橘小实蝇对甲氨基阿维菌素苯甲酸盐的抗性机制及对其他药剂的交互抗性
2023-02-03王新溪王圣印
王 敏, 王新溪, 王圣印
(1.浙江省金华市永康市农业技术推广中心, 金华 321300; 2.浙江农林大学现代农学院, 杭州 311300)
橘小实蝇Bactroceradorsalis又名橘子蛆、果蛆、黄苍蝇、柑橘小实蝇和东方果实蝇等,广泛分布于中国华东、华南、华中和云贵地区[1-2]。橘小实蝇寄主范围较广,目前发现其寄主包括花卉、水果、蔬菜共76个科211个属627种,适宜寄主种类数量高达478种[3]。橘小实蝇成虫羽化后需经历长时间补充营养,卵巢发育成熟后才能交配[4], 其雌成虫使用外生殖器刺破柑橘果皮,在果皮内产卵,每处产卵数量5~10粒不等[5]。卵期1~6 d,孵化后幼虫在果肉内为害,常导致柑橘被害果提前脱落,果肉腐烂失去经济价值[6-7]。目前橘小实蝇是重要的危险性国际检疫害虫,也是我国的检疫对象[8-9]。
橘小实蝇形态特征、分类地位、生活习性、发生规律和综合防治措施是目前的主要研究内容,由于杀虫剂广泛使用,关于橘小实蝇田间种群抗性的研究报道也逐渐增加[10-13]。橘小实蝇在我国华南、华中、华东和云贵地区为害猖獗,导致上述地区橘小实蝇抗药性发展迅速。陈朗杰等的研究发现,在深圳周边地区,橘小实蝇对生物源杀虫剂阿维菌素和菊酯类杀虫剂氯氟氰菊酯和高效氯氰菊酯产生了16.59~18.95倍的中等水平抗性,对广谱性杀虫剂敌百虫、辛硫磷、毒死蜱与灭多威产生了低水平抗性,但对生物源杀虫剂甲氨基阿维菌素苯甲酸盐(以下简称甲维盐)和多杀霉素仍处于敏感状态[14]。何凤梅等的研究发现,广西不同地区橘小实蝇抗性发展水平差异较大,玉林和百色等4个地市橘小实蝇田间种群均对氯氰菊酯产生了中等水平抗性,对阿维菌素形成中等抗性的橘小实蝇仅发现于桂林和玉林两市[15]。为探索高频使用甲维盐防治橘小实蝇是否存在抗性风险,通过室内连续筛选33代获得了橘小实蝇抗甲维盐种群(43.4倍),测定了抗甲维盐橘小实蝇种群与常用菊酯类杀虫剂高效氯氟氰菊酯、新烟碱类杀虫剂吡虫啉和噻虫胺、有机磷类杀虫剂辛硫磷和毒死蜱、生物源杀虫剂阿维菌素和多杀霉素、新型杂环杀虫剂虫螨腈的交互抗性水平,并探索了橘小实蝇对甲维盐的生化抗性机制。
1 材料与方法
1.1 供试杀虫剂和试剂
供试杀虫剂均为原药:99%甲维盐、99%阿维菌素和98%吡虫啉购于贵州利兴药业有限公司;99%辛硫磷、99%毒死蜱、99%噻虫胺、99%高效氯氟氰菊酯和99%多杀霉素购于合肥诺尔达生物科技有限公司;98%虫螨腈购于陕西惠诚生物科技有限公司。
主要试剂:98%毒扁豆碱、98%乙二胺四乙酸(EDTA)、98%牛血清白蛋白(BSA)与99%十二烷基磺酸钠(SDS)购于Sigma公司;70%考马斯亮蓝G250与99%还原型谷胱甘肽(GSH)购于Fluka公司;96% α-乙酸萘酯(α-NA)购于Aladdin公司。增效剂95%胡椒基丁醚(PBO)购于湖北汉达飞生物科技有限公司;98%磷酸三苯酯(TPP)购于山东国化化学有限公司;97%顺丁烯二酸二乙酯(DEM)购于上海昆淼实业有限公司。其他试剂为国产分析纯或化学纯。
1.2 供试虫源
相对敏感种群(S):采自浙江省金华市武义县履坦镇汤村。在果园内采集带虫柑橘落果,带回实验室内以柑橘饲喂。将虫果放置在铺有5 cm河沙的塑料盆中,果内老熟幼虫脱果进入河沙内化蛹。收集橘小实蝇成虫放置于20目纱笼内(60 cm×60 cm×60 cm),为其提供蜂蜜水(3%)补充营养,使用柑橘诱集产卵。室内饲养10代,期间不接触任何杀虫剂,建立相对敏感种群,收集2日龄成虫作为供试虫源。
金华田间种群(JH):采自浙江省金华市永康市花街镇八字墙村,虫果带回室内后参照敏感种群饲养。收集F2代的2日龄成虫供试。
甲维盐抗性种群(EB):将甲维盐原药使用丙酮配制成母液,根据橘小实蝇抗性发展速度,按照每次筛选杀死约50%成虫的剂量在蜂蜜水中加入甲维盐母液。收集敏感种群2日龄成虫约1 000头作为供试虫源封闭于纱笼内,饥饿处理2 h后,在纱笼内放入甲维盐蜂蜜水,橘小实蝇取食1 h后取出甲维盐蜂蜜水,使用无杀虫剂的柑橘继续饲养。筛选33代后,抗性种群的抗性倍数为43.4倍。收集抗性种群2日龄成虫供试。饲养环境条件为(25±0.5)℃,光周期L∥D=14 h∥10 h,RH(65±5)%。
1.3 生物测定
橘小实蝇生物测定采用玻璃管药膜法[10,14]。将供试杀虫剂原药使用丙酮配制成母液,根据预试验测定结果,将杀虫剂母液用丙酮稀释成5~7个浓度。抽取10 mL稀释后的药液,注入玻璃管中(直径5 cm,长15 cm),摇动玻璃管使药液均匀涂布,倒掉多余的药液,静置玻璃管至内壁药液风干。将20~25头橘小实蝇2日龄成虫引入涂布药膜的玻璃管,放置3%蜂蜜水浸透的棉球供成虫取食,用纱布封闭玻璃管口。24 h后观察玻璃管内橘小实蝇成虫的存活数量,以毛笔轻触虫体不能正常爬行作为标准判定为死亡。对照组为丙酮水溶液,有效试验中对照组死亡率必须低于10%。处理和对照组均重复3次。
增效剂生物测定:首先将增效剂胡椒基丁醚(PBO)、磷酸三苯酯(TPP)和顺丁烯二酸二乙酯(DEM)使用丙酮配制成母液(10 g/L)[16]。然后根据预试验结果,将供试杀虫剂稀释成5~8个浓度,药液体积除以100为增效剂母液的体积。将增效剂母液加入药液,可使增效剂在所有药液中的浓度一致(100 mg/L)。其他操作与上述生物测定方法相同。每浓度重复3次。
1.4 酶活性测定
1.4.1细胞色素P450含量测定
参考Lee等[17]的方法收集橘小实蝇2日龄雌成虫5头,置于匀浆缓冲液(0.1 mol/L pH 7.5磷酸钠缓冲液,含1 mmol/L EDTA、0.1 mmol/L DTT、1 mmol/L PTU、1 mmol/L PMSF和10%甘油)中匀浆,操作全程冰浴。4℃,10 000 g离心15 min。过滤上清液后,4℃,10 000 g下离心1 h,将微粒体沉淀在重悬浮缓冲液(0.1 mol/L pH 7.5磷酸钠缓冲液、1 mmol/L EDTA、0.1 mol/L DTT、1 mmol/L PMSF和20%甘油)中重悬浮作酶源。重复3次提取酶原。
参考Omura等[18]CO差示光谱法测定多功能氧化酶(MFO)和细胞色素P450含量。将微粒体悬浮液用重悬缓冲液稀释至1 mg/mL左右,加入连二亚硫酸钠颗粒至饱和,平衡5 min,一分为二,分别加入参比杯和样品杯中。使用紫外可见分光光度计在波长400~500 nm处扫描,以参比杯为空白,扫描样品杯的光吸收值为基线。待基线稳定平直时在样品杯中通CO,以参比杯为空白,扫描样品杯的光吸收值,记录 OD450、OD490、OD426和OD409,按下式分别计算P450和细胞色素b5含量。每次提取酶液重复测定3次。
P450含量=(OD450-OD490)/(消光系数×蛋白含量),其中消光系数为91 mmol·L-1·cm-1;
细胞色素b5含量=(OD426-OD409)/(消光系数×蛋白含量),其中消光系数为185 mmol·L-1·cm-1。
1.4.2细胞色素P450O-脱甲基酶活性测定
参考Hansen等[19]测定多功能氧化酶(MFO)O-脱甲基酶活性。反应总体系2 mL:1 270 μL 0.1 mol/L pH 7.8的磷酸缓冲液,200 μL 0.36 mmol/L NADPH,30 μL 3 mmol/L对-硝基苯甲醚,500 μL酶液。反应体系在25℃振荡温育反应30 min,加入0.5 mL 1 mol/L HCl终止反应,再加入2.5 mL三氯甲烷萃取反应产物对-硝基酚,3 000 g离心15 min。取下层即三氯甲烷层1.5 mL加入1.5 mL 0.5 mol/L NaOH反萃取。NaOH萃取液使用分光光度计在400 nm测定OD值,以0.5 mol/L NaOH溶液为对照。以对-硝基酚标准曲线和酶液中的蛋白质含量求出每分钟每毫克蛋白催化产生的对-硝基酚的摩尔数来表示酶的活性。重复3次提取酶液,每次提取酶液重复测定3次。
1.4.3羧酸酯酶活力测定
参照Van Asperen[20]的方法测定橘小实蝇CarE活力。取5头橘小实蝇2日龄雌成虫在液氮中冷冻后立刻加入0.04 mol/L磷酸缓冲液(pH 7.0)2 mL,在冰浴条件下迅速充分匀浆,匀浆液于4℃、10 000 r/min离心10 min,分离上清液即为酶液,冰浴待用。以5 mL α-乙酸萘酯(3×10-4mol/L)为底物,加入0.2 mL酶液,在37℃条件下恒温水浴振荡30 min,加入1 mL DBLS(1%坚固蓝B盐水溶液∶5%十二烷基硫酸钠水溶液=2∶5)终止反应,将混合液置于室温显色30 min,使用分光光度计测定600 nm处OD值。对照组以0.2 mL磷酸缓冲液代替酶液。根据α-萘酚标准曲线计算出每毫升酶生成的α-萘酚量。测定酶源蛋白质含量(mg·mL-1),计算出羧酸酯酶的比活力(nmol·min-1·mg-1)。重复3次提取酶液,每次提取酶液重复测定3次。
1.4.4谷胱甘肽S-转移酶活力测定
参照Wu等[21]的方法测定GST的比活力。取5头橘小实蝇2日龄雌成虫在液氮中冷冻后立刻加入0.1 mol/L磷酸缓冲液(pH 7.0)2 mL在冰浴条件下迅速充分匀浆,匀浆液于4℃、10 000 r/min离心10 min,分离上清液即为酶液,冰浴待用。以CDNB为底物,在试管中依次加入66 mmol/L磷酸缓冲液2.4 mL,50 mmol/L谷胱甘肽0.3 mL,0.03 mol/L CDNB 0.1 mL,酶液0.2 mL,立即混匀,在27℃水浴条件下反应5 min,使用分光光度计在340 nm下测OD值。对照组以0.2 mL磷酸缓冲液代替酶液。酶源经蛋白质含量测定,计算出谷胱甘肽S-转移酶的比活力(nmol·min-1·mg-1)。重复3次提取酶液,每次提取酶液重复测定3次。
1.4.5蛋白总量测定
酶源蛋白质含量采用考马斯亮蓝G-250染色后测定[22]。吸取0.3 mL酶液于10 mL试管中,空白对照以0.3 mL磷酸缓冲液代替,加入5 mL考马斯亮蓝G-250试剂,充分混匀于25℃恒温水浴染色2 min,水浴后5 min内在595 nm下比色测OD值。根据牛血清白蛋白标准曲线计算出蛋白质含量,除以0.3即每毫升酶液中的蛋白含量(mg·mL-1)。重复3次提取酶液,每次提取酶液重复测定3次。
1.5 数据处理
橘小实蝇交互抗性与增效剂生物测定数据使用Polo Plus 1.0软件分析,计算毒力回归曲线斜率、致死中浓度(LC50)和95%置信区间[23]。酶活性测定数据使用SPSS 22.0软件进行分析,用Duncan氏新复极差法进行差异显著性分析。
2 结果与分析
2.1 交互抗性
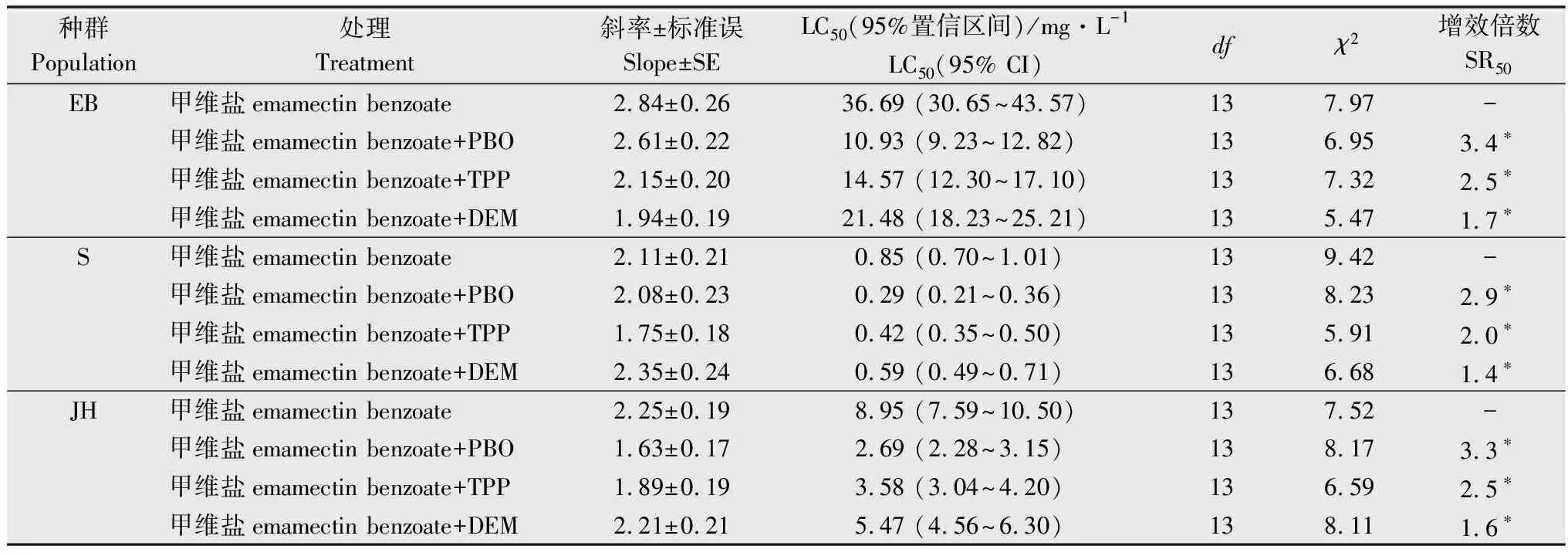
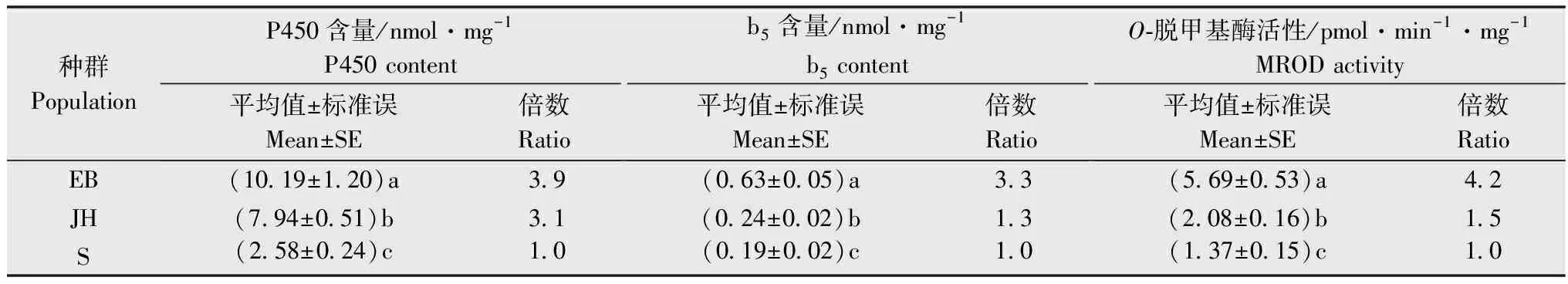
橘小实蝇EB种群与阿维菌素存在中等水平交互抗性(RR50=20.7),与吡虫啉、辛硫磷和马拉硫磷存在低水平交互抗性(5 表1 橘小实蝇抗甲维盐种群对8种杀虫剂的交互抗性水平1) 增效剂生物测定结果表明(表2), PBO对橘小实蝇EB种群、S种群和JH种群均有显著增效作用,增效倍数分别为3.4、2.9倍和3.3倍; TPP对橘小实蝇EB种群、S群和JH种群均有显著增效作用,增效倍数分别为2.5、2.0倍和2.5倍; DEM对橘小实蝇EB种群、S种群和JH种群也具有显著增效功能,增效倍数分别为1.7、1.4倍和1.6倍。 表2 增效剂PBO,TPP和DEM对甲维盐的增效作用1) 酶活性测定结果表明(表3),橘小实蝇EB种群和JH种群的细胞色素P450(3.9倍和3.1倍)和b5含量(3.3倍和1.3倍)、O-脱甲基酶活性(4.2倍和1.5倍)均较敏感种群S交显著提高。 表3 橘小实蝇不同种群MFO O-脱甲基酶活性、细胞色素P450和b5含量1) 橘小实蝇EB种群和JH种群的谷胱甘肽S-转移酶活性(2.7倍和1.4倍)和羧酸酯酶活性(3.2倍和1.9倍)均显著高于敏感种群(表4)。 表4 橘小实蝇不同种群谷胱甘肽S-转移酶和羧酸酯酶活性1) 研究发现,棉铃虫Helicoverpaarmigera、西花蓟马Frankliniellaoccidentalis、黏虫Mythimnaseparata等农业害虫经人工选育对甲维盐产生中高水平抗性之后,常对多种杀虫剂产生不同水平的交互抗性[24-26]。人工室内选育出的西花蓟马抗甲维盐种群可对阿维菌素产生高水平交互抗性(31.7倍),对甲维盐具有高抗性水平的黏虫种群也会对阿维菌素形成中等交互抗性(21.8倍)[25]。本研究发现,橘小实蝇EB种群与阿维菌素存在中等水平交互抗性。综合黏虫、西花蓟马和棉铃虫交互抗性研究结果推测,同种害虫易对作用机制相同的杀虫剂产生交互抗性。对甲维盐具有高抗性水平的西花蓟马种群可对啶虫脒形成中等交互抗性,对吡虫啉、虫螨腈、氯氟氰菊酯、毒死蜱和灭多威有低水平交互抗性;黏虫高抗甲维盐种群可对毒死蜱和灭多威形成中等交互抗性,与辛硫磷和氟氯氰菊酯之间交互抗性水平较低;西花蓟马和黏虫抗性种群的多功能氧化酶、羧酸酯酶和谷胱甘肽S-转移酶等生化解毒酶活性均显著增强[25-26]。在本研究中,橘小实蝇EB种群对吡虫啉、马拉硫磷、辛硫磷、高效氯氟氰菊酯、噻虫胺表现出低水平交互抗性,推测橘小实蝇EB种群代谢解毒酶活性升高可能是形成上述低水平交互抗性的重要生化机制。橘小实蝇EB种群对噻虫胺、高效氯氟氰菊酯、多杀霉素和虫螨腈未表现出交互抗性,因此推荐甲维盐与噻虫胺、高效氯氟氰菊酯、多杀霉素和虫螨腈在田间轮换使用,对于甲维盐与吡虫啉、辛硫磷和马拉硫磷轮用应持谨慎态度,禁止甲维盐与阿维菌素轮用。 多项研究表明,多种农林害虫对甲维盐的抗性与多功能氧化酶活性增强相关[27-29]。王圣印等[25]的研究发现,多功能氧化酶抑制剂增效醚(PBO)可显著降低甲维盐对西花蓟马的致死中浓度,对甲维盐具有高抗性水平的西花蓟马种群多功能氧化酶活性显著高于敏感种群。宋月芹等[26]的研究表明,PBO可显著提高甲维盐对黏虫的毒力,黏虫对甲维盐产生高水平抗性时,其多功能氧化酶活性亦同步显著上升。侍甜等[30]的研究表明,多功能氧化酶抑制剂增效醚(PBO)可显著提高甲维盐对甜菜夜蛾田间种群的毒力,酶活性测定结果也表明其多功能氧化酶活性比敏感种群高出1.7~7.4倍。在本研究中,PBO可显著提高甲维盐对橘小实蝇的室内毒力,橘小实蝇EB种群的O-脱甲基酶活性显著升高,细胞色素P450和b5含量也显著增高。由上述研究结果推测,多功能氧化酶活性提高可能是橘小实蝇经室内筛选对甲维盐产生高水平抗性的主要生化机制。 羧酸酯酶抑制剂磷酸三苯酯(TPP)常用于间接探索昆虫羧酸酯酶活性与抗药性的关系[31-33]。王圣印等[25]的研究发现,TPP可显著降低甲维盐对西花蓟马高抗甲维盐种群和敏感种群的致死中浓度,西花蓟马对甲维盐产生高水平抗性后,其羧酸酯酶活性也显著增高。对黏虫高抗甲维盐种群和敏感种群进行室内毒力测定时,TPP可显著降低甲维盐对上述两个种群的致死中浓度,抗甲维盐种群的羧酸酯酶活性也显著升高到1.64倍[26]。侍甜等[30]的研究表明,在使用甲维盐对甜菜夜蛾进行室内生物测定时,酯酶抑制剂DEF对甲维盐的增效倍数最高可达7.7倍,其羧酸酯酶活性可上升至敏感种群的4.7倍。本研究中,橘小实蝇EB种群的羧酸酯酶活性显著增强,而TPP可显著降低甲维盐对橘小实蝇EB、JH和S种群的致死中浓度。综上所述,羧酸酯酶活性增强可能是橘小实蝇对甲维盐抗性提高的重要因素。 在增效剂生物测定试验中,顺丁烯二酸二乙酯(DEM)常用于间接探索昆虫谷胱甘肽S-转移酶与其抗药性的关系[34-36]。谷胱甘肽S-转移酶活性抑制剂DEM可显著降低甲维盐对西花蓟马的致死中浓度,西花蓟马对甲维盐的抗性水平上升时,谷胱甘肽S-转移酶活性也随之显著提升[25]。DEM可显著提升甲维盐对黏虫的室内毒性,当黏虫对甲维盐产生高水平抗性时,抗性种群的谷胱甘肽S-转移酶活性显著上升[26]。本研究中,室内汰选得到的橘小实蝇EB种群谷胱甘肽S-转移酶活性显著提高,而DEM可显著提高甲维盐对EB、JH和S等3个橘小实蝇种群的室内生物测定毒力,因此,谷胱甘肽S-转移酶活性增强可能是橘小实蝇对甲维盐抗性水平上升的重要因素之一。 目前对橘小实蝇多进行综合防控,检验检疫是防治橘小实蝇的首要措施,预测预报有助于做好防治准备措施并决定防治时机,农业防治清洁田园可有效减少虫源,物理防治悬挂黄板或诱集液有助于防治橘小实蝇成虫[37]。在橘小实蝇成虫暴发期,化学防治也是必须采取的措施之一[37-38]。结合本研究结果,建议合理轮换使用甲维盐、噻虫胺、高效氯氟氰菊酯、虫螨腈和多杀霉素,可有效延长甲维盐、噻虫胺、高效氯氟氰菊酯、虫螨腈和多杀霉素的使用寿命。以甲维盐单剂为基础,通过添加增效剂创新杀虫剂配方与剂型,有助于实现对橘小实蝇抗性治理,提高甲维盐对橘小实蝇的田间防效。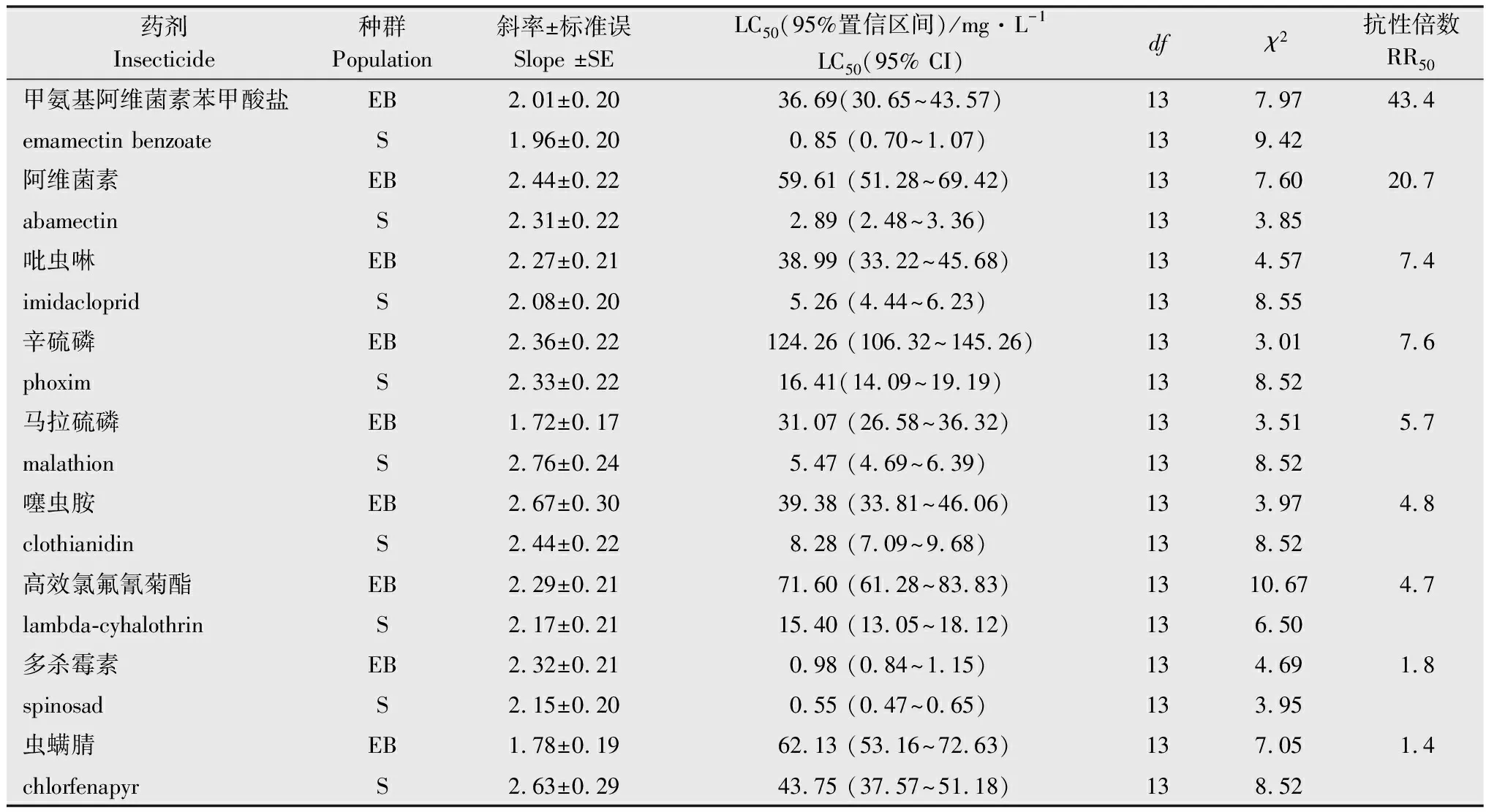
2.2 3种增效剂对甲维盐的增效作用
2.3 酶活性比较

3 讨论
