南疆陆地棉杂种F2的遗传分析及遗传主效聚类
2023-01-16于霁雯裴文锋关永虎李长喜刘金伟王宝全梅拥军
郭 宏 于霁雯 裴文锋 关永虎 李 航 李长喜 刘金伟 王 伟 王宝全 梅拥军,*
南疆陆地棉杂种F2的遗传分析及遗传主效聚类
郭 宏1于霁雯2裴文锋2关永虎1李 航1李长喜1刘金伟1王 伟1王宝全2梅拥军1,*
1塔里木大学农学院, 新疆阿拉尔 843300;2中国农业科学院棉花研究所 / 棉花生物学国家重点实验室, 河南安阳 455000
采用加性-显性及其与环境互作的遗传模型对298个陆地棉品种(系)及其884个F2组合在新疆南疆阿拉尔垦区4个产量性状和5个纤维品质性状2年的资料进行遗传分析, 并对亲本的加性效应和显性效应进行了聚类, 以明确各类品种(系)的育种利用价值。结果表明, 在南疆陆地棉“矮(0.8~1.0 m)、密(225,000~300,000株 hm–2)、早、膜”高密度种植模式下, 298个亲本的加性效应被分为12类。第2类的23个品种(系)产量(除衣分外)和纤维品质性状的加性效应较高, 可通过这类品种(系)间杂交较易获得产量性状和纤维品质性状兼优的后代。第8类品种(系)的纤维品质性状的加性效应较高, 第4类品种(系)的产量性状加性效应较高, 通过2类品种间杂交可以获得产量性状和纤维品质性状互补的后代。298个亲本的显性效应被分成了9类。第8类包括8个品种(系), 其产量性状和纤维品质性状(除马克隆值外)的显性效应较高, 可作为综合利用产量性状和纤维品质性状杂种优势兼优的一类亲本。产量性状的显性效应平均值较高的是第8类, 第3类的纤维品质性状的显性效应较高。
陆地棉; F2; 产量性状; 纤维品质性状; 加性-显性模型; 聚类分析
棉花是新疆最主要的经济作物, 棉花产业对新疆经济发展具有十分重要的作用[1]。前人的研究表明, 陆地棉的产量和纤维品质性状是受多个基因控制和环境共同作用的数量性状且存在着显著的加性效应和显性效应, 具有复杂的遗传结构[2-7]。Roy等[8]的研究说明, 加性基因在性状表达中发挥了重要作用, 在后代的选择中具有重要价值。王保勤等[9]的研究说明, 永久F2群体的5个纤维品质性状的遗传方式基本一致, 都受加性、显性以及加性和环境互作等效应的影响。代勇强等[10]和秦鸿德等[11]研究表明, 分析亲本和F1或亲本和F2可以预测亲本的加性和显性效应。棉花的品种(系)很多, 选用这些品种作为杂交亲本就很困难。杂交育种主要利用的是亲本的加性效应, 杂优育种还要利用杂交组合的显性效应等。因此, 对亲本加性效应和显性效应进行聚类可以更有选择性地利用亲本的优良性状。新疆是我国最大的棉花生产基地, 前人关于亲本主效的研究很多, 但亲本和组合数目较少[12-15], 并且利用亲本和F2进行遗传效应研究的文章鲜有报道, 对多个亲本和F2不同性状遗传主效进行聚类的研究还未见报道。本研究旨在南疆陆地棉的栽培模式下对298个陆地棉亲本和F2遗传效应分析的基础上对亲本产量和纤维品质性状的加性效应和显性效应进行聚类分析, 对南疆陆地棉育种有针对性地利用这些亲本产量和纤维品质性状的遗传潜力具有重要意义。
1 材料与方法
1.1 材料与田间试验设计
试验分别于2018年和2019年在新疆阿拉尔市塔里木大学农学试验站进行。种植298个陆地棉品种(系) (附表1) (其中以298个品种(系)为母本、以其中的8个优质品系和新陆早30号为父本按不完全双列杂交遗传交配设计配制杂交组合)及其884个F2组合(附表2)。田间亲本及其F2组合随机区组设计, 重复2次, 每小区种植2行, 行长3.0 m, 地膜(0.60 m+0.10 m+0.55 m+0.10 m)覆盖种植, 株距0.10 m, 田间管理同大田。枯霜后1周内(10月20日左右), 亲本每小区统计10株棉花的铃数, F2每小区统计2行(55~60株)所有植株的铃数并算得单株铃数, 所有小区每小区收获中部1朵籽棉, 并记录收获朵数, 对籽棉称重并算得铃重, 轧花后测得皮棉, 用皮棉/籽棉×100%算得衣分, 由单株铃数×铃重×衣分/100算得单株皮棉产量; 亲本每个小区取2个皮棉样, F2每个组合取9个混合皮棉样在HFT9000上测定上半部平均长度(mm)、整齐度指数(%)、断裂比强度(cN tex–1)、伸长率(%)、马克隆值。分别对2018年和2019年亲本和F2每年2次重复的产量性状数据以及纤维品质性状求平均值, 分别找出每年每一性状的最大值、最小值及计算这一性状的平均值。
1.2 遗传模型与统计分析方法
将2年298个陆地棉品种(系) 2次重复测得的产量性状(铃数、铃重、衣分、单株皮棉产量)数据和品质性状(纤维长度、整齐度、马克隆值、强度和伸长率)以每个小区每个性状的平均值采用加性-显性与环境互作的遗传模型进行遗传方差分解和遗传效应估计。表型值可以分解为:
=+E+A+D+AE+DE+
式中,为群体均值, E为环境效应, E~N (0, VE); A为加性效应, A~N (0, VA); D为显性效应, D~N (0, VD); AE为加性×环境互作效应, AE~N (0, VAE); DE为显性×环境, DE~N (0, VDE);为剩余,~N (0, Vε)。各项遗传参数采用朱军的软件及分析方法进行计算[16-18],运用MINQUE(1)法估算方差分量, 用Jackknife抽样方法计算各遗传参数的估计值及其标准误,测验法对遗传参数作统计检验。用R (4.1.0)[19]factoextra软件包将298个陆地棉品种(系)的加性效应和显性效应进行聚类分析(结合Ward聚类和“euclidean”)。
2 结果与分析
2.1 陆地棉参试亲本及F2组合产量和纤维品质性状的遗传分析
2.1.1 产量和纤维品质性状的表型分析 将亲本和884个F2组合4个产量性状和5个纤维品质性状在2年的最大值、最小值和平均值结果列于表1。由表1可知, 亲本9个性状除伸长率外, 2019年均优于2018年。2018年F2组合的铃重、纤维长度、整齐度、马克隆值和纤维强度均优于同一环境亲本的相应性状。2019年F2组合的铃重、单株皮棉产量、纤维长度、整齐度、马克隆值以及伸长率均优于同一环境亲本的相应性状。结合2年F2的平均值来看,铃重、纤维长度、整齐度和马克隆值均优于同一环境的亲本。这些性状说明陆地棉F2可能具有杂种优势以及基因型与环境互作。
2.1.2 产量和纤维品质性状的遗传方差分析和遗传效应估计 将供试材料4个产量性状和5个纤维品质性状2年的遗传组分方差占表现型方差的比率及遗传率列于表2。由表2可以看出, 在加性效应对表现型总变异的贡献中除强度和马克隆值的加性方差比率为0外, 其他7个性状的加性方差都达到了显著和极显著水平。其中, 伸长率的加性方差对表型变异的贡献最大, 为8%; 单株铃数和纤维长度次之, 为6%。铃重和整齐度的加性方差对表现型总变异的贡献相对较低。在显性效应对表现型总变异的贡献中除单株铃数和伸长率为0外, 其他6个性状的显性效应方差比率均达到了极显著水平。其中, 衣分和马克隆值的显性方差占表现型方差的比例最大, 达73%。加性×环境互作对表型变异的贡献均存在显著的比例, 伸长率的显性×环境互作方差比率所占比例最大, 为33%、其次是单株铃数和铃重所占比例为17%, 最小的是整齐度为1%。各性状的剩余方差所占表现型方差的比例均达到了极显著水平, 其中单株铃数的剩余方差为78%。9个性状的互作广义遗传力均达到了极显著水平, 其中单株皮棉产量的互作广义遗传力最大, 为27%, 衣分的互作广义遗传力最低为6%。
2.2 陆地棉参试亲本主要性状的遗传效应分析
2.2.1 部分参试亲本产量及纤维品质性状的加性效应分析 对参试亲本产量性状和纤维品质性状遗传效应的分析能了解各亲本的应用价值。表3从附表3中随机选取了298个参试亲本中的24个亲本的7个主要性状加性效应的估计值。由表3可知, 亲本223-28、h288、337-5和15-1993-15的纤维长度、整齐度和伸长率均具有较大的正向显著或极显著的加性效应, 用其做亲本较易获得3个品质性状均高的后代。其中亲本15-1993-15在长度(1.94 mm**)和单株铃数(0.17+个)具有较高的正向极显著加性效应, 可用来改良阿拉尔地区陆地棉后代的纤维长度和铃数。亲本银99 (0.49%+)的衣分具有较高的显著的加性效应, 可作为提高后代衣分的亲本。
2.2.2 部分参试亲本产量及纤维品质性状的显性效应分析 表4从附表4中随机选取了298个参试亲本中的24个亲本的7个主要性状显性效应的估计值。从表4可知, 亲本(中36×海7124后代)衣分以外其余性状的显性效应均达显著或极显著水平。说明与该亲本杂交其后代组合在铃重、单株皮棉产量、纤维长度、整齐度、马克隆值和强度这些性状可能具有较高的杂种优势。亲本中资10号(–4.56%*)和亲本223-14的衣分(–4.36%*)存在一定的显性效应, 对其杂种优势的表现可能具有显著作用。
2.2.3 部分参试杂交组合F2各性状的显性效应分析
表5随机选取了19个组合7个主要性状的显性效应值。从表5可以看出银99×15-1993-15的单株皮棉产量、纤维长度和整齐度具有正向极显著的显性效应, 究其原因是这2个亲本的3个性状均具负向较高显性效应。223-28×246-12、223-14×银99和中资10号×h288三个组合在衣分上具有较高的显性效应。
2.2.4 部分亲本的加性×环境互作效应分析 表6随机选取了部分亲本9个性状的加性×环境互作效应值。亲本222-13在铃重、长度、整齐度和马克隆值(除外) 2年均表现出显著的正向加性×环境互作效应, 说明亲本222-13在不同的环境中在这些性状上还有不同的表现。结合2年表现来看, 2019年亲本的加性×环境互作效应略高于2018年, 原因可能是2019年的环境条件优于2018年。其他一些亲本的9个性状在不同年份具有显著不同的显性×环境互作效应。
2.2.5 部分组合的显性×环境互作效应分析 表7随机选取了部分组合4个有显著显性×环境互作效应性状的效应值。组合15-1990-5×h288在2年里的单株皮棉产量有显著的正向显性×环境互作效应, 表明在不同年份这些组合的单株皮棉产量有可能表现出正向的杂种优势, 但2018年整齐度有显著的负向显性×环境互作效应, 2019年却表现出正向显性×环境互作效应, 说明在不同年份这个组合表现出显著差异。
2.3 陆地棉参试亲本遗传效应的聚类
2.3.1 亲本加性效应的遗传聚类 对多个品种(或系)的遗传主效进行聚类可以更好地确定不同亲本的利用途径。杂交育种主要利用的是亲本的加性效应以及加性×加性上位性效应, 分析亲本的加性效应就可以知道亲本在不同性状遗传上的优缺点, 进而有针对性地选配亲本。而采用碎石图可以确定多个亲本多个性状加性效应的聚类数目, 并进行聚类就可以明确不同类亲本在杂交育种中的利用价值。对298个品种(系) 7个性状的加性效应进行聚类的结果如图1和图2。从图2与表8中可以看出, 第8类包括13份材料(222-13、223-14、1048等), 其纤维长度(2.90 mm)、整齐度(2.71%)、伸长率(1.41%)具有很高的加性效应。因此, 在这类品种(系)中挑选亲缘关系远的亲本间杂交, 在这些品种(系)的后代中容易获得品质性状很好的后代。第4类亲本包括25个品种(系) (新陆中37号、苗宝21、新陆早30号等), 这类品种(系)的铃数(0.89个/株)、铃重(1.06 g)、衣分(0.52%)和单株皮棉产量(1.47 g)具有较高的加性效应。因此, 通过这些品种(系)间的杂交, 在这些品种(系)的后代中获得产量性状较高的后代。结合7个主要性状的平均值, 第2类品种(系)较高, 可通过这类品种(系)间的杂交获得产量性状和品质性状兼优的后代。其他品种的加性效应聚类如表9所示。


表3 部分亲本7个主要性状的加性效应
+、*、**分别表示达到0.10、0.05、0.01概率显著水平。
+,*, and**mean significance difference at the 0.10, 0.05, and 0.01 probability levels, respectively.
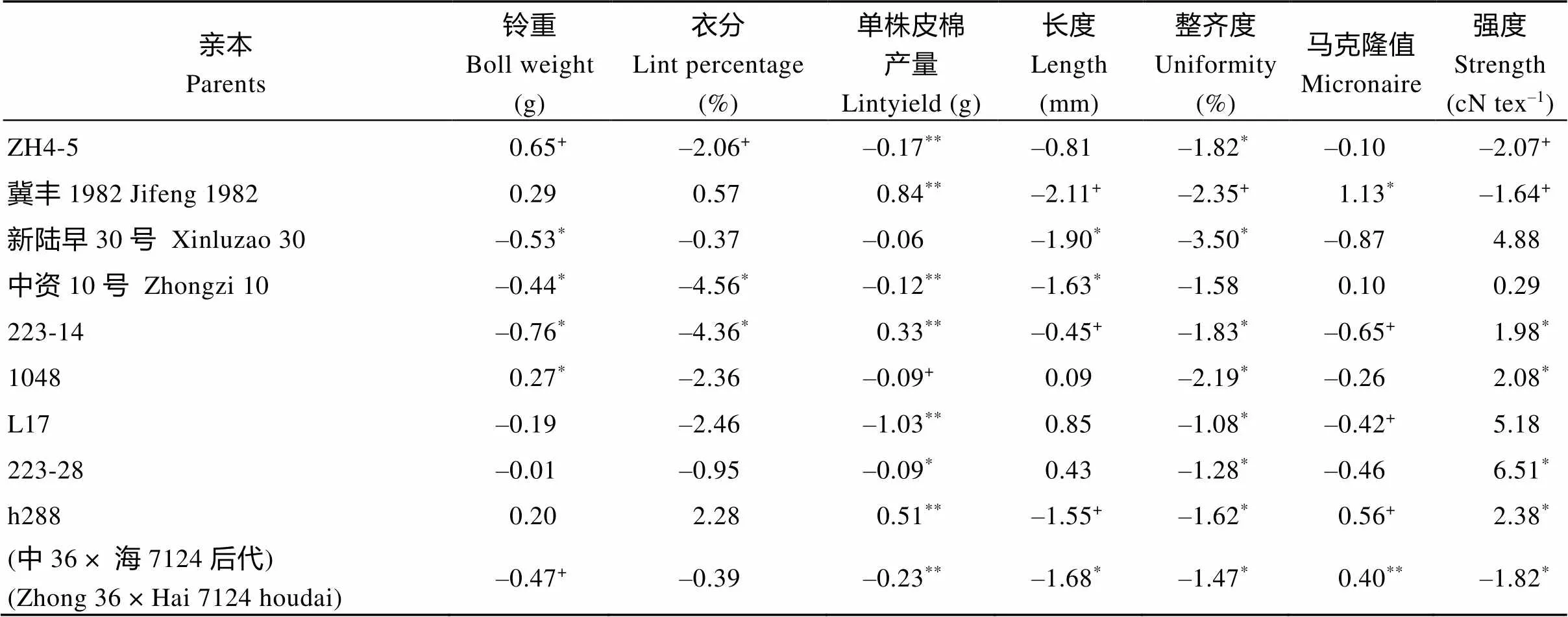
表4 部分亲本6个主要性状的显性效应

(续表4)
+、*、**分别表示达到0.10、0.05、0.01概率显著水平。
+,*, and**mean significance difference at the 0.10, 0.05, and 0.01 probability levels, respectively.

表5 部分组合8个主要性状的显性效应

(续表5)
+、*、**分别表示达到0.10、0.05、0.01概率显著水平。
+,*, and**mean significance at the 0.10, 0.05, and 0.01 probability levels, respectively.

表6 部分亲本的加性×环境互作效应
+、*、**分别表示达到0.10、0.05、0.01概率显著水平。
+,*, and**mean significance difference at the 0.10, 0.05, and 0.01 probability levels, respectively.

表7 部分组合的显性×环境互作效应
+、*、**分别表示达到0.10、0.05、0.01概率显著水平。
+,*, and**mean significance difference at the 0.10, 0.05, and 0.01 probability levels, respectively.

表8 298个品种(系) 7个性状加性效应聚类后各亲本加性效应均值
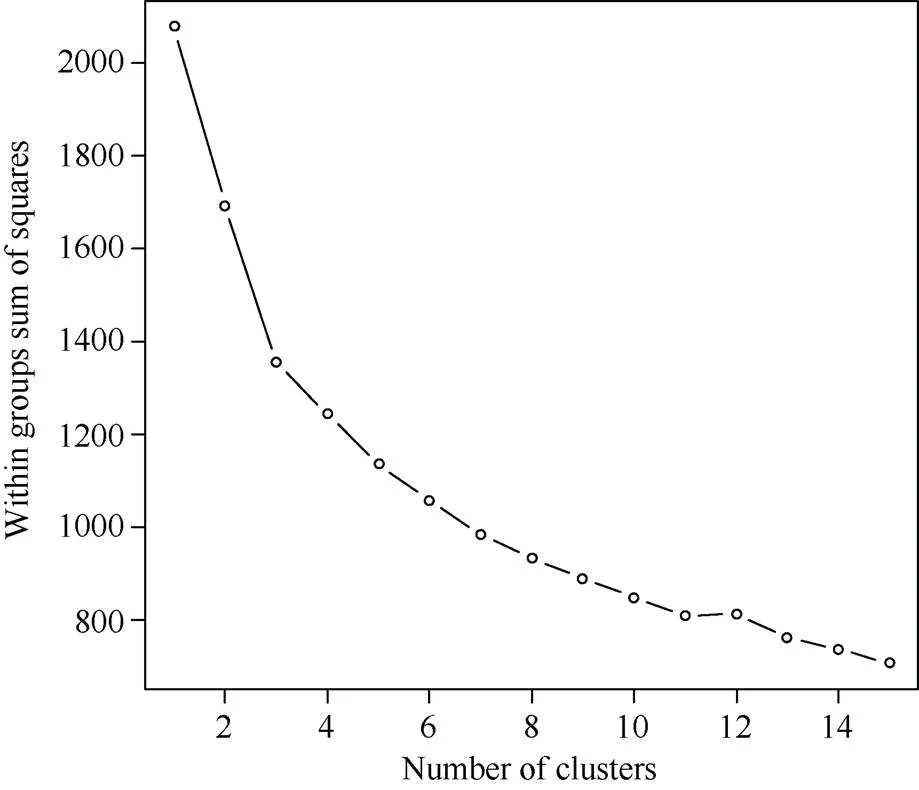
图1 亲本主要性状加性效应聚类数的碎石图

图2 298个品种(系)的加性效应聚类图

表9 12类的品种(系)构成

(续表9)
2.3.2 亲本显性效应的遗传聚类 对298个品种(系) 6个主要性状的显性效应进行分类和聚类的结果如图3和图4。从图3与表10中可以看出, 第8类品种(系)包括8份材料(中资10号、223-13、223-14等), 其铃重(-1.79 g)、衣分(-2.66%)和单株皮棉产量(-0.67 g 株-1)均具有较高的显性效应。因此, 通过这些品种(系)间的杂交可能在这些品种(系)的后代中获得这些性状较高的杂种优势。第3类的16份材料(15-1990-7、新陆中36号、j206-5等)的4个纤维品质性状均具有较高的显性效应, 通过这些亲本间的杂交较易获得长度、整齐度、马克隆值和强度较高的杂种优势, 其他品种的显性效应聚类如表11所示。
3 讨论
3.1 陆地棉参试亲本及F2组合产量和纤维品质性状的遗传分析
棉花的产量性状和纤维品质性状均属于复杂的数量性状, 它们同时受基因的加性效应和显性效应及其与环境互作效应的控制。本研究结果表明, 单株铃数和伸长率的加性效应均达到了极显著水平, 而它们的显性效应均不显著, 表明F2的这2个性状在遗传主效上受加性效应控制; 马克隆值和强度的显性效应具有很高且极显著的方差比例, 而不存在加性方差, 表明F2的这2个性状在遗传主效上受显性效应控制, 并可以在杂优育种中可以利用; 铃重、衣分、纤维长度和整齐度的加性方差和显性方差均达到了极显著水平, 并且显性方差的比例比加性方差的比例大很多, 说明在这些性状的F2表现中显性效应比加性效应起到更重要的作用。大量研究表明, 环境因素对陆地棉产量性状和纤维品质性状影响较大, 这与本研究的所有性状加性方差×环境方差/表型方差比例均达到极显著以及部分性状的显性方差比例达到极显著的结果接近, 说明加性效应和显性效应的表现存在着年份间的差异。在本研究中, 各性状的剩余方差所占表现型方差的比例均达到了极显著水平, 也与多数研究结果一致[20-22], 所以在育种过程中应注意改进田间管理措施, 提高试验数据的准确度, 降低非遗传因素的影响。9个性状的互作狭义遗传力均达到显著和极显著水平, 表明这些性状的加性效应在阿拉尔垦区内两年间差异较为明显。
3.2 陆地棉参试亲本遗传效应的聚类分析
加性效应是能够稳定遗传的遗传效应, 通过选择在产量和纤维品质性状加性效应能够互补的2类品种(系)间进行杂交, 可以获得在这些性状上表现好的后代。而选择产量和纤维品质性状加性效应兼优的同类品种(系)间杂交要比性状互补的2类品种(系)杂交获得较好后代的几率大。聚类分析可以按性状相似程度划分类别, 这种方法已经被广泛应用在前人的研究里[23-25]。本研究依据加性效应对对品种(系)聚类的结果表明, 第4类品种(系)的产量性状的加性效应较高, 若改良产量性状可以在第4类品种(系)间进行杂交; 第8类品种(系)的纤维品质性状加性效应较高, 若提高纤维品质性状可以在第8类品种(系)间进行杂交, 第4类和第8类品种(系)进行杂交可获得这些性状都好的后代。通过对品种(系)的显性效应聚类表明, 第8类品种(系)的亲本7个性状的显性效应均值较高, 可利用杂种优势。第3类的纤维品质的显性效应较高, 第8类产量性状的显性效应较高。由于基因间的加性效应可以稳定遗传, 在选择的过程中可以累加, 因此选择产量性状和纤维品质性状加性效应兼优的同类品种(系)间杂交易获得好的后代。

表10 298个品种(系) 7个性状显性效应聚类后各亲本加性效应均值

表11 9类的品种(系)构成

(续表11)

图3 亲本主要性状显性效应聚类数的碎石图

图4 298个品种(系)的显性效应聚类图
4 结论
本研究通过298个陆地棉品种(系)的F2组合及其亲本进行加性-显性及其与环境互作模型分析, 根据亲本遗传主效进行了聚类, 明确了各类品种(系)的产量性状和品质性状的遗传特性, 有利于亲本在杂交育种和杂种优势育种中的利用。结果表明, 298个亲本根据加性效应被分为12类。第2类的23个品种(系)产量(除衣分外)和纤维品质性状的加性效应较高, 是对杂种后代综合性状都有利的一类亲本; 第8类品种(系)的纤维品质性状的加性效应最好, 第4类品种(系)的产量性状加性效应较高, 通过2类品种间杂交可以获得产量性状和纤维品质性状互补的后代。298个亲本根据显性效应被分成了9类。第8类包括8个品种(系), 其产量性状和纤维品质性状(除马克隆值外)的显性效应较高, 可作为在F1和F2综合利用产量性状和纤维品质性状杂种优势兼优的一类亲本。产量性状的显性效应平均值较高的是第8类, 第3类亲本纤维品质性状的显性效应较高。
附表 请见网络版: 1) 本刊网站http://zwxb.chinacrops. org/; 2) 中国知网http://www.cnki.net/; 3) 万方数据http://c.wanfangdata.com.cn/Periodicalzuowxb.aspx。
[1] 王俊铎, 梁亚军, 龚照龙, 艾先涛, 郭江平, 买买提·莫明, 李雪源, 赵素琴, 郑巨云. 新疆植棉区2019年棉花种业报告. 棉花科学, 2021, 43(1): 3–10.
Wang J D, Liang Y J, Gong Z L, Ai X T, Guo J P, Maimaiti M, Li X Y, Zhao S Q, Zheng J Y. Cotton seed industry report in Xinjiang cotton planting area in 2019., 2021, 43(1): 3–10 (in Chinese with English abstract).
[2] Sahar A, Zafar M M, Razzaq A, Manan A, Haroon M, Sajid S, Rehman A, Mo H J, Ashraf M, Ren M Z, Shakeel A, Yuan Y L. Genetic variability for yield and fiber related traits in genetically modified cotton., 2021, 4: 219–228.
[3] Mei Y J, Guo W F, Fan S L, Song M Z, Pang C Y, Yu S X. Analysis of decision-making coefficients of the lint yield of upland cotton (L.)., 2014, 196: 95–104.
[4] Clement J D, Constable G A, Stiller W N, Liu S M. Negative associations still exist between yield and fiber quality in cotton breeding programs in Australia and USA., 2012, 128: 1–7.
[5] 喻树迅, 范术丽, 王寒涛, 魏恒玲, 庞朝友. 中国棉花高产育种研究进展. 中国农业科学, 2016, 49: 3465–3476.
Yu S X, Fan S L, Wang H T, Wei H L, Pang C Y. Advances in high yield breeding of cotton in China., 2016, 49: 3465–3476 (in Chinese with English abstract).
[6] Mei Y J, Yu J W, Xue A L, Fan S L, Song M Z, Pang C Y, Pei W F, Yu S X, Zhu J. Association mapping of genetic network for plant morphological traits in cotton.(Agric Life Sci), 2016, 42(2): 127–136.
[7] 郭介华, 邹礼平. 陆地棉12个农艺性状的基因效应估计. 棉花学报, 1994, 6: 160–162.
Guo J H, Zou L P. Gene effect estimation of 12 agronomic traits in upland cotton., 1994, 6: 160–162 (in Chinese with English abstract).
[8] Roy U, Paloti M C, Tigga A, Patil R S. Genetic variability studies in the F2populations of interspecific cotton (L. ×L.) hybrids., 2019, 11: 660–663.
[9] 王保勤, 刘书梅, 李宾, 李俊文, 韩艳红, 贾新合. 棉花永久F2群体纤维品质性状的遗传分析. 湖北农业科学, 2016, 55: 5758–5760.
Wang B Q, Liu S M, Li B, Li J W, Han Y H, Jia X H. Analysis of genetic effects for fiber quality in upland cotton immortalized F2populations., 2016, 55: 5758–5760 (in Chinese with English abstract).
[10] 代勇强, 张新宇, 孙杰. 北疆棉花三系主要选育指标的遗传效应分析. 新疆农业科学, 2017, 54(1): 1–9.
Dai Y Q, Zhang X Y, Sun J. Genetic effect analysis of main breeding targets of three lines hybrids cotton in northern Xinjiang., 2017, 54(1): 1–9 (in Chinese with English abstract).
[11] 秦鸿德, 冯常辉, 张友昌, 别墅, 张教海, 夏松波, 王孝刚, 王琼珊, 蓝家样, 陈全求, 焦春海. 基于部分NCII设计的陆地棉F1表现预测. 中国农业科学, 2021, 54: 1590–1600.
Qin H D, Feng C H, Zhang Y C, Bie S, Zhang J H, Xia S B, Wang X G, Wang Q S, Lan J Y, Chen Q Q, Jiao C H. F1performance prediction of upland cotton based on partial NCII design., 2021, 54: 1590–1600 (in Chinese with English abstract).
[12] 刘建平, 梅拥军, 张利莉, 胡守林, 郭伟锋, 熊仁次. 零式果枝海岛棉铃部性状和纤维品质的遗传及相关分析. 作物学报, 2005, 31: 1069–1073.
Liu J P, Mei Y J, Zhang L L, Hu S L, Guo W F, Xiong R C. Analyses of heredity and correlation between boll traits and fiber quality traits in “0” plant type island cotton., 2005, 31: 1069–1073 (in Chinese with English abstract).
[13] 梅拥军, 张改生, 叶子弘, 曹新川. 海岛棉零式果枝与长果枝品种间杂交F1目标产量性状的决策分析. 作物学报, 2004, 30: 1164–1168.
Mei Y J, Zhang G S, Ye Z H, Cao X C. The decision analysis on the aim yield traits of F1between “0” and “Long fruit branch” plant type in island cotton., 2004, 30: 1164–1168 (in Chinese with English abstract).
[14] 梅拥军, 张改生, 叶子弘, 曹新川, 张文英. 海岛棉不同果枝品种间杂交产量性状的遗传及F1和F2群体优势分析. 作物学报, 2004, 30: 1026–1030.
Mei Y J, Zhang G S, Ye Z H, Cao X C, Zhang W Y. Genetic analysis of yield traits and population heterosis for F1and F2between different Fruit-Branch type cultivars in island cotton., 2004, 30: 1026–1030 (in Chinese with English abstract).
[15] 梅拥军, 张改生, 叶子弘, 曹新川, 张文英. 海岛棉不同果枝品种间杂交纤维品质性状的遗传及F1和F2群体优势分析. 作物学报, 2004, 30: 796–800.
Mei Y J, Zhang G S, Ye Z H, Cao X C, Zhang W Y. Genetic Analysis of fiber traits and population heterosis for F1and F2between different Fruit-Branch type cultivars in island cotton., 2004, 30: 796–800 (in Chinese with English abstract).
[16] Zhu J. Mixed model approaches for estimating genetic variances and covariances., 1992, 1: 1–11.
[17] 朱军. 一般遗传模型和定量特征的新分析方法. 浙江农业大学学报, 1994, 20: 551–559.
Zhu J. General genetic models and new analysis methods for quantitative characteristics., 1994, 20: 551–559 (in Chinese with English abstract)
[18] 朱军. 遗传模型分析方法. 北京: 中国农业出版社, 1997. pp 56–87, 175–191.
Zhu J. Analysis Methods for Genetic Models. Beijing: China Agriculture Press, 1997. pp 56–87, 175–191 (in Chinese).
[19] Kassambara A, Mundt F. factextra: Extract and Visualize the Results of Multivariate Data Analyses. R package version 1.0.7. 2020. https://CRAN.R-project.org/package=factoextra.
[20] 詹有俊, 杨涛, 孙建船, 庄生仁, 任福成, 李渊, 张璇. 特早熟陆地棉熟性产量品质的遗传相关分析. 农业现代化研究, 2013, 34(1): 118–121.
Zhan Y J, Yang T, Sun J C, Zhuang S R, Ren F C, Li Y, Zhang X. Genetic correlation analysis of early maturity yield and quality in special-early matureL.,2013, 34(1): 118–121 (in Chinese with English abstract).
[21] 孔广超, 秦利, 徐海明, 祝水金. 棉花IF2群体构建及其在纤维品质遗传和杂种优势研究中的应用. 作物学报, 2010, 36: 940–944.
Kong G C, Qin L, Xu H M, Zhu S J. Construction of lF2population and its application in studies on genetic effects and heterosis for fiber quality in upland cotton (.L.).,2010, 36: 940–944 (in Chinese with English abstract).
[22] 沈晓佳, 孙玉强, 刘芦苇, 祝水金. 转基因抗虫棉纤维品质性状的遗传分析. 棉花学报, 2009, 21(3): 163–167.
Shen X J, Sun Y Q, Liu L W, Zhu S J. Analysis of genetic effects for fiber quality in transgenic insect resistant cotton (.L.)., 2009, 21(3): 163–167 (in Chinese with English abstract)
[23] 孙长法, 赵晖, 陈荣江. 棉花新品种产量品质性状的聚类分析与综合评价. 西北农业学报, 2010, 19(4): 77–81.
Sun C F, Zhao H, Chen R J. Cluster analysis and comprehensive evaluation on yield and quality characters of new cotton varieties., 2010, 19(4): 77–81 (in Chinese with English abstract).
[24] 董承光, 李成奇, 李生秀, 周小凤, 马晓梅, 肖光顺, 李保成. 棉花种质资源主要农艺性状的综合评价及聚类分析. 新疆农业科学, 2011, 48: 425–427.
Dong C G, Li C Q, Li S X, Zhou X F, Ma X M, Xiao G S, Li B C. Comprehensive evaluation of the major agronomic traits and cluster analysis of cotton germplasm resources.,2011, 48: 425–427 (in Chinese with English abstract).
[25] 罗海华, 邵德意, 陈功, 徐秀敏, 高欣, 袁长凯, 彭金剑, 汤飞宇. 陆地棉常规品种(系)与杂交组合性状相关性的比较分析. 作物杂志, 2017, (5): 31–37.
Luo H H, Shao D Y, Chen G, Xu X M, Gao X, Yuan C K, Peng J J, Tang F Y. Comparative analysis of trait correlation between conventional varieties (lines) and hybrids of cotton.,2017, (5): 31–37 (in Chinese with English abstract).
Genetic analysis of F2generation of upland cotton hybrids and main effect clustering in Southern Xinjiang, China
GUO Hong1, YU Ji-Wen2, PEI Wen-Feng2, GUAN Yong-Hu1, LI Hang1, LI Chang-Xi1, LIU Jin-Wei1, WANG Wei1, WANG Bao-Quan2, and MEI Yong-Jun1,*
1College of Agriculture, Tarim University, Alar 843300, Xinjiang, China;2State Key Laboratory of Cotton Biology / Institute of Cotton Research, Chinese Academy of Agricultural Sciences, Anyang 455000, Henan, China
Four yield traits and five fiber quality traits of their 298 varieties (lines) and their 884 F2crosses of upland cotton were analyzed for their additive and dominance effects by a genetic model with additive, dominance, and their interaction effects with the environment in two successive years in Alar, South Xinjiang. To explore the potential of the breeding and utilization value of varieties, all varieties were clustered using cluster analysis of the R software package based on the additive and dominance effects. The results indicated that under the high-density planting mode of “low plant growth (plant height: 0.8–1.0 m), high density (225,000–300,000 plant hm–2), early ripe and film cover” in southern Xinjiang province, the additive effects of 298 parents were divided into 12 groups. The average additive effects of yield and fiber quality traits of 23 varieties (lines) in the sixth group were in a good level. It was easy to obtain offspring with both yield (except lint percentage) and fiber quality traits through crossing between these varieties (lines). The eighth group had a better average additive effect on fiber quality traits, while the fourth group had a higher average additive effect on yield traits. The progeny with complementary yield and fiber quality traits could be obtained by crossing these two kinds of varieties. The dominance effects of 298 parents were divided into nine groups. The eighth group included eight varieties (lines) and their yield traits and fiber quality traits (except micronaire) were all at a better average level in dominance effect, indicating that could be used as parents for hybrid utilization of both yield traits and fiber quality traits. The average dominance effect of yield traits was higher in the eighth category, and the average value of dominance effect of fiber quality traits in the third group was higher.
upland cotton; F2population; yield traits; fiber quality traits; additive-dominance model; clustering analysis
10.3724/SP.J.1006.2023.24008
本研究由国家自然科学基金项目“新疆南疆陆地棉产量和纤维品质性状QTS的全基因组挖掘” (31560408)和棉花生物学国家重点实验室开放课题基金项目(CB2021A28)资助。
This study was supported by the National Natural Science Foundation of China “Genome-wide Mining of Specific Yield Traits (QTs) in Upland Cotton from Southern Xinjiang” (31560408) and the State Key Laboratory of Cotton Biology and State Key Laboratory of Cotton Biology Open Fund (CB2021A28).
通信作者(Corresponding author):梅拥军, E-mail: xnmeiyj @126.com
E-mail: 495334380@qq.com
2022-01-05;
2022-06-07;
2022-07-04.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20220701.1835.006.html
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
