低氧胁迫和复氧对长吻鮠鳃组织低氧应答基因和生理生化指标的影响
2023-02-14杨智茹程景颢张国松尹绍武
李 谣,杨智茹,程景颢,李 杰,王 涛,张 凯,张国松,尹绍武
(1南京师范大学海洋科学与工程学院/江苏省特色水产育种与绿色高效养殖技术工程研究中心,南京 210023;2菏泽学院农业与生物工程学院/山东省高校生理生化及应用重点实验室,山东菏泽 274015)
0 引言
在自然环境中,溶解氧(DO)是水生生物生存不可缺少的条件[1]。由于水温、昼夜变化等原因,水生态系统中DO浓度不断变化,导致鱼类常常暴露于低氧环境中。鱼类已进化出多种适应策略应对低氧环境,但是当DO长时间处于较低水平或者严重低氧时,仍会对其生长产生不利影响,甚至导致死亡,给水产养殖业带来严重损失[2]。鳃组织是鱼类主要的呼吸器官,具有气体交换、排泄废物和渗透压调节等重要功能[3-4],研究长吻鮠鳃组织在低氧胁迫和复氧下低氧应答基因和生理生化指标的变化,可以为低氧应激的分子机制研究提供基础资料,并为其健康养殖提供参考。目前,研究人员针对低氧环境给鱼类鳃组织带来的影响已展开部分研究,如低氧胁迫可引起鲫鱼(Carassius auratus)[5]、花斑溪鳉(Rivulusmarmoratus)[6]和团头鲂(Megalobrama amblycephala)[7]分化迁移鳃小片间细胞团或提高平均突出片层高度和平均片层面来增加鱼体的呼吸表面积。在分子水平上,鱼类鳃组织中氧传感蛋白(包含HIFs、PHDs、Vhl等)作为低溶解氧反应的核心调节因子,在低氧环境中被不同程度的激活[8],如大弹 涂 鱼 (Boleophthalmus pectinirostris)[9]、多 鳞 鱚(Sillago sihama)[10]和斑点叉尾鮰(Ictalurus punctatus)[11]等。同时氧传感蛋白主导的HIF signaling pathway通过激活一系列相关基因致使鳃组织代谢模式由有氧呼吸向无氧呼吸转变[12],如翘嘴鳜(Siniperca chuatsi)在急性低氧下糖酵解酶活力显著增加,此现象也同样出现在草鱼(Ctenopharyngodon idella)和卵形鲳鲹(Trachinotus ovatus)中[13-14]。常氧下鱼体内线粒体会产生少量的活性氧(ROS)来满足正常的生理活动[15],而低氧胁迫下鲫鱼[16]、鲢(Hypophthalmichthys molitrix)[17]和花鲈幼鱼(Lateolabrax japonicus)[18]鳃组织中氧化应激指标和抗氧化酶活性显著上调。这些氧化应激状态可能会导致细胞凋亡产生,如Jun等[19]研究发现低氧胁迫能显著提高大口黑鲈(Micropterus salmoides)鳃组织促凋亡基因的表达水平,另外也有报道称鳃组织的细胞凋亡是发生在鳃丝间,目的是为了在低氧下增加鳃组织的呼吸表面积[20]。
长吻鮠(L.longirostris)隶属于鲇形目(Siluriformes)、鲿科(Bagridae)、鮠属(Leiocassis),是一种主要生活在长江、淮河、珠江等水域的底层经济鱼类,具有营养丰富、肉质细腻、生长速度较快等特点[21],深受广大消费者喜爱。由于阴雨、高温、高密度放养等原因导致养殖水体内的DO含量减少,致使长吻鮠经常产生浮头,甚至窒息或死亡。因此,研究该鱼低氧胁迫的分子调控机制显得尤为重要。目前国内外关于该鱼的研究主要集中在其繁养殖技术[22]、产品加工[23]等方面,针对其鳃组织低氧应答基因和生理生化指标的研究尚未见报道。本研究以长吻鮠为对象,研究低氧胁迫和复氧对其鳃组织氧传感蛋白、呼吸代谢、氧化应激、组织形态和细胞凋亡的影响,系统地探讨该鱼低氧应答基因和生理生化指标的变化,初步揭示该鱼鳃组织应对低氧胁迫的分子调控机制。
1 材料与方法
1.1 试验材料
试验所用长吻鮠采自江苏省南京市水产科学研究所禄口基地,选取500尾体格健壮、规格均匀的长吻鮠,体长(12±1.1)cm,体重(30±2.3)g消毒处理后,随机转移到10个具有生物过滤器的水循环养殖玻璃缸(1.2 m×0.85 m×0.55 m)中驯养14天,每缸50尾。试验水温为(27±1)℃,DO浓度为(7.3±0.5)mg/L,水的流速为5 L/min,每天投喂人工配合饲料2次,时间为9时和17时。试验前禁食24 h。试验于2021年3—10月在江苏省南京市水产科学研究所禄口基地进行。
1.2 试验设计与样本采集
试验设置2个处理组:低氧胁迫(0.8±0.1)mg/L和复氧(7.3±0.5)mg/L,每组3个重复。在低氧处理前,分别从常氧状态[DO浓度为(7.3±0.5)mg/L]的3个水循环养殖玻璃缸中各取4尾长吻鮠鳃组织作为对照组(标记为C),共12尾,其中同1个水循环养殖玻璃缸中3尾鱼的鳃组织混为1个样本,液氮处理后-80℃保存,用于检测基因表达量和生理生化指标;另外3尾鱼鳃组织(第二鳃弓)保存于加有多聚甲醛的离心管中,用于切片检测。随后,关闭水循环养殖玻璃缸的充气设备直接将纯氮气充入水体30 min左右,使水中DO浓度降至(0.8±0.1)mg/L后,调节氮气的冲入量使DO浓度稳定在(0.8±0.1)mg/L,分别在0、2、4、6 h(记为H0、H2、H4、H6)进行取材,每个水循环养殖玻璃缸取4尾。低氧胁迫试验结束后停止充入氮气并充入空气,待水中DO浓度恢复至(7.3±0.5)mg/L时,调节空气冲入量维持水中DO含量,分别在2、4、6 h(记为R2、R4、R6)进行取材,每个水循环养殖玻璃缸取4尾。整个试验期间每5 min使用溶氧测定仪(LDO101,上海鑫嵩公司)检测一次水体中DO浓度。
1.3 鳃组织实时荧光定量PCR(qRT-PCR)检测
每个时段各取3个样提取总RNA,使用反转录试剂盒(Vazyme,南京)得到cDNA。在已有的转录组序列中获取基因CDs序列,使用Premier 5.0软件设计基因qRT-PCR上下游引物(见表1),其中β-actin为内参基因。qRT-PCR(Light Cycler®96实时荧光定量PCR仪,Roche)反应体系为正、反向引物各0.45μL,Mix10μL,ddH2O 7.1 μL,模板cDNA2 μL,总体积为20 μL。反应条件为95℃ 5 min,1个循环;95℃ 10 s,60℃ 30 s,40个循环。使用2-ΔΔCt公式计算基因的相对表达量。

表1 引物列表

续表1
1.4 鳃组织酶活性检测
在冰浴条件下准确称取鳃组织0.1 g,迅速剪碎,按1:9比例加入0.9%的生理盐水,机械匀浆之后经低温离心机3000 r/min离心10 min,取上清液分装保存于-20℃冰箱备测。鳃组织匀浆上清液中的蛋白浓度(TP)、谷胱甘肽过氧化物酶(GSH-Px)活力、过氧化氢酶(CAT)活力、超氧化物歧化酶(SOD)活力、果糖磷酸激酶(PFK)活力、己糖激酶(HK)活力、丙酮酸激酶(PK)活力、乳酸脱氢酶(LDH)活力、琥珀酸脱氢酶(SDH)活力、苹果酸脱氢酶(MDH)活力、丙二醛(MDA)和脂质过氧化物(LPO)含量均使用南京建成生物工程研究所所售的试剂盒测定。
1.5 鳃组织苏木精-伊红(H&E)染色切片
将固定于4%多聚甲醛中的鳃组织,经石蜡切片脱蜡至水→苏木素染色→伊红染色→脱水封片后使用显微镜观察并拍照,细胞核和细胞质分别呈蓝色和红色。
1.6 鳃组织TUNEL切片检测
将保存在4%多聚甲醛溶液中的鳃组织进行常规石蜡包埋并切片,操作步骤如下:石蜡切片,脱蜡水化,PBS漂洗,通透,PBS漂洗,室温平衡,DAB显色,PBS漂洗,苏木素复染细胞核,PBS漂洗,脱水中性树胶封片和显微镜观察。
1.7 数据统计分析
使用SPSS 22.0软件对数据进行统计分析。对照组和低氧组表达模式的数据采用双尾t-test检验统计差异,当P-value<0.05时认为差异显著(标为*),P-value<0.01时认为差异显著(标为**),P值均为试验组与对照组相比得出,结果用平均值±标准差(mean±SD)表示。
2 结果与分析
2.1 低氧胁迫和复氧对长吻鮠鳃组织氧传感蛋白的影响
如图1所示,低氧胁迫下长吻鮠鳃组织中HIF-1α、HIF-2α、PHD1和PHD3基因的表达量在H4到达顶峰,复氧后逐渐下降,R6较对照组仍有显著差异(P<0.05)。PHD2和Vhl基因的表达量在H2时达到峰值,复氧6 h后仍显著高于对照组(P<0.05)。
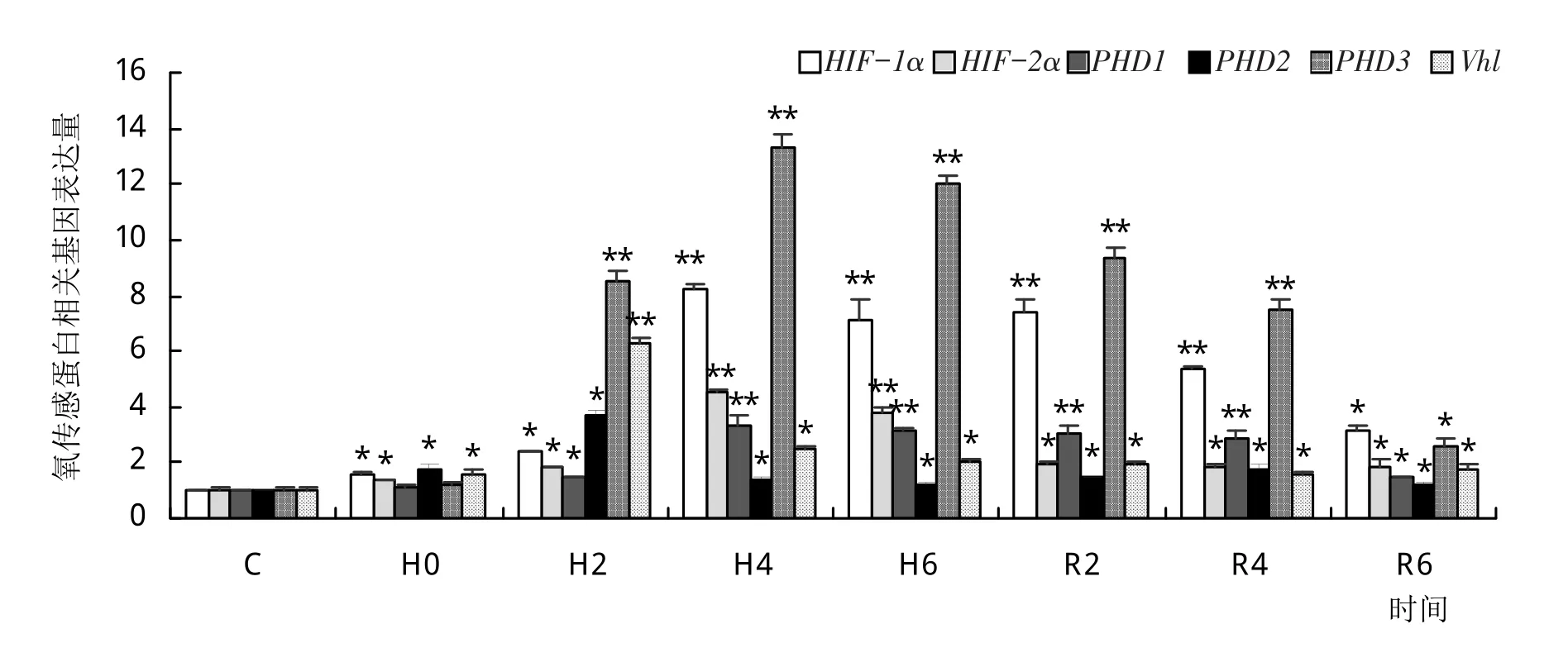
图1 低氧胁迫和复氧下长吻鮠的鳃组织氧传感蛋白基因的表达模式
2.2 低氧胁迫和复氧对长吻鮠鳃组织呼吸代谢相关酶活性的影响
在低氧胁迫下长吻鮠鳃组织中PFK和HK活性显著升高,在R2时达到峰值,随后逐渐降低,R4时活性较对照组无显著差异(P>0.05)。PK和LDH活性分别在H6和H2时到达峰值,复氧后逐渐恢复至对照组水平。SDH和MDH活性分别在H6和H4时显著低于对照组(P<0.05),复氧6 h后与对照组已无显著性差异(P>0.05)(图2)。
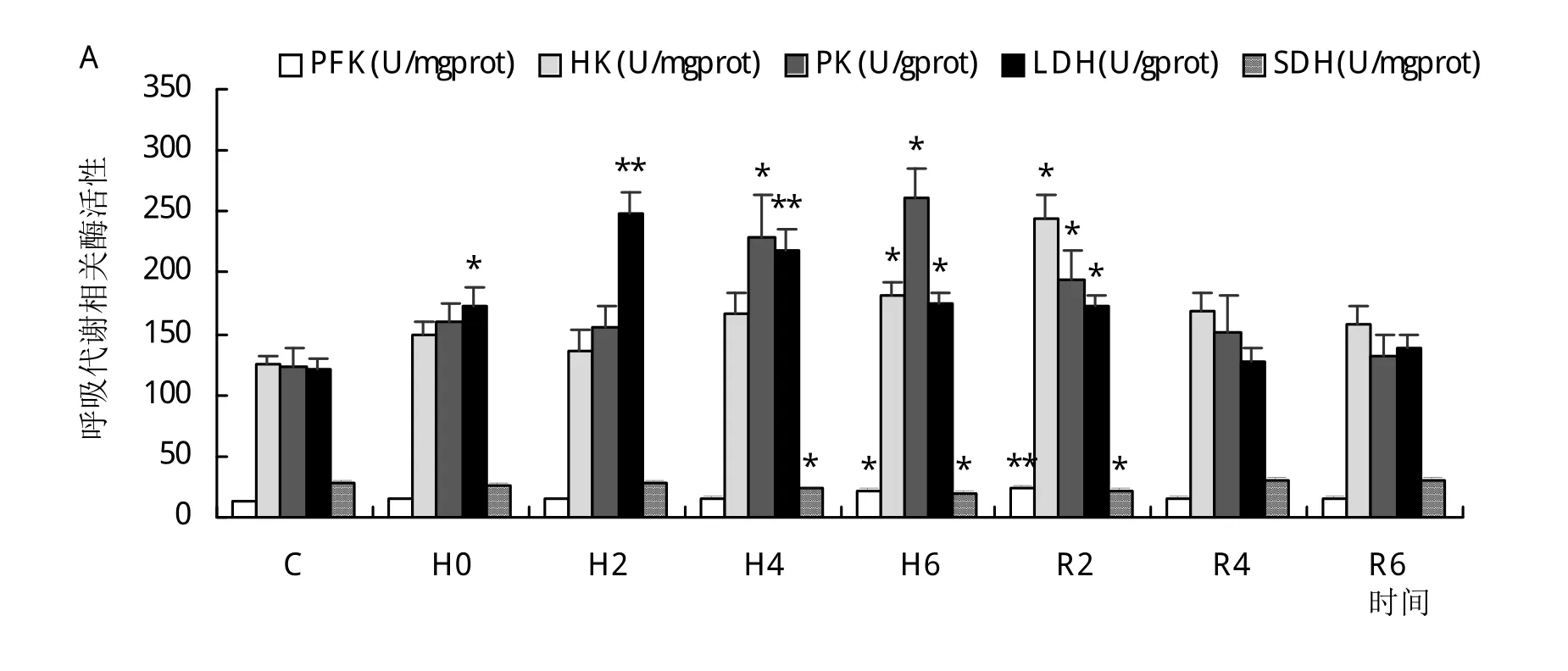
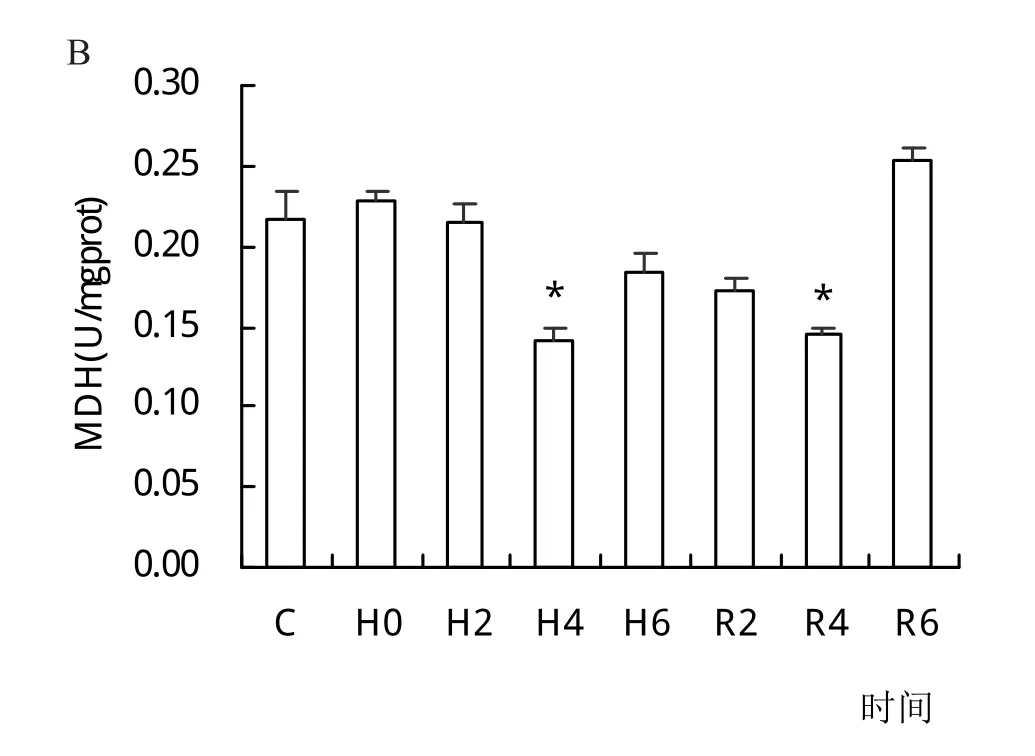
图2 低氧胁迫与复氧对长吻鮠鳃组织PFK、HK、PK、LDH、SDH、MDH活性的影响
2.3 低氧胁迫和复氧对长吻鮠鳃组织氧化应激酶活性与参数的影响
如图3所示,长吻鮠在低氧胁迫下GSH-Px和LPO活性在H4和H6时达到顶峰,随后逐渐下降,R6较对照组已无显著性差异(P>0.05)。CAT活性在低氧胁迫下不断上升,在H2时到达顶峰,恢复溶氧后呈下降趋势,R6较对照组仍有显著性差异(P<0.05)。SOD和MDA活性分别在H6和H4时达到峰值,随着溶氧的恢复其活性逐渐下降,R2时已恢复至对照水平。
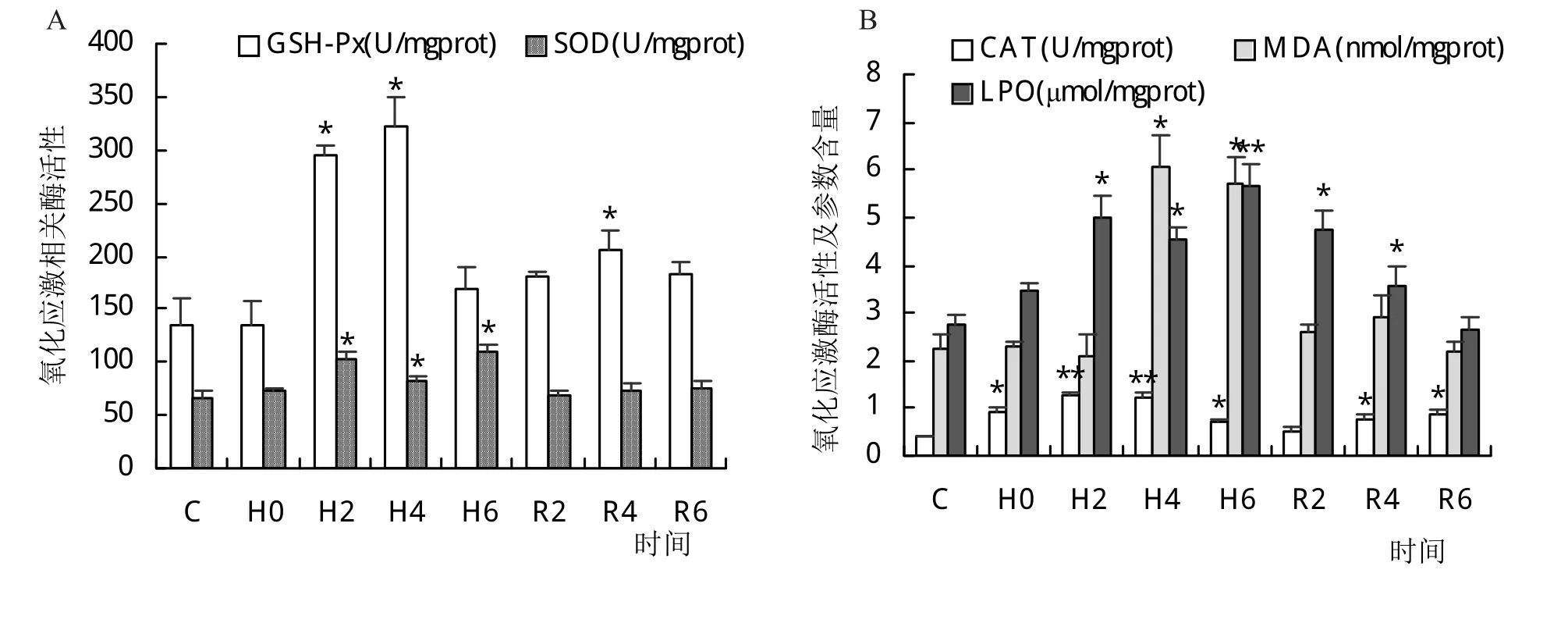
图3 低氧胁迫与复氧对长吻鮠鳃组织GSH-Px、SOD、CAT、MDA和LPO活性的影响
2.4 低氧胁迫和复氧对长吻鮠鳃组织结构的影响
如图4所示,在常氧状态下长吻鮠鳃丝两侧鳃小片排列整齐,形状良好,线粒体丰富细胞聚集在鳃小片基部,血细胞排列有序,低氧胁迫后鳃组织上皮出现了抬升,随着低氧时间的延长部分血细胞大量聚集使鳃小片顶端呈棒状,并发生线粒体丰富细胞肿胀和增生等病变现象,在溶氧恢复后,低氧引起的鳃组织变化并未得到改善。
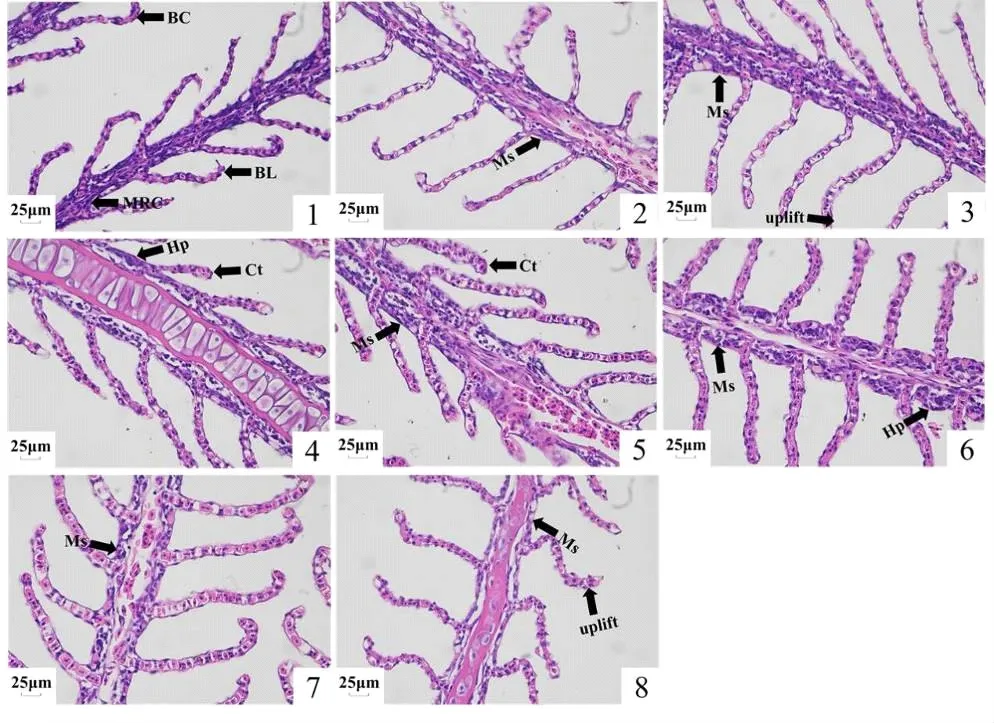
图4 低氧胁迫和复氧下长吻鮠鳃组织形态
2.5 低氧胁迫和复氧对长吻鮠鳃组织细胞凋亡的影响
2.5.1 鳃组织TUNEL结果分析 低氧胁迫下,长吻鮠鳃组织细胞凋亡程度不断增加,复氧6 h后细胞凋亡现象仍然存在,对凋亡指数统计分析显示,低氧可促进鳃组织发生凋亡,各处理组之间的凋亡指数随着低氧时间的延长逐渐增加,在H6时到达顶峰,复氧后不断下降,R6细胞凋亡指数较对照组仍有显著差异(P<0.05)(图5)。

图5 鳃组织TUNEL切片检测和凋亡指数统计结果
2.5.2 凋亡基因的时序表达分析 在低氧胁迫下,长吻鮠鳃组织中Bax和Apaf-1基因的表达量在H0时显著上升,复氧后逐渐回落,R6较对照组仍有显著性差异(P<0.05)。Caspase 3和p53基因的表达量在低氧胁迫下显著升高,在H2时达到峰值,复氧6 h后逐渐恢复至对照组水平。Bcl-2基因的表达量在H6时显著低于对照组(P<0.01),复氧后表达量逐渐上升,R6与对照组相比仍有显著差异(P<0.05)(图6)。
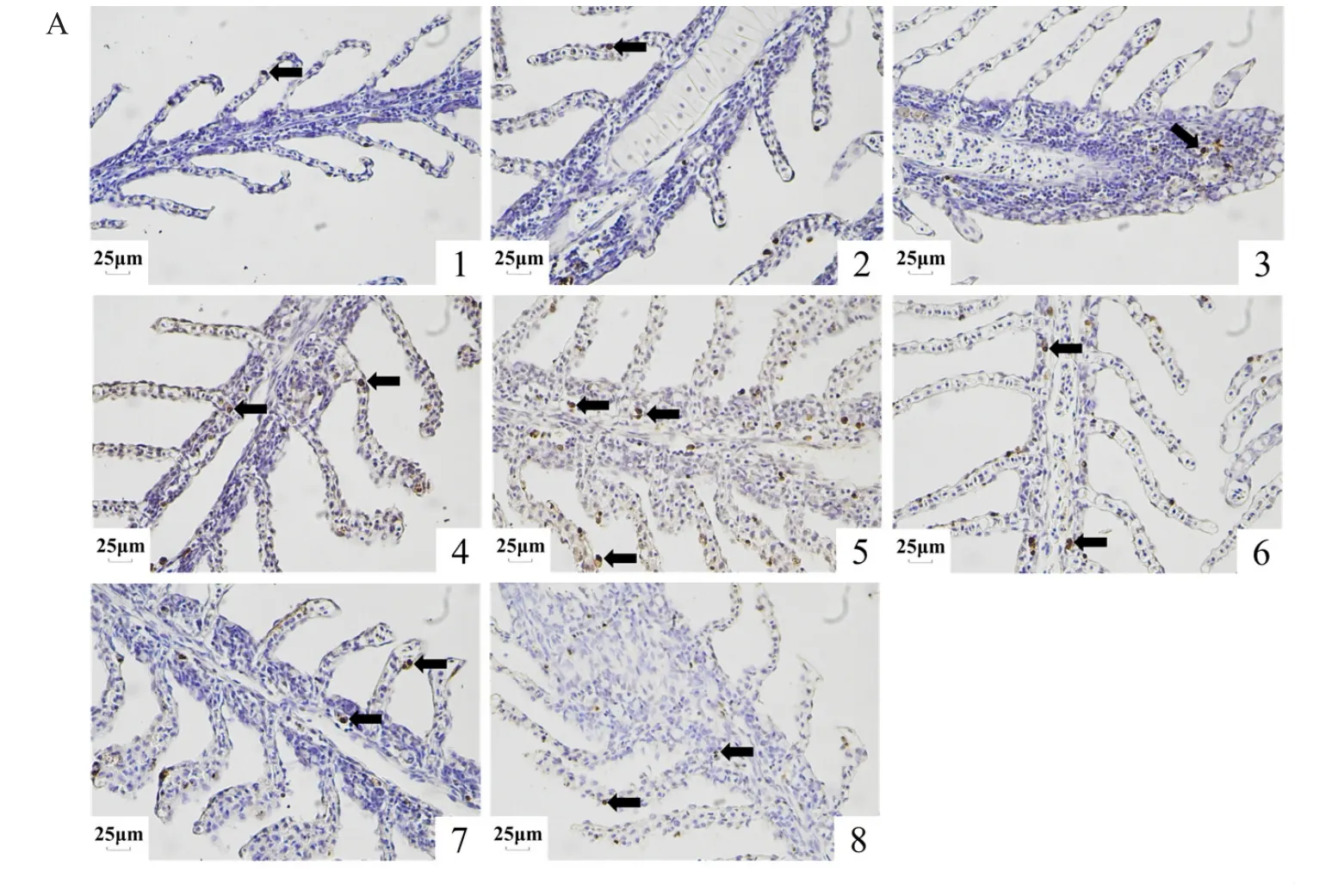

图6 低氧胁迫和复氧下长吻鮠鳃组织凋亡相关基因的表达模式
3 讨论
3.1 低氧胁迫和复氧对长吻鮠鳃组织氧传感蛋白和呼吸代谢的影响
在自然水体中,鱼类为了减轻DO浓度剧烈变化所造成的损害,演化出了许多复杂的低氧反应机制[8],如HIFs、PHDs和Vhl等氧传感蛋白的低氧诱导研究被广泛报道,麦穗鱼(Pseudorasbora parva)在低氧胁迫处理6 h时鳃中HIF-1α基因表达量到达峰值[24],大弹涂鱼在低氧处理后鳃组织中HIF-1α基因表达量显著高于对照组[9],多鳞鱚在低氧处理后鳃组织中PHDs、Vhl的表达量显著高于常氧组[25],同样低氧胁迫能显著诱导鲫鱼[26]、斑点叉尾鮰[11]和半滑舌鳎(Cynoglossus semilaevis)[27]鳃组织氧传感蛋白相关基因的表达。本研究发现,随低氧胁迫时间的延长,长吻鮠鳃组织中氧传感蛋白相关基因HIF-1α、HIF-2α、PHDs和Vhl的表达量整体呈逐渐上升趋势,复氧后其表达量逐渐下降但仍有较高的表达量,其中,PHD1、PHD2、PHD3和Vhl基因的高表达可能充当终止低氧反应的反馈调节机制,防止长吻鮠鳃组织一直处于低氧效应期[28];以上结果表明长吻鮠与上述鱼类氧传感蛋白的功能类似,氧传感蛋白在进化过程中具有一定的保守性。值得注意的是,复氧后HIF-1α、HIF-2α、PHDs和Vhl仍有较高的表达量,类似现象也出现在瓦氏黄颡鱼(Pelteobagrus vachelli)脑组织中[12],这可能是由于鳃组织是鱼体重要呼吸器官,在复氧过程中部分氧传感蛋白基因需要通过持续高表达来缓冲低氧造成的影响,使机体快速恢复到稳定状态。
在低氧胁迫下细胞可通过改变代谢途径、代谢方式来减轻低氧产生的不利影响[29]。有研究表明,SDH和MDH参与三羧酸循环是该循环的关键酶,PFK、HK和PK是糖酵解过程中的限速酶,LDH是无氧呼吸的指示剂。钱辰颖等[30]研究发现,团头鲂鳃组织在低氧胁迫下LDH活性显著升高,SDH活性显著降低。张国松等[12]研究发现,瓦氏黄颡鱼大脑和肝脏中糖酵解相关酶(PFK、HK和PK)活性和LDH活性随低氧胁迫时间延长逐渐增加。在本研究中,长吻鮠在低氧胁迫下鳃组织糖酵解关键酶(PFK、HK和PK)、LDH的活性与对照组相比显著升高,TCA循环的关键酶SDH和MDH活性显著降低,复氧后呈上升趋势,表明低氧胁迫可以抑制长吻鮠鳃组织三羧酸循环途径,而增强了无氧呼吸相关酶活性,如提高糖酵解活动,以及LDH活性催化丙酮酸转化为乳酸,从而增加无氧呼吸;原因可能是在低氧胁迫下长吻鮠体内ATP产生减少,鱼体需要通过增强无氧呼吸来弥补有氧呼吸活动抑制导致的能量缺失[31]。
3.2 低氧胁迫和复氧对长吻鮠鳃组织氧化应激、细胞凋亡和组织结构的影响
生物体内ROS增加与消除速率处于动态平衡状态,在多种形式胁迫下体内ROS不断积累从而造成机体发生氧化应激损伤[32],而生物体可通过激活机体抗氧化防御系统来减轻ROS增加造成的损伤。贾秀琪等[33]研究发现,河川沙塘鳢(Odontobutis potamophilus)在急性低氧5 h时,鳃组织中CAT、GSH-Px活性和SOD含量显著升高。Huang等[34]研究发现,在低氧胁迫下金曼龙(Trichogaster microlepis)鳃组织中CAT、GSH-Px和SOD的活性均增加。本研究发现低氧胁迫能显著提高长吻鮠鳃组织MDA和LPO的含量和GSH-Px、CAT和SOD活性。然而,CAT活性在复氧6 h后较对照组仍有显著差异,说明在低氧条件下长吻鮠鳃组织可通过提高抗氧化防御系统(CAT、GSH-Px和SOD)来减少氧化应激对鳃组织的伤害,而在复氧过程中随着环境中氧气大量导入,导致机体代谢补偿增强,有氧呼吸增强引起ROS浓度上升,从而再次出现氧化应激现象;此结果也印证了M.Hermes-Lima等[35]所提出的关于“氧化应激准备(在低氧条件下动物能够通过提高自身的抗氧化潜力,从而为解决再氧化后体内的氧化应激做好准备)”的猜想。
有研究显示,在动物组织低氧或缺血过程中,细胞出现代谢紊乱、细胞周期阻滞等,同时线粒体功能受损生成过量ROS,引起细胞凋亡[36]。丁晨雨等[37]研究发现,低氧胁迫致使鲢心肌细胞发生了凋亡,并随着低氧浓度下降凋亡指数逐渐升高。本研究采用TUNEL检测发现,低氧胁迫会造成长吻鮠鳃组织细胞凋亡加剧,对凋亡细胞统计发现,其数量随着低氧时间的延长逐渐增加,在复氧6 h时仍有较多的凋亡细胞数量,说明低氧胁迫能显著造成长吻鮠鳃组织细胞发生凋亡,但凋亡加剧的起始溶解氧浓度还有待进一步研究。Bcl-2和Bax基因在鱼类细胞凋亡过程中起着关键作用,二者间的比率关系是决定细胞存亡的关键[38]。Yuan等[39]研究发现,斑点叉尾鮰在低氧处理后其组织中Bcl-2基因的表达受到抑制。本研究结果显示长吻鮠鳃组织中Bcl-2的表达量在低氧胁迫下显著降低,而Bax的表达量显著升高,在复氧过程中Bcl-2、Bax的表达量较对照组仍有显著差异,这可能是由于Bax/Bax同源二聚体的数量在低氧胁迫和复氧下明显增加,鳃组织细胞对凋亡信号的反应性持续增强,从而促进了细胞凋亡。此外,裴雪莹等[40]研究发现,杂交黄颡鱼“黄优1号”在低氧条件下,细胞凋亡相关基因(Apaf-1、Caspase 3、Caspase 9和p53)的表达量均显著高于对照组。本研究发现Caspase 3、p53和Apaf-1基因表达量在低氧胁迫下显著升高,说明长吻鮠鳃组织细胞凋亡反应在低氧胁迫下进一步加剧。值得注意的是,复氧6 h后Apaf-1基因的表达量仍显著高于对照组,出现这种现象的原因可能是复氧过程中重新输入的氧气使鱼体再次产生了氧化应激反应,从而产生了细胞凋亡程序。
鱼类鳃组织肩负着呼吸、排泄、调节渗透压、维持酸碱平衡的功能,相比其他器官更容易受到低氧的影响,从而危及鱼体的正常生长[41]。有研究发现低氧胁迫会使鱼类鳃组织发生上皮抬升、增生、线粒体丰富细胞肿胀、鳃小片棒状化等结构变化[42]。陈世喜等[43]研究发现,急性低氧胁迫致会使卵形鲳鲹(Trachinotus ovatus)鳃小片上皮出现肿胀和抬升;随低氧胁迫时间的延长,会出现鳃小片上皮细胞分离现象,从而增加鳃呼吸表面积。本研究中通过H&E染色切片观察发现,在低氧胁迫下长吻鮠鳃组织上皮出现了抬升,随着低氧时间的延长部分血细胞大量聚集使鳃小片顶端呈棒状,并发生线粒体丰富细胞肿胀和增生等现象,在溶氧恢复后,低氧引起的鳃组织的变化并未得到改善,表明低氧胁迫显著影响了长吻鮠鳃组织的正常组织形态。
4 结论
本研究对低氧胁迫和复氧下长吻鮠鳃组织低氧应答基因和生理生化指标进行了系统分析,发现长吻鮠在低氧胁迫和复氧下可以通过迅速启动氧传感通路,上调PHDs和Vhl基因的表达量来终止低氧反应的反馈调节;通过提高无氧呼吸活动来补偿氧气供应不足所导致的ATP紧缺。随着低氧时间的延长,鳃组织上皮抬升、鳃小片顶端棒状、线粒体丰富细胞肿胀和增生等现象加剧,同时产生了氧化应激损伤与细胞凋亡,机体通过激活抗氧化防御系统来维持体内环境稳态,本研究结果为阐明低氧胁迫和复氧下鱼类鳃组织低氧应答与生理生化机制提供了参考资料。
