脱氧雪腐镰刀菌烯醇对猪毒害作用及毒理机制研究进展
2023-02-03戴超辉赵为民付言峰李碧侠王学敏程金花
戴超辉,李 辉,赵为民,付言峰,廖 超,李碧侠,王学敏,程金花
(江苏省农业科学院畜牧研究所,农业农村部种养结合重点实验室,江苏省农业种质资源保护与利用平台,南京 210014)
猪饲料主要由大豆、玉米、豆粕、稻糠、大麦、矿物质和饲料添加剂等组成,极易受到霉菌毒素的污染。有人员对全球猪饲料的污染情况进行了研究,结果检测的饲料样本中有96%的样本至少被一种霉菌毒素污染,其中被检测到最多的霉菌毒素主要是脱氧雪腐镰刀菌烯醇、伏马毒素和玉米赤霉烯酮,而猪是对脱氧雪腐镰刀菌烯醇最易感的动物之一[1-3]。脱氧雪腐镰刀菌烯醇(deoxynivalenol, DON)又名呕吐毒素(vomitoxin, VT),是由禾谷镰刀菌(Fusariumgraminearum)、尖孢镰刀菌(Fusariumoxysporum)、串珠镰刀菌(Fusariummoniliforme)、拟枝孢镰刀菌(Fusariumsporotrichioides)、粉红镰刀菌(Fusariumfoseum)和雪腐镰刀菌(Fusariumnivale)等产生的单端孢菌毒素[4-5]。DON主要存在于小麦、大麦、燕麦、玉米等谷类作物中,在低温条件下仍能保持其活性[6-7]。DON化学结构稳定,耐高温和强酸条件,在饲料加工过程中不易被破坏,因此DON污染已成为饲料质量安全的重大威胁之一[8]。DON对所有动物都有毒性作用,而猪对DON的敏感性最高[9-10]。近年来DON的发生率越来越高,严重损害了猪的健康和生长,给全球养猪业带来了巨大挑战[11]。本文主要综述了DON对猪的毒害作用、毒理机制以及DON解毒剂开发研究进展,尤其关注DON毒理作用的遗传调控研究,旨在为从遗传本质提高猪抵御脱氧雪腐镰刀菌烯醇毒害作用提供更全面的参考。
1 DON对猪的毒害作用
随着饲料工业和贮藏技术的发展,在饲料加工和储存等方面采用了大量防DON污染措施。尽管如此,猪饲料中DON的检测超标率仍很普遍[11]。猪感染DON主要是由于食用被DON污染的饲料,DON对猪的毒害作用主要体现在出现不良反应(拒食、呕吐、生长抑制等)、组织器官损伤(肝、脾、胃、肠道等)、细胞毒性(氧化应激、内质网应激、细胞凋亡和增殖受阻等)和免疫毒性(免疫刺激和免疫抑制)等方面(图1)。

图1 DON对猪的毒害作用(原始图片素材来自https://www.pigprogress.net/health-tool/)Fig.1 Toxic effects of DON on pigs (the original image comes from https://www.pigprogress.net/health-tool/)
1.1 DON引起猪不良反应
猪在采食DON污染的饲料后,可能会出现呕吐、腹泻等中毒症状,有时还可能站立不稳[12]。研究表明,饲料摄入DON会降低猪的采食量和增重,当饲喂含1~3 mg·kg-1的DON饲料时,猪的生长性能开始下降;每增加1 mg·kg-1的DON就会使猪的增重降低8%左右[13-14]。长期、低剂量食用被DON污染的饲料可导致采食量降低甚至拒食、体重增加减少和营养效率改变;急性、高剂量暴露于DON可能导致呕吐、腹泻、肠道炎症和胃肠道出血[12]。进一步研究发现,DON通过调节局部血清素和减少肠蠕动、增加饱腹感信号、促炎细胞因子的释放以及可能引起呕吐来减少猪的采食量[15-17]。此外,DON诱导的肠道屏障破坏和通过激活猪的丝裂原活化蛋白激酶(MAPK)途径增加肠道通透性也会导致生长抑制[18]。研究表明,饲料中10 mg·kg-1的DON会降低猪对必需氨基酸的消化率,导致能量和营养摄入量、营养吸收降低,以及细胞代谢受损,最终对猪的生长产生负面影响[18-19]。另外,4 000和8 000 ng·mL-1浓度DON会抑制猪空肠对丙氨酸和葡萄糖的吸收,表明DON对通过猪小肠营养物质的运输具有抑制作用[20]。怀孕母猪在采食含有DON的饲料后,机体对疾病的抵抗力下降,并且在所生胎儿的血清、肝和肾等器官中均能检测到DON[21]。
1.2 DON对猪组织器官和细胞的毒害作用
猪采食受DON污染的饲料,经消化道吸收后在肝中首先检测到DON,采食DON污染饲料组猪的肝重量比对照组要重[22];并且肝中血清白蛋白的含量显著降低,出现肝细胞组织紊乱;不论是急性还是慢性DON中毒,仔猪肝组织都会发生病变[23-24]。DON通过抑制肝的蛋白合成,易引发宿主氨基酸血症,进而对肝器官的生长造成影响[25]。另外,DON对未成熟的后备母猪肝组织学和超微结构也产生了影响,包括小叶周围结缔组织的厚度增加及其对小叶的渗透、总微观肝评分(组织学活动指数,HAI)增加、猪肝窦扩张、糖原含量变化、肝细胞中铁离子积累增加、猪肝细胞内质网组织的变化以及肝巨噬细胞形态的变化,也会间接诱导猪肝细胞凋亡[26-27]。DON还可能对猪造成其他一些不良影响,包括胃食管区域、肺和肾的病变,以及不同临床化学参数(血浆营养素和血浆酶活性)的变化[28]。另外,有研究报道DON导致原代猪脾淋巴细胞产生氧化损伤,从而抑制线粒体融合并促进线粒体自噬[29]。总之,DON可削弱肝、脾、肾、胃等组织器官的功能。
此外,有研究报道了低剂量DON对猪大肠组织结构和超微结构的影响,发现低剂量的DON不会影响大肠黏膜和黏膜下层的结构,不影响增殖细胞核抗原的表达和肠道屏障通透性,但显著减少了盲肠和降结肠中杯状细胞的数量,增加了大肠中淋巴细胞和浆细胞的数量,可能会影响大肠的防御机制[30]。但当给猪饲喂高剂量的DON后,可观察到其空肠绒毛萎缩、溶解,绒毛高度降低和肠道细胞增殖受到影响等现象[31-32]。并且,DON能够损伤肠道运输并影响营养物质的吸收[33]。DON对营养吸收的毒性作用包括抑制小肠刷状缘膜中钠葡萄糖共转运载体蛋白SGLT-1的活性,从而限制机体对关键营养素(例如葡萄糖)的吸收[34-35]。
关于DON对猪生殖系统的毒害作用,目前也有相关报道。Tiemann和Dänicke[28]研究指出,DON可抑制母猪子宫细胞增殖,阻碍卵母细胞和胚胎发育。孙文涛等[36]发现,含DON的污染饲料会引起断奶仔猪子宫器官指数极显著增加、子宫腺腺泡数量增多、密度增大、子宫内膜厚度极显著增加。另外,霉变玉米中的DON导致仔猪相对卵巢重量显著增加,并使仔猪卵巢组织抗氧化水平降低[37]。此外,DON可以诱导猪卵巢颗粒细胞(porcine granulosa cells, pGCs)的形态变化和凋亡,并抑制细胞增殖[38]。并且,DON能够导致公猪睾丸支持细胞结构破坏、细胞周期停滞和氧化应激[39]。
1.3 DON对猪的免疫毒性
免疫系统对DON敏感,DON会使机体免疫调节异常[40]。DON免疫毒性相关的MAPK信号通路激活的结果取决于剂量和持续时间:急性低剂量暴露会导致免疫刺激,其特征是细胞因子、趋化因子和其他促炎相关蛋白的上调,而长期高剂量暴露通常会导致细胞凋亡、细胞周期停滞和免疫抑制[12]。这种免疫功能失调是核毒性应激的结果,之所以会出现免疫激活和免疫抑制两种截然不同的影响,是因为DON激活了p38和ERK1/2两个丝裂原活化蛋白激酶相互竞争的信号通路,p38通路促进细胞凋亡,而ERK1/2通路有利于细胞存活和细胞因子的表达[41]。研究发现,低浓度的DON诱导T细胞和B细胞中促炎基因mRNA和蛋白表达;而高浓度的DON会导致白细胞迅速凋亡并伴随免疫抑制[42-43]。另外,DON会损害体液和细胞介导的免疫反应,导致体液免疫反应失调,使机体对疾病的抵抗力减弱;且DON会改变血清IgA水平,而IgA和免疫球蛋白肾炎息息相关[44]。DON还会对骨髓造血干细胞产生细胞毒性,当其作用于T、B淋巴细胞时,造成的免疫毒性作用会诱导细胞凋亡[45]。DON能够抑制蛋白质的合成和细胞增殖,减少巨噬细胞、淋巴细胞和红细胞的数量,抑制细胞合成[9]。长期饲喂DON污染饲料会导致猪只IL-8和GPX-2的表达增加,血清总免疫球蛋白A和特异性免疫球蛋白增加[46-47];并且DON能够降低T细胞增殖和参与T细胞活化的分子表达,导致免疫抑制[48]。此外,DON可以改变猪多形核细胞(polymorphonuclear cells, PMNC)的免疫功能,能够导致LPS诱导的IL-8分泌、趋化性和吞噬能力都发生变化,并引起PMNC发生凋亡[49]。
2 DON毒理机制研究进展
2.1 DON毒性作用机制
从化学角度看,DON分子中C12/C13(图2)上的环氧基团决定了其主要毒性,其毒性也与3个羟自由基有关[50]。从分子生物学角度看,DON有两种已知的致病机制:1)DON靶向核糖体,DON分子的环氧基与细胞的肽基转移酶中心(PTC)的A位结合,导致蛋白质合成链延伸步骤的延长,抑制DNA、RNA和蛋白质的合成;2)DON能够激活与增殖、分化和凋亡有关的(包括p38、JNK、ERK1/2等)信号转导细胞激酶,引起“核糖体毒性应激”效应,从而破坏正常细胞功能[40]。有研究报道,DON会通过使细胞周期停留在DNA合成期前期,抑制细胞进入DNA合成期,造成细胞休眠,使细胞增殖出现阻碍,最终导致细胞数量下降[51-52]。另外,DON可通过氧化应激介导细胞DNA损伤,造成基因毒性[53]。DON通过诱导核糖体应激反应改变细胞膜完整性,使细胞氧化还原状态和细胞信号传导发生改变,致使宿主机体各处细胞发生凋亡[12]。总之,DON已被证明可影响猪细胞中的信号通路、氧化应激、内质网应激、细胞凋亡和增殖,最终发挥毒性作用[54]。

图2 脱氧雪腐镰刀菌烯醇(DON)的化学结构示意图[64]Fig.2 Schematic diagram of the chemical structure of deoxynivalenol (DON)[64]
2.2 DON对猪肠道损害的机制研究
目前关于DON对猪肠道损害的机制研究比较多。一方面,DON能通过改变猪肠道的跨膜上皮电阻或经上皮的离子导电性,损伤猪肠道屏障功能、降低肠道细胞间常见离子传递和肠道细胞完整性[18]。另外,DON可以通过激活MAPK信号通路和改变负责猪肠道组织关键生理和免疫功能的基因表达产生毒性;MAPK信号级联的激活导致肠道屏障功能中断,并通过减少紧密连接蛋白的表达增加通透性[35]。Kang等[55]研究表明,DON以剂量依赖性方式诱导产生猪肠上皮细胞(IPEC-J2)的细胞毒性,增加了与炎症和细胞凋亡相关基因的表达,并降低细胞抗氧化状态,揭示了DON诱导的IPEC-J2细胞损伤机制与活性氧ROS产生增加、细胞凋亡和炎症相关的基因和蛋白质的表达有关。另外,虽然DON短时间处理不影响猪肠道屏障功能,但显著抑制了猪肠上皮细胞增殖和肠道干细胞扩增,其作用机制与ABC transporter、PI3K/Akt/m TOR信号通路、JAK/STAT信号通路和Wnt/β-catenin信号通路有关[56]。DON还会对IPEC-J2细胞中的炎症损伤核因子kappa-B(NF-κB)通路产生影响,当DON浓度增加时,NF-κB p65蛋白在细胞质和细胞核中的表达逐渐增加,一些炎症因子的转录上调,从而导致NF-κB相关基因的表达或细胞中的蛋白质发生改变[57]。另外,DON通过MAPK信号通路的磷酸化触发IPEC-J2细胞的炎症,主要表现在MAPK信号通路(p38、ERK和JNK)的磷酸化增加[58]。DON能够影响黏膜免疫反应,导致免疫抑制或刺激局部免疫细胞和细胞因子释放,并促进肠道病原体在肠道中的持久存在,这些免疫反应均会加剧猪肠道感染和局部炎症,使肠细胞和宿主更容易受到管腔毒性化合物的影响[35]。DON可以在长时间低剂量情况下使猪回肠中IL-1β和IL-8表达下调[59]。而且,DON可通过改变肠道IL-1β和IL-6的分布和表达进而影响肠道的免疫机能[60]。此外,DON可能会抑制Wnt/β-catenin通路,导致肠隐窝细胞的有丝分裂减少[61]。DON可以改变IPEC-J2细胞的形态,破坏其亚微观结构,增强细胞膜的通透性,降低跨上皮电阻,破坏肠道屏障[57, 62]。总之,DON能够改变肠道屏障,削弱免疫反应并增加对传染病的易感性[63]。笔者综合上述文献资料绘制了DON对肠道的毒害作用及其可能机制(图3)。
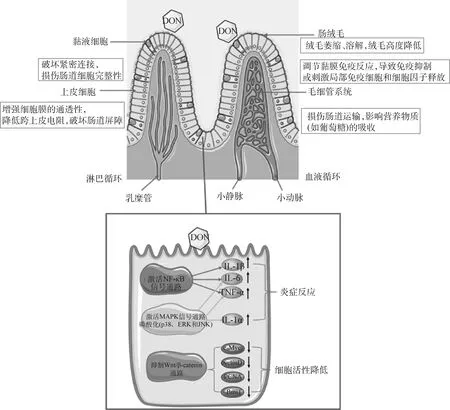
图3 DON对猪肠道的毒害作用及其可能机制Fig.3 Toxic effects of DON on pig intestines and its possible mechanism
2.3 调控DON毒理作用的分子遗传解析研究进展
随着分子生物学技术的发展和组学分析技术的应用,从分子水平(DNA和RNA等)探析DON毒理作用的研究也逐渐受到关注。Zhang等[41]使用猪肠上皮细胞系研究了DON对IPEC-J2的细胞毒性,通过RNA-seq筛选并鉴定出320个上调基因和160个下调基因,这些差异基因主要富集在免疫相关途径上;进一步分析发现,DON通过激活p38 MAPK和ERK1诱导IPEC-J2细胞中促炎因子的产生。Xu等[65]研究发现,DON处理显著降低了IPEC-J2的细胞活力,阻断了G2/M期细胞周期,增加了细胞凋亡率,显著下调了SLC4A11和MFSD3基因的表达水平;而且SLC4A11和MFSD3的过表达可以增强细胞活力,提示猪SLC4A11和MFSD3基因过表达可能具有缓减DON诱导的细胞毒性的作用。赵呈祥[66]利用全基因组DNA甲基化分析技术,鉴定了DON处理的IPEC-J2细胞全基因组DNA甲基化和基因表达变化模式,发现DON可诱导IPEC-J2细胞全基因组DNA甲基化水平升高;另外,确定27个基因启动子甲基化水平与其表达水平呈负相关,初步揭示VAV3、SLC4A11、ST3GAL5和DUSP9等基因在DON引起的细胞毒性反应中的表达变化可能受其启动子DNA甲基化的调控。
此外,Segura-Wang等[67]研究了DON诱导的猪肝、空肠和血清中microRNA表达的变化。在组织中仅检测到microRNA表达的轻微变化,其中ssc-miR-10b在暴露于DON的仔猪肝中被下调;在血清中,一些microRNA在DON暴露后差异表达,其中4种通过qPCR进行了验证(ssc-miR-16、ssc-miR-128、ssc-miR-451、ssc-miR-205);预测的microRNA靶基因显示出多种途径的富集,包括PIK3-AKT、Wnt/β-连环蛋白和黏附连接。Yang等[38]研究表明,DON可以诱导pGCs的形态变化和凋亡,同时抑制细胞增殖;RNA-seq进一步分析揭示了5 937个差异表达基因,其中1 995个在DON处理组中下调,3 942个上调;KEGG富集分析表明,差异基因主要富集在IL-17信号通路、真核核糖体合成通路、RNA转运通路和RNA降解等重要代谢通路,并推测DON对pGCs的影响与DNA损伤过程有关。
Wang等[68]揭示了暴露于DON后猪肠上皮细胞的全基因组转录谱,在DON暴露和对照细胞样品之间共鉴定出110个差异表达的miRNAs和143个差异表达的lncRNAs,其中miR-330直接靶向MAPK15基因进而调控DON毒性,是一种潜在生物标志物,可用于监测DON毒性。Xie等[69]发现,猪乳外泌体上调了IPEC-J2细胞中miR-181a、miR-30c、miR-365-5p和miR-769-3p的表达,然后下调其靶向基因在p53通路中的表达,继而通过促进细胞增殖和抑制细胞凋亡来减轻DON诱导的损伤,最终保护肠道免受损伤。因此,在饲料或食物中添加富含miRNA的外泌体可作为一种新型预防措施应对DON导致的坏死性小肠结肠炎。表1 列出了目前可能参与调控DON毒理作用的潜在生物标志物。

表1 参与调控DON毒理作用的潜在生物标志物Table 1 Potential biomarkers involved in regulating the toxicological effects of DON
3 DON解毒剂研究进展
3.1 饲料添加剂的解毒作用
解毒生物学方法表明,精氨酸或谷氨酸等饲料添加剂可有效降低霉菌毒素对幼猪的毒性作用,主要表现在缓解猪DON应激和免疫相关细胞因子引起的损伤、改善了抗氧化系统的不平衡和霉菌毒素引起的肠道结构异常,改善DON导致的空肠形态、血清、空肠和回肠中的氨基酸浓度和转运蛋白的表达水平[70-72]。N-乙酰半胱氨酸能缓解由DON引起的仔猪支持细胞结构的损伤、周期停滞和氧化应激损伤[39]。另外,有研究表明乳果糖和富氢水组成的添加剂通过升高抗氧化水平能有效缓解DON对仔猪卵巢造成的危害[37](表2)。
3.2 吸附剂的解毒作用
毒素污染饲料添加2 mg·kg-1蒙脱石或1.0 mg·kg-1蒙脱石+0.5 mg·kg-1酵母培养物复合物可降低霉菌毒素对免疫系统和肝的影响,改善仔猪生长性能[73]。霉菌毒素吸附剂(纳米级硅酸盐血小板以及蒙脱石和酵母细胞壁的混合物)可以通过调节p38、ERK和JNK的磷酸化来减弱IPEC-J2细胞中DON诱导的炎症反应[58]。Zhang等[74]发现,改良水合铝硅酸钙钠(HSCAS)吸附剂能减轻DON诱导的断奶仔猪血清抗体滴度、胸腺组织病理学、细胞凋亡、氧化还原状态、炎症和细胞凋亡信号的变化。Horky等[75]发现,含有纯化和活化膨润土的霉菌毒素吸附剂改善了DON诱导的育肥猪中肝的组织病理变化,降低了氧化应激,并且保护了空肠组织不受破坏。Santos等[76]研究表明,由吸附性粘土矿物组成的霉菌毒素灭活剂可以减轻DON对保育猪生长性能和氧化应激的负面影响(表2)。
3.3 微生物的解毒作用
鼠李糖乳杆菌RC007降低了DON导致的组织学改变、促炎症反应、细胞旁通透性和MAPK激酶的磷酸化,可以作为益生菌使用,降低DON对猪只的肠道毒性[77]。肠道微生物群已被证明有助于对抗霉菌毒素的影响,Reddy等[78]通过对猪盲肠内容物进行16S rRNA测序发现,乳酸杆菌和类杆菌在缓减DON对猪盲肠的毒害作用和改善免疫应答过程中起关键作用。细菌(真杆菌BBSH797)和酶(伏马菌素酯酶 FumD)能够有效改善DON引起的仔猪血液学、生物化学、组织形态学变化和免疫反应[79]。Garcia等[80]研究发现,酿酒酵母RC016可改善动物生长和肠道免疫系统,并且可以通过抵消DON诱导的仔猪断奶相关肠道炎症来改善肠道健康。同样,酿酒酵母S.cerevisiaeboulardii可以减轻DON诱导的氧化应激,减少DON诱导的肠道转录组改变[81](表2)。
3.4 天然成分(提取物)的解毒作用
小檗碱(berberine, BBR)亦称黄连素,是黄连抗菌的主要有效成分,一方面可以通过增加血清抗氧化酶和T细胞表面抗原的表达,减少小肠促炎细胞因子的释放来抑制DON诱导的肠道损伤;另一方面可以通过调控NF-κB和MAPK信号通路来调节DON诱导的肠道损伤、免疫抑制和氧化应激,最终维持仔猪肠道健康[82]。白藜芦醇(resveratrol, RES)是一种植物中的酚类植物抗毒素,RES补充剂通过增强肠道屏障功能、减轻肠道炎症和氧化损伤以及正向调节肠道微生物群来改善DON诱导的仔猪肠道健康[83]。落新妇苷(astilbin, AST)是一种天然抗氧化剂,20 μg·mL-1的AST可通过增强抗氧化酶活性和肠道屏障功能来改善DON诱导的IPEC-J2细胞氧化应激、炎症和细胞凋亡,降低炎症和凋亡基因的表达,改善屏障功能和营养转运吸收[84]。绿原酸(chlorogenic acid, CGA)是金银花的主要抗菌、抗病毒有效药理成分之一,CGA预处理可减轻DON诱导的IPEC-J2细胞毒性、炎症和凋亡,并保护肠细胞完整性免受DON损伤[85]。番茄红素(lycopene, LYC)是一种有效的抗氧化类胡萝卜素,LYC能够在DON暴露下调节Keap1/Nrf2信号通路来保护肠上皮免受氧化损伤[86]。表2总结了目前可能的DON解毒剂及主要功效。
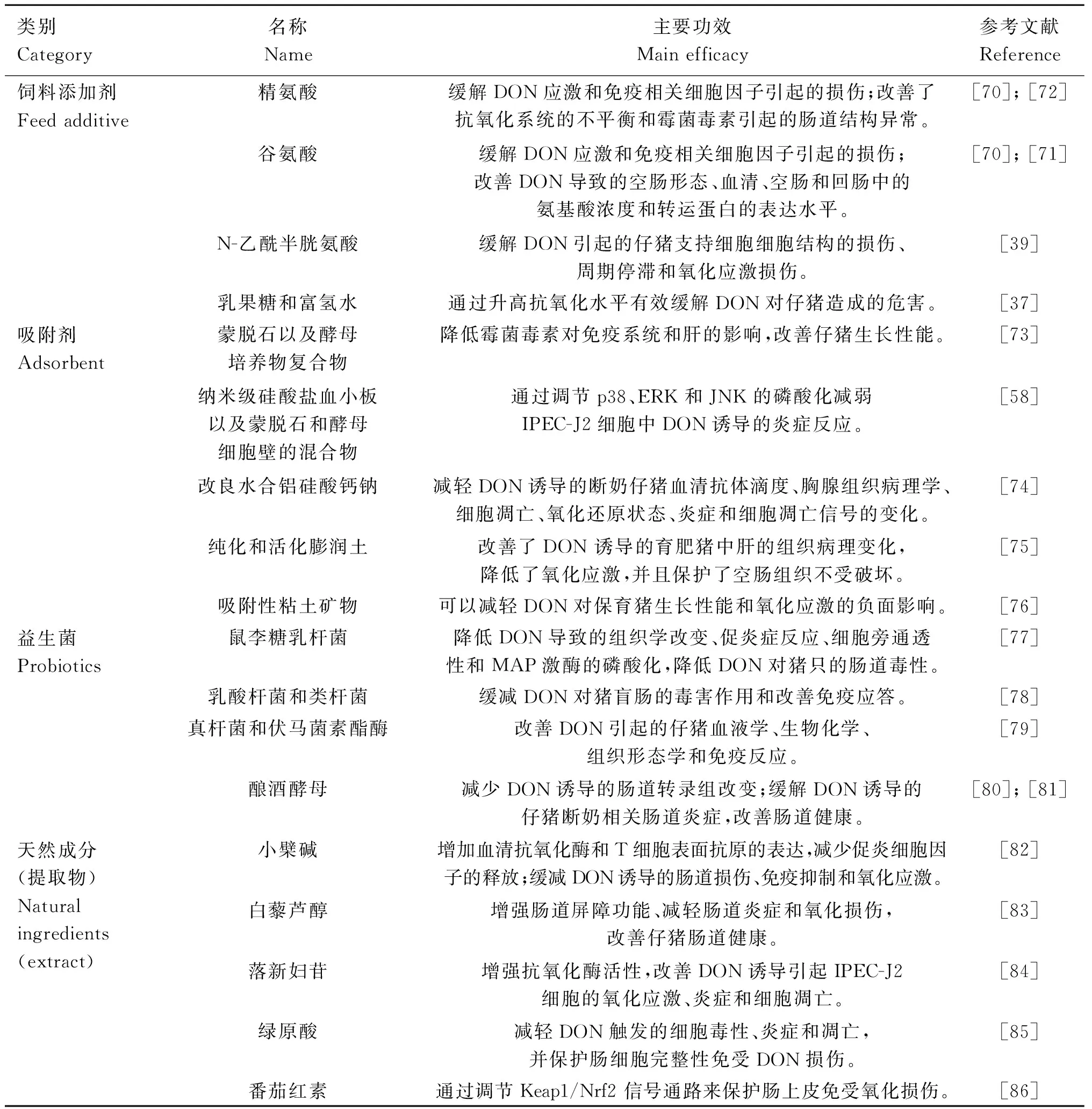
表2 DON解毒剂及主要功效Table 2 DON antidotes and their main functions
4 总结与展望
DON作为一种猪最易感的霉菌毒素,能够导致猪全身组织和器官的损伤和病变,造成猪只生产性能的下降,给养猪业带来巨大的挑战。目前饲料加工工艺的改进、饲料防霉措施的改善、以吸附剂和生物制剂为主的DON解毒剂的研发能够在一定程度上缓减DON的毒害作用,但并不能从根本上提高猪对DON的抵抗力。随着分子生物学技术的迅速发展,调控DON毒理作用的分子遗传机制得到进一步揭示。因此,利用现代分子生物学技术(高通量测序技术、多组学分析技术、基因编辑技术等)对猪耐受(抵抗)DON的毒害作用进行基因功能和调控机制的深入研究,挖掘有意义的生物标志物或分子标记并应用于猪的抗病育种,从遗传本质提高猪对DON的抵抗力,将对保障养猪业持续健康发展具有深远的影响。
