺磷条件下同源转基因植株的瞬时磷吸收速率[15]。另有研究发现,水稻中的OsbHLH6基因能够通过调节水杨酸、茉莉酸的信号转导来调节植株在感染稻瘟病时的防御机制[16];小麦(Triticumaestivum)过表达TabHLH49基因后能够提高植株的耐旱性[17];烟草(Nicotianatabacum)NtbHLH123基因能够调控下游NtCBF基因表达,其过表达能够增强转基因烟草中的抗氧化酶活性,从而调控植株的耐寒能力[18]。由此可见,bHLH TFs对植物适应和抵御不同逆境胁迫具有重要调控作用。
铁皮石斛(DendrobiumofficinaleKimura et Migo)是兰科石斛属多年生草本药用植物,其干燥茎被《中华人民共和国药典》历版单独收载,具有益胃生津、滋阴清热的功效,是我国传统名贵中药材,被誉为“中华九大仙草之首”,主要分布于我国安徽、浙江、福建等地。现代药理学研究发现,铁皮石斛中有多糖、生物碱、氨基酸等多种有效成分,具有增强免疫力、抗疲劳、抗氧化、降血压、保护肝脏等作用[19]。在自然条件下,野生铁皮石斛繁育率较低,加上近年来生存环境恶化、人为过度采挖等因素的影响,导致野生铁皮石斛种质资源遭到严重破坏,已濒临灭绝[20]。近些年来,随着组织培养、集成栽培技术的发展,人们已经完成从野生获取铁皮石斛到栽培铁皮石斛的转变,但目前还存在栽培品种混杂、成品质量良莠不齐等问题[21]。因此,深入研究铁皮石斛bHLH转录因子响应逆境胁迫的分子遗传机制,对于进一步通过遗传改良方法提高铁皮石斛药用有效成分含量及培育高产、优质、抗逆的药用铁皮石斛优良品种具有重要意义。本研究旨在通过筛选铁皮石斛转录组数据,克隆获得DobHLH96cDNA全长序列,并对其理化性质、蛋白结构、进化关系等进行预测分析,对其组织表达特性及其在低温、干旱胁迫和ABA处理下的表达模式进行初步分析,以期为进一步挖掘DobHLH96基因的生物学功能提供理论基础。
1 材料与方法
1.1 植物材料与处理
本研究所用试验材料为广东丹霞铁皮石斛,由韶关市石斛工程技术开发中心提供。将铁皮石斛蒴果进行消毒处理后,播种于生根培养基中,无菌培养约6个月后,选取生长状况一致的幼苗进行后续试验。以正常生长的植株为对照,将低温处理组置于4 ℃恒温、恒湿培养箱中;模拟干旱处理组用20%聚乙二醇(PEG)-6000进行浇灌,对照组浇灌水;ABA处理组用100 μmol/L ABA浇灌,对照组浇灌水;3个处理组均在处理0、1、3、6、9、12、24 h分别进行取样,用液氮处理1 min后,于-80 ℃超低温冰箱中保存。每个样品设3次生物学重复。
1.2 基因克隆
以铁皮石斛叶片RNA为模板,利用反转录试剂盒[购自宝生物工程(大连)有限公司,RR036A]合成cDNA。根据美国国家生物技术信息中心(NCBI)数据库中的云南铁皮石斛同源序列(序列号:LOC110116474)设计引物(F:5′-A T G T C A T T C G A A A T G C T T T C T T-3′;R:5′-T T A G A A C G A C A G G G G C G G G T T-3′),并委托生工生物工程(上海)股份有限公司合成。使用25.0 μL PCR扩增体系:12.5 μL 2×Master Mix(购自北京兰博利德生物技术有限公司,T0201),各1.0 μL上、下游引物,2.0 μL cDNA,用ddH2O补足至25.0 μL。PCR反应程序:94 ℃ 3 min;94 ℃ 30 s,58 ℃ 30 s,72 ℃ 60 s,30个循环;72 ℃ 7 min。将扩增产物连接至植物表达载体pROKⅡ中,转化大肠杆菌DH5α感受态细胞并选取阳性单克隆进行测序。
1.3 生物信息学分析
在NCBI数据库(https://www.ncbi.nlm.nih.gov)中使用Blast P检索其他物种与铁皮石斛DobHLH96蛋白的同源序列;用DNAMAN、MEGA 11邻接法对这些序列进行多序列比对和系统进化树的构建;蛋白结构域、理化性质分别用Pfam数据库(http://pfam.xfam.org/)、ExPASy ProtParam(https://web.expasy.org/protparam/)进行预测;二级结构、亲疏水性用SPLIT 4.0 SERVER(http://split.pmfst.hr/split/4/)进行分析;三级结构用SWISS-MODEL(https://swissmodel.expasy.org/)进行构建;通过New PLACE(https://www.dna.affrc.go.jp/PLACE/?action=newplace)对DobHLH96启动子的顺式作用元件进行预测分析。
1.4 基因表达分析
将上述样品抽提RNA后送至深圳华大基因科技有限公司构建RNA文库并测序(Illumina HiSeqTM2500/MiseqTM)。移除接头和低质量的原始数据(raw reads),用Tophat 2.0将高质量可用数据(clean reads)与铁皮石斛基因组参考序列进行比对,用Cufflinks进行组装,获得转录组数据,基因表达量以FPKM(fragments per kilobase million)表示,对铁皮石斛DobHLH96基因在低温、干旱和ABA胁迫处理下的表达模式进行分析。用SPSS 24.0软件对数据进行处理和显著性分析。
2 结果与分析
2.1 铁皮石斛DobHLH96的克隆与序列分析
以铁皮石斛叶片cDNA为模板,利用引物进行特异性扩增,电泳结果显示,特异性条带的长度在 1 000 bp 左右,测序获得铁皮石斛DobHLH96完整的开放阅读框全长序列为960 bp(图1-A),编码319个氨基酸残基(图2)。与参考序列(序列号:LOC110116474)进行对比发现有6个碱基差异,其中1个非同义替换,5个同义替换。基因结构和染色体定位分析结果显示,DobHLH96基因位于染色体骨架NW_021318852.1区域, 含有2个内含子、3个外显子(图1-B)。


2.2 铁皮石斛DobHLH96蛋白理化性质分析
ExPASy ProtParam分析结果显示,DobHLH96蛋白的分子式为C1 357H2 174N398O417S14,分子量为31.2 ku,理论等电点为6.51,不稳定性指数为82.59,是不稳定蛋白。用SPLIT 4.0 SERVER在线软件对其亲疏水性、蛋白二级结构进行预测,推测其为亲水性蛋白(图3-A),其二级结构由22.26% α-螺旋、9.09% β-折叠构象延伸链和68.65%无规则卷曲组成(图3-B)。用SWISS-MODEL构建的DobHLH96蛋白的三级结构如图3-C所示,可见DobHLH96蛋白与植物转录因子MYC2的相似度为31.25%。在Pfam上分析铁皮石斛DobHLH96蛋白的保守结构域,结果显示,DobHLH96蛋白具有HLH结构域(图4),属于bHLH TFs家族特有的基因结构域,表明本研究克隆获得的基因为DobHLH96。


2.3 DobHLH96蛋白的同源性和系统发育关系分析
在NCBI上利用Blast P对铁皮石斛的bHLH96氨基酸序列进行同源序列分析,选择不同物种同源性较高的序列进行比对发现,与其同源性最高的是鼓槌石斛(KAH0457194),相似度为96.5%,其次是建兰(QDL88322.1),相似度为84.95%,与小兰屿蝴蝶兰(XP_020572213.1)的相似度为83.39%,此外与海枣(XP_026662269.2)的相似度也较高。多重序列对比结果显示,DobHLH96蛋白序列与其他植株的bHLH蛋白序列均在88~139的位置含有HLH结构域(图5)。用MAGA 11.0构建进化树,发现铁皮石斛DobHLH96蛋白与鼓槌石斛bHLH蛋白聚在同一分支(图6),表明其亲缘关系最近。


2.4 DobHLH96的表达模式分析
为了研究DobHLH96在铁皮石斛不同组织中的表达情况,从NCBI中下载8个铁皮石斛组织(花柱、花蕾、叶、唇瓣、灰白根、绿根尖、萼片和茎)的差异表达数据[22]。结果显示,DobHLH96在铁皮石斛的不同部位均有表达,但是其相对表达量存在显著差异,相对表达量排序为花蕾>花柱>萼片>叶>灰白根>唇瓣>茎>绿根尖(图7)。

2.5 DobHLH96在不同胁迫下的表达分析
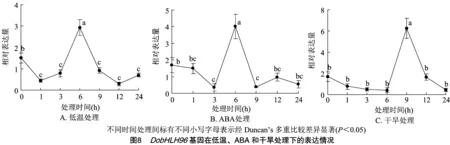
bHLH转录因子可响应多种非生物胁迫,从而提高植株的抗逆性。启动子元件分析结果显示,DobHLH96基因启动子序列中含有低温响应、干旱响应、水分胁迫响应以及ABA响应等元件(表1)。因此, 对DobHLH96基因在低温、干旱和ABA处理下的表达表达模式进行分析,结果表明,随着时间的增加,DobHLH96的相对表达量在低温处理下呈先下降后上升而后再下降的趋势,在处理后6 h达到最高值(图8-A)。在ABA处理下,DobHLH96相对表达量的变化趋势与低温处理的变化趋势基本一致,同样是先下降后上升再下降,且相对表达量在处理后6 h最高(图8-B),表明DobHLH96相对表达量在ABA处理、冷胁迫处理下具有相同的表达模式;在干旱处理下,DobHLH96相对表达量的变化趋势与ABA处理类似,其表达水平在处理后9 h最高(图8-C)。

表1 DobHLH96基因启动子元件分析
3 讨论
bHLH TFs广泛存在于动植物中,其蛋白结构最早于1989年在小鼠的肌肉中被发现并得到鉴定[23],目前已在多种植物中被发现并得到鉴定,其蛋白功能特性大多数是通过在模式植物拟南芥中研究发现的[4]。前人研究发现,真核生物中的bHLH家族成员分为六大类(A、B、C、D、E、F),6组成员能与不同碱基序列特异性结合,例如A组能与E-Box核心序列特异性结合,E组优先结合CACGNG序列,它们在生物体内发挥不同的生物学功能,其中D组缺少1个典型的碱性区域,被称为非典型的bHLH蛋白,主要与其他bHLH蛋白形成同源二聚体,从而抑制其转录活性[24]。本研究克隆得到的DobHLH96的开放阅读框全长序列为 960 bp,编码319个氨基酸残基,但是预测保守结构域时发现,该蛋白仅有1个HLH结构域,缺少碱性区域,属于D组的非典型bHLH蛋白。
bHLH TFs在植物不同组织器官中的表达水平存在显著差异,具有不同的功能。管丽婷研究发现,火龙果(Hylocereusundatus)HubHLH2基因主要在根中表达,而在茎、果实中的表达量较低[25];甘蓝型油菜(Brassicanapus)BnbHLH122-1在各组织器官中均有表达,从苗期到花期,在根、叶中的表达量升高,但在茎中的相对表达量始终较低[26];烟草NtbHLH112在叶中的相对表达量较高,尤其是在衰老叶片中的相对表达量最高,在根、花蕾中的相对表达量较低[27]。本研究发现,DobHLH96在花器官中的相对表达量最高,而在其他组织中的相对表达量较低,暗示该基因在调控铁皮石斛花器官的分化、生殖发育过程中发挥了重要作用。另外,顺式作用元件分析结果显示,DobHLH96启动子序列中含有与花粉及其他组织特异表达相关的元件,因此推测DobHLH96基因编码非典型的bHLH蛋白可能与其他bHLH蛋白互作,在铁皮石斛开花和生殖发育过程中起着重要的调控作用。
bHLH TFs在植物应对非生物胁迫的应答过程中具有重要的调控作用,其参与调控逆境的信号通路可以分为3种:独立于ABA的信号通路、依赖于ABA的信号通路以及2种信号通路相互重叠[4]。以往的研究表明,ICE1-CBF-COR是植物抵御低温胁迫的主要信号转导途径[28],如在柠檬(Citrus×limon)中异源过表达枳(Poncirustrifoliata)PtrICE1基因能够提高转基因植株中的抗氧化酶活性,减少活性氧的积累,进而提高植株对低温的耐受性[29];在烟草中过表达枳PtrbHLH基因能够显著增强植株内的POD活性,提高转基因植株在低温条件下的耐寒性[30]。本研究发现,铁皮石斛DobHLH96基因在低温和ABA处理下的表达模式一致,暗示DobHLH96基因可能是通过依赖于ABA的信号通路响应低温胁迫。值得一提的是,本研究还在DobHLH96启动子序列中发现了MYCCONSENSUSAT(CANNTG)元件,该元件为ICE1-CBF-COR调控网络中ICE1能够识别并结合的1个关键序列[31]。因此,推测DobHLH96基因可能在有ABA存在时通过ABA信号途径调控下游基因的表达,从而参与铁皮石斛低温胁迫应答过程。在没有ABA存在时,DobHLH96可能通过ICE1-CBF-COR低温信号途径抵御低温胁迫,但具体响应机制尚未可知。此外,前人研究发现,bHLH TFs在干旱胁迫应答时主要通过调节ABA的敏感性、气孔开闭、叶毛状体和根毛的发育来增强植株对干旱的耐受性[32]。例如在拟南芥中过表达苹果(Maluspumila)bHLH转录因子MdCIB1基因能通过提高脯氨酸含量、降低过氧化物积累及增强抗氧化酶活性来增强转基因植株的耐旱能力[33];苦荞(Fagopyrumtataricum)FtbHLH3基因在拟南芥中异源过表达,转基因植株能通过ABA依赖的信号途径正向调节干旱/氧化胁迫耐受性[34];在水分胁迫下,外源ABA处理可提高甘蔗(Saccharumofficinarum)内源ABA含量,增强抗氧化酶活性[35]。上述研究均证实,ABA能通过感知和传递逆境信号,提高ABA的信号转导能力,进一步调控植株的抗逆性。本研究发现,DobHLH96基因的表达明显受到低温、干旱和ABA处理的诱导,并且在3种处理下,DobHLH96的表达水平整体呈现相似的变化趋势。另外,DobHLH96基因启动子序列含有与非生物胁迫、ABA、低温及脱水响应相关的顺式作用元件,提示DobHLH96基因可能通过ABA介导的信号途径来响应低温、干旱胁迫,但具体分子调控机制暂不明确。因此,今后应深入挖掘DobHLH96基因响应干旱和低温胁迫的分子调控机制,为铁皮石斛的遗传改良提供进一步的理论基础。
