特种稻研究进展
2023-02-11李潜龙康云海郑国利赵玉军
杜 明 李潜龙 康云海 郑国利 赵玉军 方 玉,2
(1 上海中科荃银分子育种技术有限公司,上海 200030;2 安徽荃银高科股份有限公司,合肥 230088)
水稻是我国重要的粮食作物,在我国粮食生产中占据重要的地位。全球大约有65%的人口以大米作为主食[1]。专用稻是指稻谷、米粒或植株具有专门用途的栽培稻。目前,随着我国饲料工业、食品加工业和旅游业的迅速发展,专用稻的需求在增加。一方面,随着人民生活水平的提高,对于肉、蛋、奶的需求逐年增加,而人均口粮消费在减少,从而导致饲料用粮增加。另一方面,伴随着高血压、高血脂和糖尿病等疾病的发生,根据“药食同源”的概念,功能稻作为一种营养保健食品的需求量在增加。此外,伴随着我国乡村旅游和休闲农业的发展,彩色稻逐渐成为一种发展农村文化创意,拓展农业多功能发展的特用稻。本文综述了水稻作为特种稻方面的研究进展及部分调节机制,以期为我国水稻产业多样化发展提供参考信息。
1 饲用稻
通常情况下有两种饲用稻,一种是栽培品种作为食用稻收获,仅谷物作为精饲料用于喂养家禽和动物,另一种是全作物青贮稻,其栽培品种在黄熟期收获,包括穗、叶和茎的全稻植物被加工成青贮饲料,用于饲喂养牛。到目前为止,将水稻作为饲料利用的国家和地区较多,但是利用率普遍比较低,大多数都是直接进行秸秆还田,且没有形成规模。20 世纪90 年代初,水稻栽培专家青先国率先提出饲料稻研究开发,随后,在1994 年和1996 年国家将其列为重点攻关计划。通过对饲料稻的选育和饲用价值进行研究,得到了一批具有“三高”特点的优质饲用稻,如威优56、高蛋白食饲两用稻湘早籼24 号、金早47 等,同时这些品种都得到了国家或当地部门的推广与应用。在“十一五”期间,卢永根院士提出对全株饲料稻的筛选和加工进行研究。野生稻相对于普通栽培稻,具有栽培稻已经消失或不具备的优良特性,其抗病虫性较强、蛋白含量较高、生物量大及具有广亲和性,因此将野生稻开发成全株饲料稻有很大的前景。对14 个野生稻品种进行分析,发现部分野生稻的粗蛋白含量、分蘖能力高于现栽培的饲料稻,干物质量接近或者大于现栽培的饲料稻;此外,野生稻在某个营养指标(如蛋白含量)具有极强的优势,可以作为饲料稻改良的优质种质材料[2]。
秸秆具有纤维素含量低、半纤维素含量高等优势,同时还具有蛋白质、纤维素和糖等成分,而饲喂试验表明,利用脆秆稻草进行湖羊的饲养,可以增加羊群的采食量[3],因此,脆秆育种成为当下的一个新的育种内容。1963 年第1 个脆秆性状基因bc1被发现,截至目前,已鉴定出的脆秆基因有bc1、bc3、bc7等20 多个,通过作用于不同的细胞器从而进一步导致细胞壁纤维素合成酶的变异,最终导致秸秆的机械强度发生改变(图1)。BC3/DRP2B编码一个发动蛋白OsDRP2B,其突变后会降低水稻茎秆的机械强度;BC10突变后会增加水稻茎秆和叶片的脆性;在水稻突变体中,由于质膜上的OsCESA 蛋白数量减少,造成植株矮化,薄壁细胞塌陷,从而导致纤维素含量降低;BC14突变后会降低茎秆的机械强度;BC15突变后会减少细胞壁中的纤维素含量;OsCESA4编码纤维素合成酶中的催化亚基,其突变后会产生bc7和bc11两种不同突变位点的突变体,后代都表现出茎秆纤维素含量下降[4]。2008 年中国水稻研究所采用复合杂交技术培育出印水型水稻不育系中脆A,被鉴定为可以广泛用于选配两系脆秆杂交稻新组合[5]。中国科学院合肥物质科学研究院智能机械研究所离子束生物工程与绿色农业研究中心利用重离子诱变得到了一个半显性水稻脆秆突变体sdbc1,其表型表现出茎秆和叶片都较脆,2018年科辐粳7 号成为首个通过审定的脆秆品种,2019年进行示范推广,利用理想脆秆基因IBC培育出了水稻新品种科脆粳1 号,其半纤维素含量相对于对照增加15.7%[6]。利用脆秆137 为底盘材料,与连粳7 号、徐稻3 号杂交,采用回交的方式导入脆秆基因,从而得到两个脆秆新品系CG16-3 和CG16-9[7]。
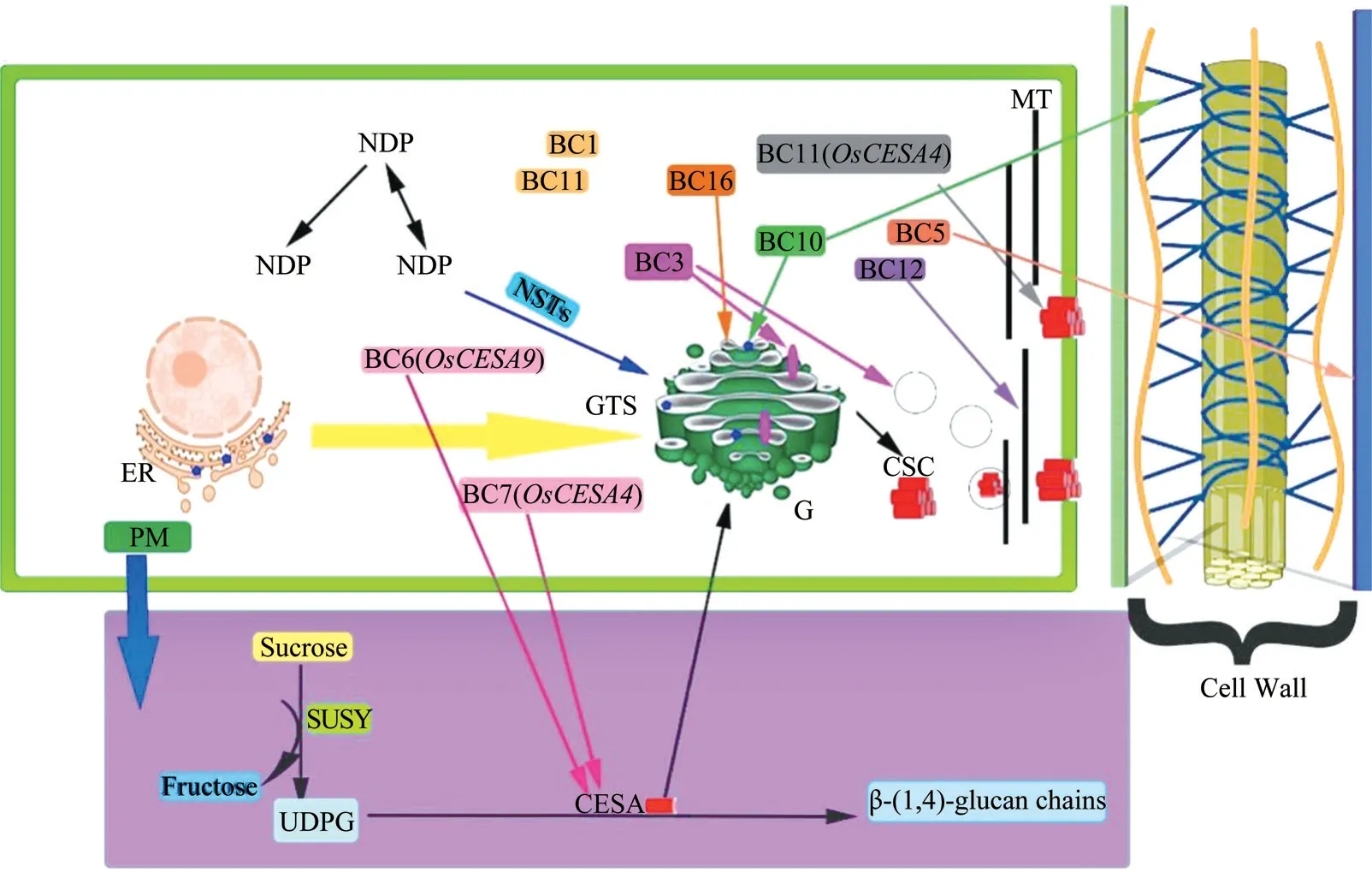
图1 细胞壁结构及其合成过程
2 功能稻
2.1 抗性淀粉类抗性淀粉(RS,Resistant starch)指的是在健康个体小肠中未被吸收的淀粉和降解产物的总和,具有调节人体血糖水平,促进矿物质吸收,保护肠道及调控体重等作用,其含量会受到环境与基因型的影响[8]。根据来源和性质,抗性淀粉可以分为5 种,分别为物理包埋淀粉RS1、天然抗性淀粉RS2、回生淀粉RS3、化学改性淀粉RS4和淀粉-脂质复合物RS5。在水稻中,淀粉的生物合成代谢途径比较明确,水稻籽粒中的淀粉含量为70%~80%,通过对水稻籽粒淀粉合成途径进行归纳分析可以明确抗性淀粉在淀粉合成途径中的位置,从而为增加水稻籽粒中抗性淀粉含量提供重要的理论依据(图2)。蜡质基因Wx和RS 之间是显著相关的,为调控RS 的主效基因,采用全基因组关联分析的方法进一步得到了与淀粉合成相关的Wx、SSIIa、ISA1和AGPS1基因都与RS 的含量有关,而抗性淀粉含量由少数主效基因和多个微效基因及非等位基因间的相互作用来调控[9]。通过对OsSBE3进行基因编辑,得到了抗性淀粉含量超过10%的水稻材料,而当OsSBE3基因发生单碱基突变时也会提高RS 含量[10]。研究表明,淀粉合成酶SSIIa 能够调节抗性淀粉中RSm 的含量,OsSSIIIa基因通过调节AGpase 和PPDK 的活性来提高胚乳中抗性淀粉的含量,在水稻突变体OsSSIIIa中,而WxaSSIIa能够显著提高抗性淀粉的含量[11]。杨瑞芳等[12]指出利用航天搭载诱变获得抗性淀粉突变体RS111,优化蒸煮方法,米饭中的抗性淀粉含量可以达到10%;通过辐射诱变获得了高抗性淀粉突变体浙辐201,其抗性淀粉含量为3.6%;以高抗性淀粉11JD64(降糖稻1 号)为母本、沪稻55 为父本杂交选育而成的优糖稻3 号,抗性淀粉含量为13.2%,是普通大米的20 倍以上;将ae突变体与IR36 杂交选育得到的高抗性淀粉水稻AE,抗性淀粉含量为8.25%;以R7954与粳稻突变体RS102杂交,得到糖尿病患者专用的水稻品种宜糖米1 号,蒸煮后其抗性淀粉含量为10.17%。此外,如宁农黑粳、功米3号、优糖稻2 号、降糖稻1 号等也富含抗性淀粉。
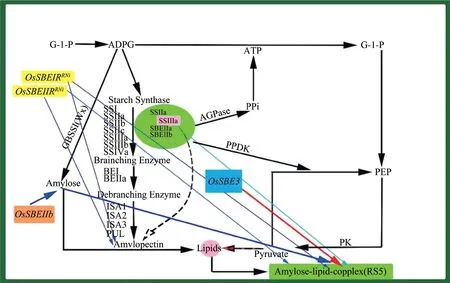
图2 抗性淀粉生物合成途径
2.2 富含微量元素类据报道,在稻米皮层中含有较多的人们所需要的微量元素,如铁、硒、锌。其中铁可以预防贫血症,硒具有保护心血管、预防肿瘤的功效,而锌具有提高免疫力的作用[13]。
由苏资2 号和螃蟹谷杂交选育而成的龙睛4号经美国农业部儿童营养研究中心和中国食品工业产品质量检测中心分析,富含铁、硒等微量元素和人体必需氨基酸。利用云南紫糯和(75069×香糯7-2)复合杂交选育而成的香血糯新品种,经研究表明其微量元素铁为普通糯米的3 倍,锌元素的含量也高于普通糯米[14]。1986 年上海农学院从早季晚熟、感温性籼稻品种驯化得到的乌贡1 号,其Fe、Zn 元素含量明显高于普通大米[15]。2016 年宁夏大学农学院对102 份水稻种质资源材料进行了Zn 含量的测定,发现杨和白皮稻、大粒稻、宁大62 和常规稻-5 等籽粒Zn 含量较高,通过方差分析发现Zn含量与籽粒的大小、长宽比和千粒重呈正相关[16]。
2.3 花青素、γ-氨基丁酸类花青素又称为花色素,广泛分布在各种植物体内,是一种黄酮类的次生代谢物。研究表明紫米、黑米和红米富含花青-3-葡萄糖苷及生物碱物质,能够抵抗癌症、心血管疾病,起到延缓衰老、养肝补血等作用[17]。
了解花青素的生物合成途径,对了解花青素在籽粒中的分布情况有重要作用。花青素和原花青素作为类黄酮合成途径中的产物,需要一系列酶的参与,同时,对与花青素相关的基因的功能机制也比较明确(图3)。研究表明,PAL、CHS、CHI、F3H、DFR和ANS可以催化花青素的合成,ANR和LAR催化形成原花青素,CHI突变导致水稻颖壳颜色呈现深棕色,总黄酮的含量提高,超表达OsANS基因,能够提高水稻品种的黄酮含量及增强抗氧化能力[18]。bHLH 家族转录因子OsB1 和OsB2 需要与MYB 转录因子结合才能参与水稻中花青素的调控,OsB2 可以与OsCHS的启动子结合,激活OsCHS基因的表达。将OsC1、OsB2和OsPAC1导入白米粳稻品种Chao2 和中花11 中超量表达,Chao2 和中花11 中超表达OsC1后,花青素代谢基因表达上调;OsPAC1超表达后,能够使OsCHS、OsCHI、OSANS和OsUFGT表达上调;OsB2在2 个品种中超表达后,OsCHS、OsCHI、OsANS、OsUFGT和OsF3’H表达量上调[19]。对93-11(白米)、SB7(红米)、IR1552(紫叶)和豫南黑籼糯(黑米)4 种水稻材料研究发现,3 个基本表达基因OsPAL、OsCHS和OsCHI在叶片、花期和花后1~2 周表达;而OsANS在黑米和紫米中表达[20]。将彩叶草中的CHS、CHI、F3H、DFR和ANS以及玉米转录因子中的Lc和PI导入同一载体,利用水稻胚乳特异性启动子转化水稻,得到富含花青素的紫晶米[21]。此外,鸭血糯、籼型香糯黑米闽紫香1 号、宁农黑粳和闽紫香2 号也是富含花青素的水稻品种。
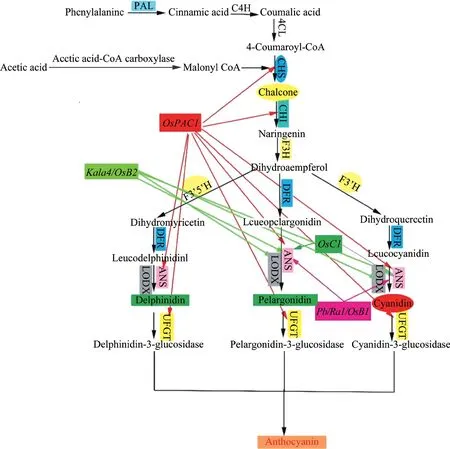
图3 花青素的生物合成途径
γ-氨基丁酸(GABA,γ-Aminobutiric acid)由谷氨酸脱酸酶脱羧形成的一种四碳非蛋白组成性氨基酸。研究表明,国内的水稻品种GABA 的含量大多在6~8mg/100g,GABA 在胚中的含量较高,而在主食大米中含量很低,胚芽中的蛋白质分解酶催化谷氨酸转化为GABA,在谷氨酸脱羧酶的作用下合成GABA。GABA 天然存在,具有治疗癫痫和帕金森病,降血压、防止动脉硬化、调节心律失常、抗疲劳和衰老等作用[22]。
在水稻中,以谷氨酸为前体物质,经过谷氨酸脱羧酶和谷氨酸脱氢酶的联合作用转化为γ-氨基丁酸,此外,一部分GABA 通过多胺衍生途径和GABA 分解作用,以鸟氨酸作为前体物质进行合成(图4)。方慧敏等[23]指出利用RNA 干扰技术,构建OsGABA-T1RNi载体,导入宁粳1 号中,降低OsGABA-T1的表达,导致糙米和精米中GABA 的含量相对于宁粳1 号增加;利用N-甲基-N 亚硝基对金南风进行诱变处理,从而得到了巨大胚突变体ge,相对于原品种胚增加2~3 倍。胡时开等[24]指出利用95-20 为父本、巨胚稻花晴M-2-565-11-3-B 为母本选育出GABA 含量为7.8mg/100g 的巨胚粳1 号;对高产糯稻恢复系嘉糯恢2 号采用Co60辐射得到的福巨糯2 号与巨胚不育系TgeA、1813geA 进行组配,获得了富含GABA 的巨胚杂交稻组合T 优巨胚2 号和1813 优巨胚2 号;以黑帅为母本,与巨胚1 号/早25 杂交,得到GABA 含量较高并且具有降压功能的黑珍珠[25]。王迎超等[26]提出巨胚稻中的γ-氨基丁酸含量高于常规稻米。通过对黑龙江7 个粳稻品种的研究,筛选出了肯糯2 以及龙粳31 两个富含γ-氨基丁酸的品种[27]。此外,富含GABA 的水稻品系还有Haiminori、龙特浦A 和龙特浦B 的突变体、巨胚一号、伽马-1、奥羽359、巨胚香糯1547、垦糯2号、6601、白巨胚米1575、MhgeR 和西巨胚1 号等。
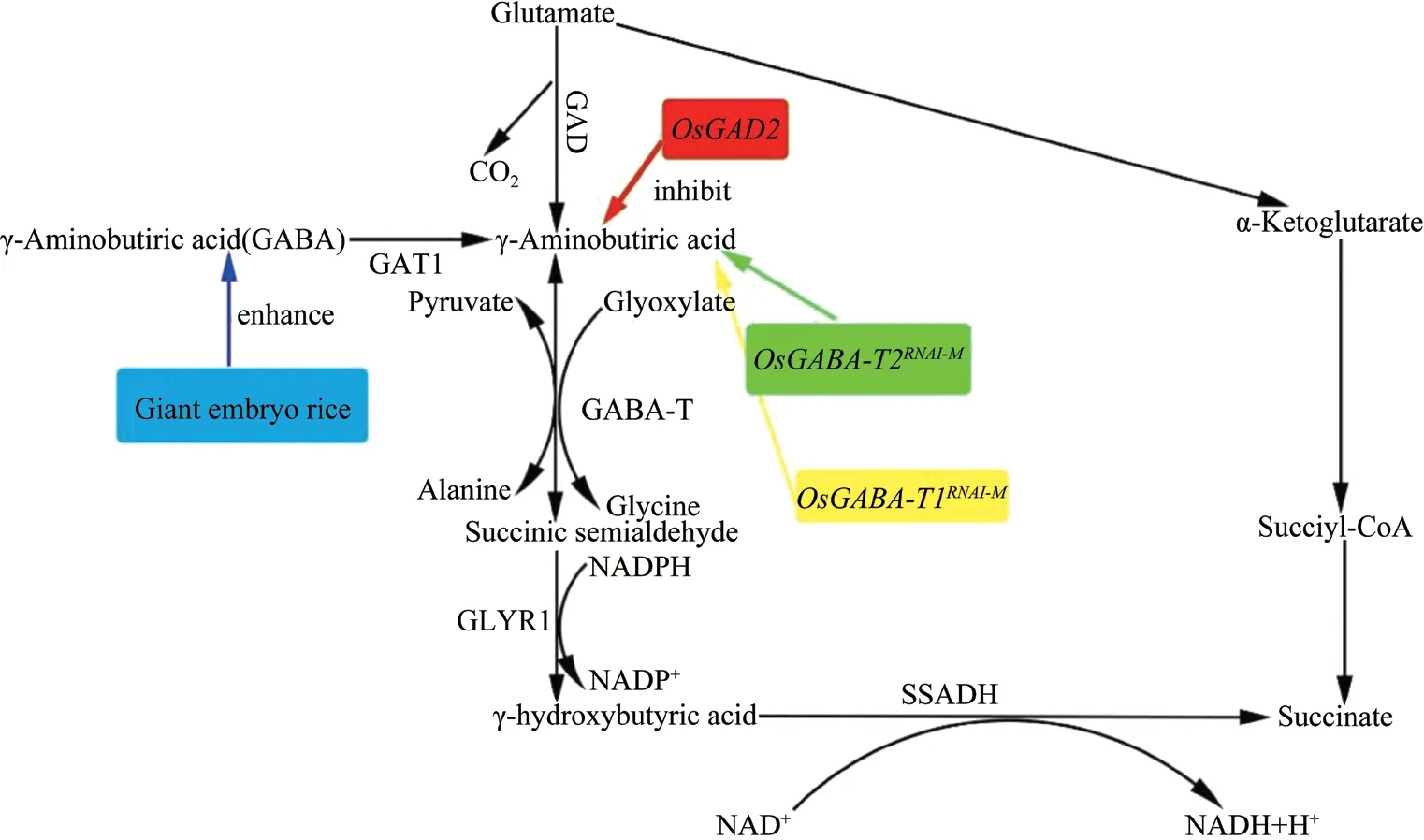
图4 γ-氨基丁酸的生物合成途径
2.4 低谷蛋白类根据溶解度的不同将蛋白分为水溶性的白蛋白、盐溶性的球蛋白、碱溶性的谷蛋白和醇溶性的醇溶蛋白。据报道,2017 年全球大约120 万人死于慢性肾脏疾病,该类疾病主要是由于尿蛋白的不平衡导致,低蛋白饮食被证实是除了药物以外能够有效控制尿蛋白水平的重要方法[28]。因此,发展和培育低谷蛋白水稻新品种是当下水稻育种的一个新目标。
目前,水稻谷蛋白基因多数已经得到克隆,按照其序列的相似性分为GluA 和GluB 两类,根据低谷蛋白突变体材料NM67 育成的Lgc-1,揭示了GluB 亚家族基因GluB4和被剪切了部分片段的GluB5基因以尾对尾反向重复形成的结构从而导致Lgc-1的低谷蛋白特性[29]。张莉萍等[30]提到日本学者利用化学诱变得到了谷蛋白含量下降4%的LGC-1;南京农业大学通过将低谷蛋白基因导入部分主栽品种,培育出了W3660、W204 等低谷蛋白含量增加的新品种;以超级粳稻宁粳4 号为母本、W1721 为父本,选育出产量较高、农艺性状较好的低谷蛋白新品种W0868;利用分子辅助与常规技术相结合的方法,将低谷蛋白基因LGC-1导入五山丝苗,得到了籼型低谷蛋白新品种N198,以及利用武育粳3 号与LGC-1进行杂交,后代利用分子标记辅助选择及SDS-PAGE 验证,得到了农艺性状优良的低谷蛋白粳稻新品种。此外,培育出的低谷蛋白水稻品种还有低谷1 号、武2812、春阳、LGC-jun等。
2.5 富含其他营养元素目前,随着基因工程的快速发展,研究人员利用转基因技术和CRISPR/CaS9基因编辑技术进一步改善和提高水稻中其他一些元素含量。据报道,先正达利用玉米种鉴定的八氢番茄红素合成酶(psy)与产生黄金大米的欧文氏菌胡萝卜素去饱和酶(crtl)结合,培育出胡萝卜素含量高达37μg/g 的第2 代“黄金大米”[31]。维生素B9(叶酸)被证实能够治疗贫血,降低心血管疾病等作用,用基因工程的方法将拟南芥ADCS和GCH1基因连接到水稻胚乳特异性启动子gluB1和glb-1,会提高水稻中叶酸的含量(1723μg/100g),为对照的100 倍[32]。将小麦HPPK/DHPS导入水稻,发现水稻叶酸含量会提高75%[33]。此外,利用基因工程技术选育出了能够促进大脑发育、预防心肌梗死和脑梗死的富含α-亚麻酸的水稻品种宁粳1-FAD3,以及能够降低血糖和血脂的富含降血糖短肽的龙稻5-mGLP 品种。
3 观赏稻
观赏植物作为重要的栽培植物,在社会经济发展中具有重要的作用,是21 世纪的“朝阳产业”。2007 年赵则胜在《特种稻研究与利用》中首次提出观赏稻。而彩色水稻作为特有的品种,被越来越多的人所了解,其不仅具有观赏作用,同时也可以食用,最终可以促进农业的发展和农民的增收。
对于观赏稻,常见的颜色主要有紫叶、黄叶和白色条纹叶稻等。日本是开展观赏稻育种较早的国家,其经常用观赏水稻品种的紫穗作为紫花的替代品。在国内,西昌学院戴红燕研究员对观赏稻的研究较多,利用日本稻縞稻为母本、Labell 为父本,经过有目的性的多代选择,培育出性状较为稳定的白拂;以縞稻为母本、北谷4 作父本,以其杂交后代中紫穗单株作为母本与日本稻C239 杂交,后代连续选择,得到新的观赏稻绿叶红妆;此外,利用日本稻红浪漫、阿久根糯、C239、北谷3 赤穗、水紫、胡枝香、对馬A46、PLW、縞稻相邻种植,使其自然杂交,后代定向选择红芒和黑芒的植株,得到新的观赏稻彩云和紫薇[34]。2019 年江西省农业科学院水稻研究所利用矮秆丛生突变体与受体亲本杂交,后代利用分子标记辅助选择,得到了一种盆景观赏稻培育方法[35]。福建省农业科学院水稻研究所利用抗病矮秆绿色叶水稻品种作为轮回亲本,分别与不同颜色、株型的品种进行杂交,后代与轮回亲本多代回交,从而得到一种新的观赏彩色稻品种的选育方法[36]。
4 问题与展望
目前,伴随着人民生活水平的提高,大多数育种工作者的研究方向主要以优质、高产和抗病作为主要目标,对于饲料稻、功能稻和观赏稻的关注较少,从而导致特种稻的发展较为缓慢。通过对一些国内外特种稻的发展形式进行分析,明确了当前特种稻发展急需解决的问题。首先,对于饲料稻,大多数的水稻种植者主要收获稻谷,而秸秆主要适用于还田,或是在收割之后堆积起来进行焚烧,或是被当作垃圾处理掉,导致秸秆利用率较低;对于脆秆水稻,近年来的研究虽然较多,有许多基因被克隆(如bc1、bc12等),但是对于脆秆相关的生物合成代谢途径还相对比较薄弱。其次,对于功能稻米,虽然市场上其售价比较高,但我国现有的功能稻品种较少,且缺少功能稻米品牌。此外,消费者对于功能稻米的认识程度也远远不足,国内也缺乏可以参照的功能稻米及其制品的行业标准。对于观赏稻,国内的种质资源比较短缺,大多数是通过引进的品种作为底盘材料进行改良,研究的内容比较单一。此外,缺乏配套的栽培技术,用途也比较单一。
基于上述问题,未来特种稻的发展方向应该重点发展以下几个方面:(1)加强对特种稻品种的选育,结合当代全基因组育种来进一步解析特种稻功能成分的生物合成代谢途径;(2)加强对特种稻的推广,使其达到产业化,从而提高消费者对于特种稻的认识度和接受度;(3)改变部分特种稻的育种方式,如目前花卉市场比较丰富,因此通过对观赏稻的改良,将其作为花卉进行盆栽;(4)加大特种稻种质资源的创新力度,完善特种稻的审定标准。
