两种池塘养殖模式下异育银鲫(Carassius auratus gibelio)的形质特征差异分析*
2023-01-17徐英杰戎华南刘哲宇陈汉春王志铮
徐英杰 戎华南, 陈 雨 刘哲宇 陈汉春 王志铮
两种池塘养殖模式下异育银鲫()的形质特征差异分析*
徐英杰1戎华南1, 2陈 雨1刘哲宇1陈汉春2王志铮1①
(1. 浙江海洋大学水产学院 浙江舟山 316022; 2. 慈溪市水产技术推广中心 浙江慈溪 315300)
探究引起银鲫在不同养殖模式下的形质差异原因及形质饰变途径和机制, 进而揭示养殖模式致其改变生存对策的内在逻辑, 对于指导银鲫养殖模式的改进与优化具重要现实意义。随机选取池塘生态主养模式(M1)和池塘生态套养模式(M2)下经7个月养殖的异育银鲫夏花苗种各70尾为研究对象, 采用主成分分析和判别分析方法, 系统开展了两种池养模式下异育银鲫形质差异研究, 结果表明: (1) 池养期间两实验群体的生长速度总体上呈M2>M1, 在21项生物学表型性状中, M2实验群体显著大于M1实验群体的为1(体长)、4(头宽)、11(肛后体长)和NM (净体质量) (<0.05); (2) 所涉20项形质评价性状中, M1、M2实验群体间具显著差异(<0.05)的高达16项, 聚类分析也指示两者间的欧式距离已达到显著水平(<0.05), 即M1、M2实验群体已在形质特征上出现显著分化; (3) 经主成分分析, 提取到的 5个特征值均大于 1 的主成分, 其累积贡献率达 80.844%, 其中PC1可归纳为表征机体消化代谢水平的公共因子, PC2可归纳为表征脏器可容纳空间几何比例的公共因子, PC3、PC4和PC5可统归为表征食物获取方式的公共因子; (4) 采用逐步判别法, 以判别贡献率较大的眼间距/侧线长、净重系数、肝系数、胃系数、肠系数、鳃系数和Fulton指数为自变量, 所建Fisher分类函数方程组可较清晰区分M1和M2实验个体, 其中M1实验群体的判别准确率1、2分别为98.6%和94.5%, M2实验群体则分别为94.3%和98.5%, 两实验群体的综合判别准确率为96.4%。所得结果可为异育银鲫池塘生态主养模式和生态套养模式的改良与优化提供科学依据。
池塘套养; 池塘主养; 异育银鲫(); 形质特征; 多元分析
鲫()隶属于硬骨鱼纲(Osteichthyes)、鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鲫属(), 与青、草、鲢、鳙、鲤、鳊并称我国七大大宗淡水养殖鱼类(戈贤平, 2010)。异育银鲫系中国科学院水生生物研究所于20世纪70年代以方正银鲫() (♀) × 兴国红鲤() (♂) 经雌核发育培育的三倍体鲫鱼(楼允东, 1999)。自20世纪80年代在全国推广以来, 现已成为国内主要的养殖鲫种(Xue, 2001)。其中, 池塘主养和池塘套养是其目前最为常见的两种传统养殖模式。水产动物所处的生存环境与其养殖模式息息相关。Wagner等(2001)指出, 同种生物的不同群体为长期适应不同的生活环境而形成了各自特有的包括生长、发育、繁殖以及对环境因子的适应力等的生物学特性。Killen等(2011)也认为, 受内外环境、食物资源和遗传特性等的影响, 同种生物个体对环境变化存在不同的适应方式。形质作为反映生物外观表型和体质状况的生物学表型信息(王志铮等, 2012), 无疑其可塑性变化既是对所处生存环境的真实反映, 也是生存适应对策在宏观生物学水平的客观体现。因此, 探究引起目标水产养殖动物在不同养殖模式下的形质差异原因及形质饰变途径和机制, 进而揭示养殖模式致目标水产养殖动物改变生存对策的内在逻辑, 无疑对于指导养殖模式的改进与优化具重要现实意义。但迄今为止国内外尚未见有关不同池塘养殖模式下异育银鲫形质特征差异的专题报道。鉴于此, 本研究团队通过开展池塘生态主养和池塘生态套养两种养殖模式下异育银鲫生物学表型性状对体质量的影响效果差异研究, 发现两者已在-生存对策选择轴上发生明显偏移的基础上(陈雨等, 2022), 拟从形质特征角度, 借助主成分分析和判别分析, 进一步从宏观生物学水平降维解读两者间生存对策的差异, 以期为异育银鲫养殖模式改良与优化提供科学依据。
1 材料与方法
1.1 实验鱼
系2020年12月15日捕自慈溪市浒山华乔淡水鱼养殖场按常规方法养殖的异育银鲫, 养殖模式分别为池塘生态主养模式(记为M1)和池塘生态套养模式(记为M2), 具体放养参数见表1。M1和M2实验群体停食驯养24 h后, 各随机选取鳞片齐整、鱼体完好、反应灵敏、无病无伤的健壮个体70尾备测。
1.2 生物学测定
按陈雨等(2022)所用测量仪器和测量步骤, 逐尾开展M1和M2实验群体的生物学测定。其中, 体尺性状指标为体长(1, 吻端至尾椎骨末端的水平距离)、体高(2, 鱼体背腹部间的最大垂直距离)、头长(3, 吻端至鳃盖骨后缘的最大水平距离)、头宽(4, 头部左右两侧的最大水平距离)、眼后头长(5, 眼球中心到鳃盖后缘的水平距离)、眼间距(6, 左右眼球间的水平距离)、鼻间距(7, 左右鼻孔间的水平距离)、鳃盖高(8, 鳃盖骨在鱼体背腹部两交叉点间的垂直距离)、侧线长(9, 侧线起始点间的水平距离)、肛长(10, 吻端至肛门孔的水平距离)、肛后体长(11, 肛门孔至尾椎骨末端的水平距离)、肛部体高(12, 经过肛门孔的体高), 具体测量部位见图1; 称量性状指标为体质量(MB)、净体质量(NM, 去除内脏和鳃质量之后的体质量)、内脏质量(MV)、肝质量(ML)、胃质量(MS)、肠质量(MI)、鳃质量(MG)、心质量(MH)和鳔质量(MSB)。

表1 异育银鲫两种池塘养殖模式的放养参数
注: 池塘生态主养模式(M1)沿池塘四周密植铜钱草()
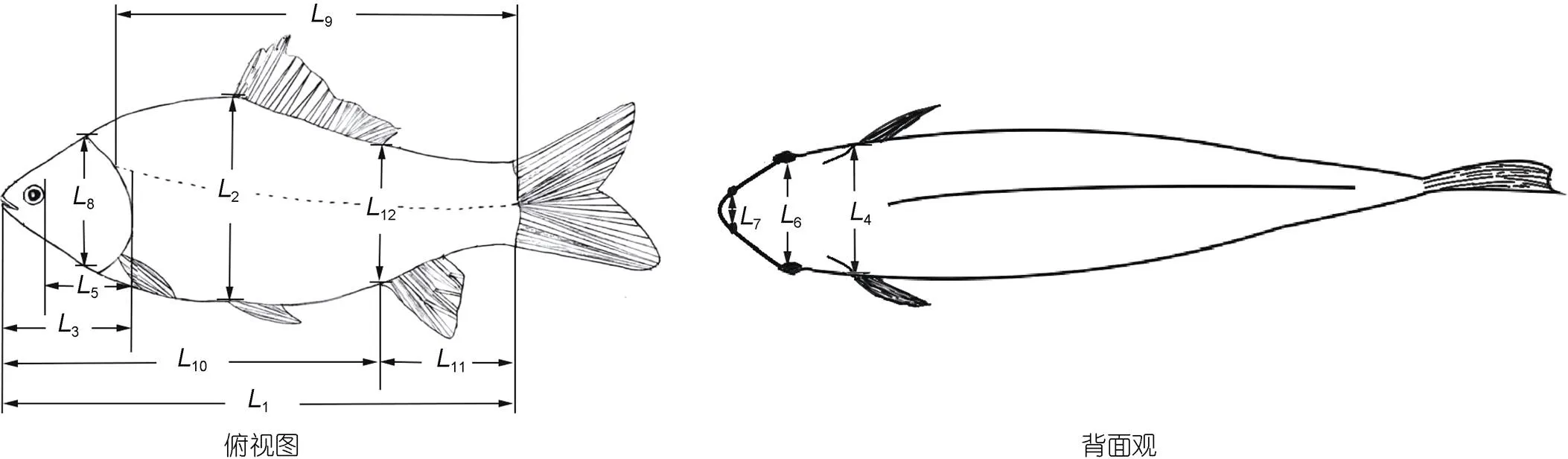
图1 形态测量示意图
注:1表示体长;2表示体高;3表示头长;4表示头宽;5表示眼后头长;6表示眼间距;7表示鼻间距;8表示鳃盖高;9表示侧线长;10表示肛长;11表示肛后体长;12表示肛部体高
1.3 数据处理
采用EXCEL(2019)软件整理上述测定结果, 分别计算M1实验群体和M2实验群体的各项体尺比例性状(以侧线长为分母)、质量比例性状(以体质量为分母)、肥满度性状(Fulton指数=100×NM/13)等形质评价指标的均值与标准差, 并检验组间差异显著性(视<0.05为显著水平); 以通过Bartlett球形检验(<0.01)和KMO适合度检验(KMO值>0.700)为因子分析前置条件, 以特征值大于1且累计贡献率大于80%为主成分(PC)提取原则遴选形质评价指标, 并运用SPSS23.0软件开展主成分分析; 按王志铮等(2012)的方法, 采用逐步导入剔除法进行判别分析, 并通过计算判别准确率和综合判别率验证所建Fisher分类函数方程组的可靠性。
2 结果
2.1 测量表型性状
由表1可见, M1实验群体和M2实验群体在本研究所涉21项表型性状间的差异主要表现为: (1) 从体尺性状看, 除5(眼后头长)、6(眼间距)、8(鳃盖高)和10(肛长)无显著差异(>0.05)外, M1实验群体显著大于M2实验群体的为2(体高)、3(头长)、7(鼻间距)、9(侧线长)和12(肛部体高) (<0.05), 小于M2实验群体的为1(体长)、4(头宽)和11(肛后体长) (<0.05)。鉴于M2实验群体放养时间略迟于M1实验群体(表1), 故其5(眼后头长)、6(眼间距)、8(鳃盖高)、10(肛长)、1(体长)、4(头宽)和11(肛后体长)等7个性状的增速均明显大于M1实验群体, 尤以1(体长)、4(头宽)和11(肛后体长)为甚; (2) 从质量性状看, 除MB (体质量)无显著差异(>0.05)外, M1实验群体显著小于M2实验群体的仅为NM (净体质量) (<0.05), 其余性状均呈M1实验群体显著大于M2实验群体(<0.05), 即M2实验群体MB (体质量)和NM (净体质量)的增速明显大于M1实验群体, 尤以NM (净体质量)为甚。综上可知, 本研究所涉两实验群体在池养期间的生长速度总体上呈M2>M1(<0.05)。
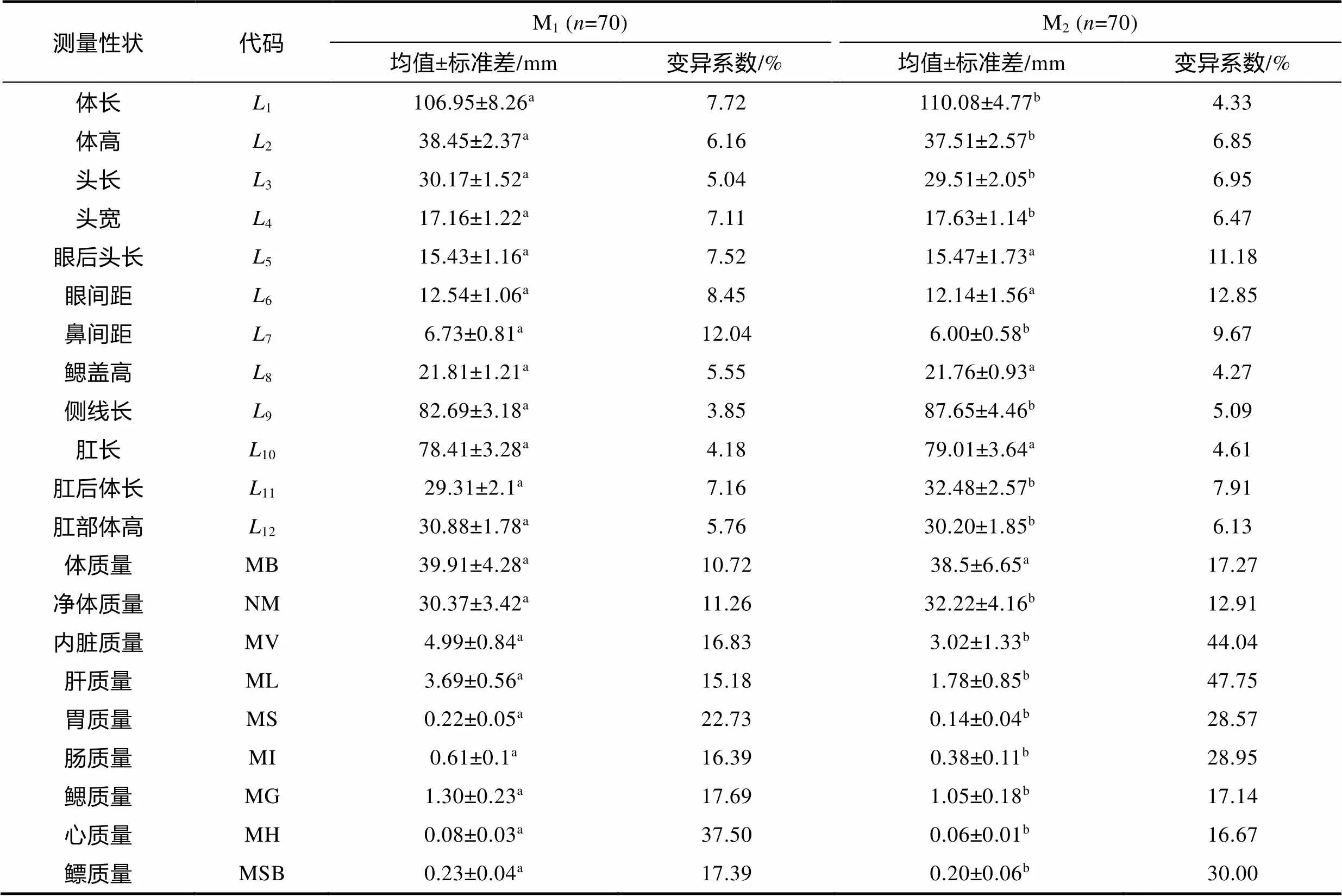
表2 实验群体间测量表型的差异
注: 上标不同字母表示组间存在显著差异(<0.05)
2.2 形质评价指标
对表2所涉21项测量表型性状做归一化处理后, 得表3。由表3可见, M1实验群体和M2实验群体在本研究所涉20项形质评价指标的差异主要表现为: (1) 从体尺比例性状看, 除10(肛后体长/侧线长)呈M1实验群体显著小于M2实验群体(<0.05)外, 其余10项性状均呈M1实验群体显著大于M2实验群体(<0.05), 即以侧线长为标尺对甄别M1、M2两实验群体间体型差异具良好的区分度; (2) 从质量比例性状看, 除1(净重系数)、6(鳃系数)、7(心系数)、8(鳔系数)无显著差异(>0.05)外, 余下的2(内脏系数)、3(肝系数)、4(胃系数)、5(肠系数)均呈M1实验群体显著大于M2实验群体(<0.05), 即消化机能在区分M1、M2两实验群体间脏器生理功能差异上更具重要性; (3) Fulton指数呈M1实验群体显著大于M2实验群体(<0.05), 即M1实验群体的生理营养状况显著优于M2实验群体。综上可知, M1、M2两实验群体的形质特征差异主要集中于体型、消化机能和营养生理状况上。
2.3 主成分分析
经Bartlett球形检验(<0.01)和KMO适合度检验(KMO值为0.85), 有必要对表3所涉形质评价指标做主成分分析。由表4可见, 经降维处理提取到的5个主成分的特征值均大于1且方差累计贡献率为80.844%, 表明它们是能概括M1和M2两实验群体间形质差异的主要公共因子。其中, PC1的方差贡献率为37.263%, 主要影响变量(载荷绝对值>0.6)为8个质量比例性状, 其影响力排序呈6(鳃系数)>7(心系数)>1(净重系数)>8(鳔系数)>5(肠系数)>2(内脏系数)>4(胃系数)>3(肝系数); PC2的方差贡献率为22.801%, 主要影响变量为7个体尺比例性状, 其影响力排序呈2(体高/侧线长)>11(肛部体高/侧线长)>3(头长/侧线长)>9(肛长/侧线长)>8(鳃盖高/侧线长)>7(鼻间距/侧线长)>4(头宽/侧线长); PC3的方差贡献率为8.426%, 其主要影响变量为(Fulton指数)和1(体长/侧线长), 其中1(体长/侧线长)为本研究所涉5个主成分中唯一的负向主要影响变量; PC4的方差贡献率为6.312%, 主要影响变量的影响力排序为5(眼后头长/侧线长)>6(鼻间距/躯干长); PC5的方差贡献率为6.006%, 主要影响变量仅为10(肛后体长/侧线长)。综上可知, 上述5个主要公共因子所涉的20个主要影响变量可综合反映两实验群体间在体质、体型、运动、摄食和消化等方面的差异。鉴于, PC1的方差贡献率最大且主要影响变量也最多的实际, 分别绘制PC1与其他4个主成分间的得分散布图得图1。由图2可见, 本研究所涉两实验群体沿FAC1轴自左向右大体上分别占据各自独立的区域, 但仍存在一定程度的重叠或错位, 表明PC1在归纳本研究实测样本的类群归属上明显受到了某些关键影响变量的干扰。
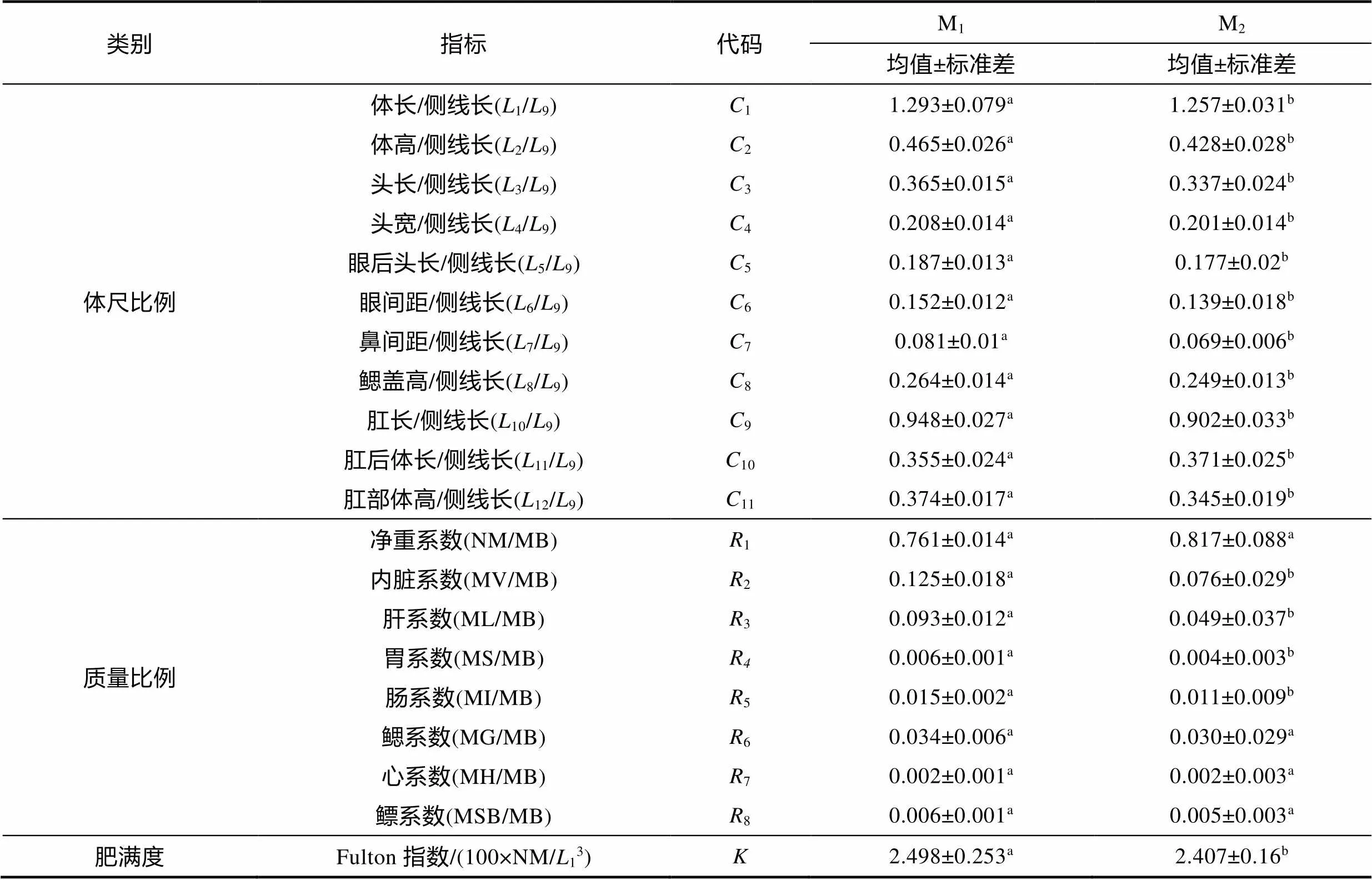
表3 两种养殖模式异育银鲫形质评价指标参数统计
注: 上标不同字母表示组间存在显著差异(<0.05)

表4 M1实验群体和M2实验群体形质量评价指标的主成分分析
注: 载荷值>0.6示各主成分的主要影响变量; 上标星号表示该变量为主要影响变量

注: ▲表示M1实验群体, ○表示M2实验群体, 下同
2.4 判别分析
采用逐步导入剔除法, 从表3所列的20个形质性状中筛留对判别贡献较大的6(眼间距/侧线长)、1(净重系数)、3(肝系数)、4(胃系数)、5(肠系数)、6(鳃系数)和(Fulton指数)进行判别分析, F检验表明这些性状均达到极显著水平(<0.01)。根据上述7个性状建立基于实测样本的Fisher分类函数方程组于表5。经验证, M1实验群体的判别准确率1、2分别为98.6%和94.5%, M1实验群体的则分别为94.3%和98.5%, 两实验群体的综合判别准确率为96.4% (表6)。另, 所绘典型判别函数判别得分散布图(图3), 亦直观印证了上述判别分析的可靠性。
3 讨论
3.1 取食策略与形质特征间的关联分析
“消耗-获利”的权衡既是生物受生境驱动选择生存方式与生存对策的核心, 也是生物为响应生境改变谋求利益最大化而引起形质饰变的重要生理生态基础。动物的运动代谢和摄食代谢既是两个相互关联的重要生理过程(Hicks, 2004; Fu, 2007),也是动物基于所处生境选定“消耗-获利”生存模式的具体表现。有学者根据运动和摄食这两个生理过程的相对重要性, 将其分为优先保证某一过程的“优先模式”和同时保证这两个过程的“添加模式”(Blaikie, 1996; Alsop, 1997)。鱼类基于“消耗-获利”权衡之考量, 势必会就生境适应性分别形成并固化倾向以伏击取食为主或以积极取食为主的两种食物获取策略。观察发现, 本研究所涉M1养殖模式银鲫处于低烈度种间斗争的主养环境, 运动抢食是其获取食物的主要途径, M2养殖模式银鲫较主养对象凡纳滨对虾养殖密度低且放养时间迟, 在运动抢食上明显处于弱势地位, 致使伏击取食和摄取凡纳滨对虾摄食后残留的饲料成为其获取食物的主要途径, 即异育银鲫在M1和M2养殖模式下分别采取了运动和摄食并重的“添加模式”和更注重摄食的“优先模式”。研究发现, 伏击取食动物与积极摄食的动物相比, 具饥饿期间消化脏器质量小和摄食后消化脏器质量大幅增加的特质(Secor, 1998; Secor, 2001; 付世建等, 2008)。无疑, 本研究中M2实验群体肝质量、胃质量、肠质量以及肝系数、胃系数和肠系数的变异系数均明显大于M1实验群体的结果(表2, 表3), 在进一步印证M1、M2实验群体分别采取积极取食为主和伏击取食为主的食物获取策略可靠性的同时,也揭示了种间竞食强度差异是导致异育银鲫取食策略发生明显改变的主因。故, M1和 M2实验群体为贯彻各自的食物获取策略势必在形质特征上出现一定程度的分化。

表5 Fisher分类函数方程组

表6 判别分类结果
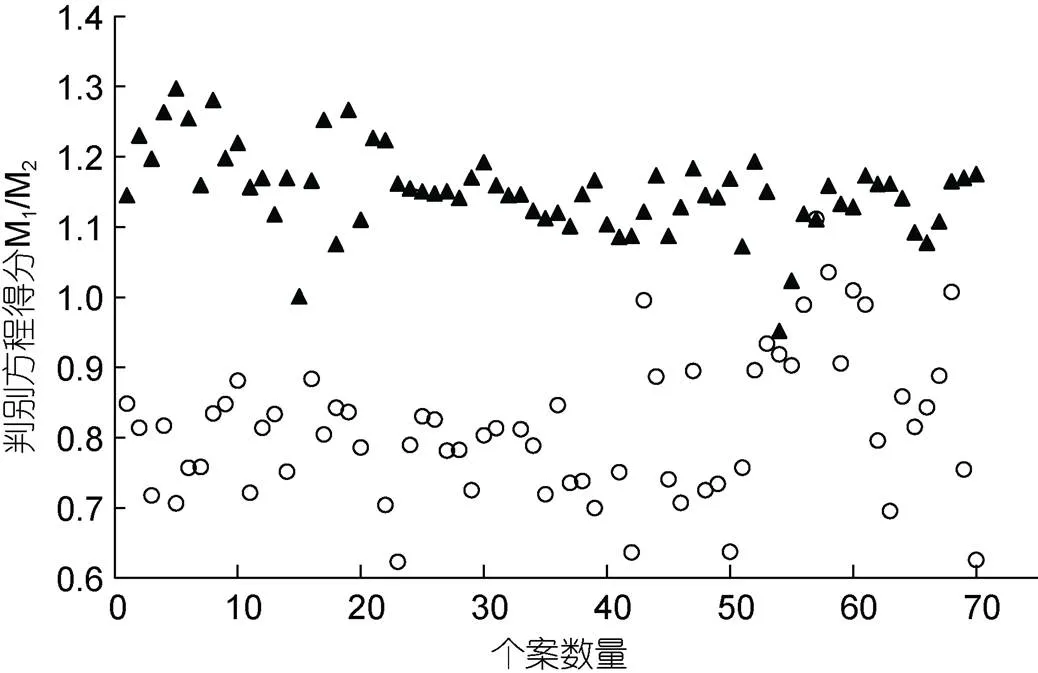
图3 典型判别函数判别得分散布图
肝、胃、肠系动物最重要的活性消化器官。研究表明, 活性器官的脏器系数与机体代谢水平呈正相关(Itazawa, 1983; Oikawa, 1992)。由表3、表4和表5可见, PC1主要影响变量中被列入的5个判别变量, 除1(净重系数)和6(鳃系数)无组间显著差异(>0.05)外, 余下的3(肝系数)、4(胃系数)、5(肠系数)均呈M1显著大于M2(<0.05), 即M1实验群体的消化代谢水平显著高于M2实验群体, 故可将PC1归纳为表征机体消化代谢水平的公共因子; PC2主要影响变量2(体高/侧线长)、3(头长/侧线长)、4(头宽/侧线长)、7(鼻间距/侧线长)、8(鳃盖高/侧线长)、9(肛长/侧线长)、11(肛部体高/侧线长)所示肛前鱼体几何空间比例, 因与PC1所涉5个判别变量在体尺维度上高度叠意, 而均未被列入判别变量, 故可将PC2归纳为表征脏器可容纳空间几何比例的公共因子; 一般而言, 鱼类的摄食过程按序依次为寻找、发现、摄取和消化(Toften, 2003; Hara, 2006)。其中, 测线感受系统具识别和定位猎物的功能(梁旭方, 1998), 鱼眼在鱼类运动过程中具补偿运动以保持视野稳定的作用(Collin, 2003)。肥满度系反映动物生理状态或营养状况的重要指标(王寿兵等, 1999; 王志铮等, 2012)。无疑, PC3、PC4、PC5主要影响变量中被列入的2个判别变量(Fulton指数)和6(眼间距/侧线长)均呈M1显著大于M2(<0.05)的结果, 在强调异育银鲫主要依靠视觉和侧线来感知和定位食场, 以及M1实验群体较M2实验群体具更佳的营养生理状况和更为宽广且稳定的寻食视野的同时, 也进一步印证了M1、M2实验群体分别采取积极取食为主和伏击取食为主的食物获取策略的客观事实, 故可将PC3、PC4、PC5统归为表征食物获取方式的公共因子。M1、M2实验群体具不同的体质量增长机制, 研究表明影响M1实验群体体质量的核心体尺性状为侧线长, 从属性状为全长和体高, 核心称量性状为净体质量, 从属性状为肝质量、肠质量和胃质量, 而影响M2实验群体体质量的核心体尺性状为体高, 从属性状为体宽、全长和侧线长, 核心称量性状为净体质量, 从属性状仅为肝质量(陈雨等, 2022)。综上可知, 食物获取方式与机体消化代谢水平是两个互为关联的方面, 即食物获取方式可通过影响机体消化代谢水平, 进而显著改变体质量的增长机制。
3.2 形质特征与生存对策间的关联分析
已有研究表明, 适量种植水生植物对改善并稳定养殖水体环境(Pokorny, 1990; 周遗品等, 2011), 进而提高池养水产动物存活率、生长效率和养成品质(刘鑫等, 2003; 徐增洪等, 2016)具重要作用。异育银鲫属底层杂食性鱼类, 本研究所涉M1养殖模式通过种植水草和大量养殖滤食性鱼类(M1的鳙鱼放养密度为M2的10倍), 在改善养殖环境并提高池塘养殖承载力上较M2养殖模式具明显的优越性, 但高烈度的种内竞争(M1异育银鲫养殖密度为M2的25倍)依然是M1实验群体必须要面对的最大生存胁迫因子。与此相反, 套养于凡纳滨对虾高密度养殖生境下的M2实验群体则必须面对激烈的种间竞争和高严峻度的理化环境。由表3和表5所示, 本研究所涉7个判别变量中仅1(净重系数)和6(鳃系数)无组间显著差异(>0.05), 而其余则均呈M1显著大于M2(<0.05)的结果看, 维持1(净重系数)和6(鳃系数)稳定既是本研究所涉M1、M2实验群体针对不同生境压力选择相应生存对策的重要生理基础, 也是M1、M2实验群体间生长速度(表2)和消化脏器系数(表3)差异被显著放大的根本原因。即(1) M1实验群体为响应低严峻度理化环境, 通过显著上调消化代谢水平以提高消化脏器系数并促进干物质积累, 有效贯彻并实施了与高烈度种内竞争相适配的以运动抢食为特征的积极取食策略, 这种高能耗取食代谢特征无疑会限制其生物学表型的快速增长; (2) M2实验群体为弥补高严峻度理化环境下的供氧不足, 通过显著下调消化代谢水平以降低消化脏器系数并延缓干物质积累, 有效贯彻并实施了与高烈度种间竞争相适配的伏击取食策略, 这种低能耗取食代谢特征势必会促进其生物学表型的快速增长。综上可知, 实现供氧压力与消化代谢强度间的有机统一, 既是M1和M2实验群体基于各自所处食场生境选择生存方式的两个互为关联的方面, 也是M1和M2实验群体有效贯彻并实施不同取食策略的重要生态生理基础。无疑, 陈雨等(2022)所述M2实验群体在-生存对策选择轴上明显表露出朝选择偏移的原因, 实际上是与M2实验群体因消化代谢水平持续受限, 致使后续机体能量供应难以支持其进一步朝个体大型化方向发展有关。
4 结论
(1) 种间竞食强度差异是导致M1、M2实验群体间取食策略发生明显分化的主因。基于各自所处食场生境, M1实验群体通过显著上调消化代谢水平和促进机体干物质积累, 以贯彻积极取食的食物获取策略, 固化了运动和摄食并重的高能耗“添加模式”, 从而限制了其生物学表型的快速增长; M2实验群体通过显著下调消化代谢强度和延缓机体干物质积累, 以贯彻伏击取食的食物获取策略, 固化了更注重摄食的低能耗 “优先模式”, 从而促进了生物学表型的快速增长, 这既是导致本研究养殖期间M1实验群体生长速度总体上显著低于M2实验群体的根本原因, 也是引起M1和M2实验群体形质特征出现明显分化的生态生理基础。
(2) 异育银鲫主要依靠视觉和侧线来感知并定位食场, 其机体消化代谢水平取决于所选定的食物获取方式。经主成分分析, 提取到的 5个特征值均大于 1 的主成分中, PC1可归纳为表征机体消化代谢水平的公共因子, PC2可归纳为表征脏器可容纳空间几何比例的公共因子, PC3、PC4和PC5可统归为表征食物获取方式的公共因子。判别分析显示, 以判别贡献率较大的眼间距/侧线长、净重系数、肝系数、胃系数、肠系数、鳃系数和Fulton指数为自变量, 建立的Fisher分类函数方程组可较清晰区分M1和M2实验个体, 其中M1实验群体的判别准确率1、2分别为98.6%和94.5%, M2实验群体的则分别为94.3%和98.5%, 两实验群体的综合判别准确率为96.4%。
(3) 实现供氧压力与消化代谢强度间的有机统一, 既是异育银鲫基于所处生境选择生存方式的两个互为关联的方面, 也是异育银鲫有效贯彻不同取食策略的重要生态生理基础。与M1实验群体相较, 在-生存对策选择轴上M2实验群体明显表露出朝选择偏移的原因, 与其消化脏器系数明显偏低, 导致后续基于消化代谢的能量供应难以支持进一步朝个体大型化方向发展有关。
王寿兵, 蒋朝光, 屈云芳, 等, 1999. 野生和人工养殖辽宁中国林蛙肥满度和重/长指标的初步研究[J]. 应用生态学报, 10(1): 91-94.
王志铮, 杨磊, 朱卫东, 2012. 三种养殖模式下日本鳗鲡养成品的形质差异[J]. 应用生态学报, 23(5): 1385-1392.
戈贤平, 2010. 我国大宗淡水鱼养殖现状及产业技术体系建设[J]. 中国水产(5): 5-9.
付世建, 曹振东, 谢小军, 2008. 鱼类摄食代谢和运动代谢研究进展[J]. 动物学杂志, 43(2): 150-159.
刘鑫, 王哲, 张一, 等, 2003. 两种沉水植物对黄颡鱼()夏花培育水体主要水质因子的影响[J]. 湖泊科学, 15(2): 184-188.
陈雨, 戎华南, 刘哲宇, 等, 2022. 两种池养模式下异育银鲫()生物学表型对体质量影响效果的差异分析. 海洋与湖沼, 53(5): 1161-1169.
周遗品, 刘雯, 雷泽湘, 等, 2011. 铜钱草对城市生活污水的净化效果[J]. 仲恺农业工程学院学报, 24(2): 9-12.
徐增洪, 刘国锋, 水燕, 等, 2016. 人工栽培水草对池塘养殖生态环境和虾蟹生长的影响[J]. 江苏农业科学, 44(6): 328-331.
梁旭方, 1998. 鱼类侧线机械感觉猎物识别和定位的研究进展[J]. 动物学杂志, 33(2): 49-52.
楼允东, 1999. 鱼类育种学[M]. 北京: 中国农业出版社.
ALSOP D H, WOOD C M, 1997. The interactive effects of feeding and exercise on oxygen consumption, swimming performance and protein usage in juvenile rainbow trout () [J]. The Journal of Experimental Biology, 200(17): 2337-2346.
BLAIKIE H B, KERR S R, 1996. Effect of activity level on apparent heat increment in Atlantic cod,[J]. Canadian Journal of Fisheries and Aquatic Sciences, 53(9): 2093-2099.
COLLIN S P, SHAND J, 2003. Retinal sampling and the visual field in fishes [M] // COLLIN S P, MARSHALL N J. Sensory Processing in Aquatic Environments. New York: Springer: 139-169.
FU S J, CAO Z D, PENG J L, 2007. Effect of feeding and fasting on excess post-exercise oxygen consumption in juvenile southern catfish (Chen) [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 146(3): 435-439.
HARA T J, 2006. Feeding behaviour in some teleosts is triggered by single amino acids primarily through olfaction [J]. Journal of Fish Biology, 68(3): 810-825.
HICKS J W, BENNETT A F, 2004. Eat and run: prioritization of oxygen delivery during elevated metabolic states [J]. Respiratory Physiology & Neurobiology, 144(2/3): 215-224.
ITAZAWA Y, OIKAWA S, 1983. Metabolic rates in excised tissues of carp [J]. Experientia, 39(2): 160-161.
KILLEN S S, MARRAS S, MCKENZIE D J, 2011. Fuel, fasting, fear: routine metabolic rate and food deprivation exert synergistic effects on risk-taking in individual juvenile European sea bass [J]. Journal of Animal Ecology, 80(5): 1024-1033.
OIKAWA S, TAKEMORI M, ITAZAWA Y, 1992. Relative growth of organs and parts of a marine teleost, the porgy,, with special reference to metabolism-size relationships [J]. Japanese Journal of Ichthyology, 39(3): 243-249.
POKORNY J, KVET J, ONDOK J P, 1990. Functioning of the plant component in densely stocked fish ponds [J]. Bulletin of Ecology, 21(3): 44-48.
SECOR S M, 2001. Regulation of digestive performance: a proposed adaptive response [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 128(3): 563-575.
SECOR S M, DIAMOND J, 1998. A vertebrate model of extreme physiological regulation [J]. Nature, 395(6703): 659-662.
TOFTEN H, ARNESEN A M, JOBLING M, 2003. Feed intake, growth and ionoregulation in Atlantic salmon (L.) smolts in relation to dietary addition of a feeding stimulant and time of seawater transfer [J]. Aquaculture, 217(1/2/3/4): 647-662.
WAGNER E J, ARNDT R E, BROUGH M, 2001. Comparative tolerance of four stocks of cuttthroat trout to extremes in temperature, salinity, and hypoxia [J]. Western North American Naturalist, 61(4): 434-444.
XUE M, CUI Y B, 2001. Effect of several feeding stimulants on diet preference by juvenile Gibel carp (), fed diets with or without partial replacement of fish meal by meat and bone meal [J]. Aquaculture, 198(3/4): 281-292.
ANALYSIS OF THE DIFFERENCES IN MORPHOLOGY AND QUALITY OFUNDER TWO POND AQUACULTURE MODES
XU Ying-Jie1, RONG Hua-Nan1, 2, CHEN Yu1, LIU Zhe-Yu1, CHEN Han-Chun2, WANG Zhi-Zheng1
(1. School of Fisheries, Zhejiang Ocean University, Zhoushan 316022, China; 2. Cixi Fishery Technology Promotion Center, Cixi 315300, China)
Exploring the reasons for the differences in morphology and quality ofinduced by different aquaculture modes, finding the pathways and mechanisms of the difference-formation, and then revealing the internal logic of the mode inducing the changes in their survival strategy is of great practical significance for guiding the improvement and optimization ofaquaculture modeSeventy summer fingerlings ofwere randomly selected from the pond ecological monoculture mode (M1) and pond ecological polyculture mode (M2). Principal component analysis and discriminant analysis methods were used to systematically study the differences in morphology and quality ofunder two pond culture modes. Results showed that: (1) The growth rate of the two experimental groups during the pond culture was generally M2>M1. Among the 21 biological phenotypic traits, the1(body length),4(head width),11(post-anal body length), and NM (net body mass) of M2were significantly greater than M1(<0.05); (2) Among the 20 morphological evaluation traits involved, 16 items were significantly different (<0.05) between M1and M2groups, and cluster analysis also indicated that the Euclidean distance between the two groups had reached a significant level (<0.05), that is, the M1and M2were significantly differentiated in terms of morphological characteristics; (3) By the principal component analysis, the five extracted principal components whose eigenvalues were all greater than 1 had a cumulative contribution rate of 80.844%. Among them, PC1can be classified as a common factor that characterizes the level of digestion and metabolism, PC2can represent the geometric proportion of the space that the organ can accommodate, PC3, PC4, and PC5can be collectively classified as common factors that characterize the way of food acquisition; (4)Through the stepwise discriminant method, taking the interorbital space/lateral line length, net weight coefficient, liver coefficient, stomach coefficient, gut coefficient, gill coefficient, and Fulton index with a more significant contribution rate as independent variables, the Fisher classification function equations established can clearly distinguish the M1and M2.The discrimination accuracy rates1and2of the M1experimental group were 98.6% and 94.5%, respectively, of which the M2experimental group were 94.3% and 98.5%, respectively, and the complete discriminant accuracy of the two experimental groups was 96.4%%. The results obtained in this study can provide a scientific basis for improving and optimizing both the pond ecological monoculture mode and the pond ecological polyculture mode of the.
pond polyculture; pond monoculture;; characteristics of morphology and quality; multivariate analysis
Q953; S965
10.11693/hyhz20220500130
*国家星火计划项目, 2013GA701022号; 宁波市重大科技攻关项目, 2012C10032号; 慈溪市农业科技重点项目, CN201901号。徐英杰, 硕士研究生, E-mail: 1548117363@qq.com; 同等贡献第一作者: 戎华南, 高级工程师, 硕士生导师, E-mail: 13706742262@139.com
王志铮, 教授, E-mail: wzz_1225@163.com
2022-05-16,
2022-07-10
