温度和光照对缘管浒苔光合作用的短期交互影响
2023-01-14刘宣宏周玲洁蔡春尔洪彦杰邵慧婷何培民
刘宣宏,周玲洁,蔡春尔,2* ,洪彦杰,邵慧婷,何培民,2
(1.上海海洋大学 海洋生态与环境学院,上海 201306; 2.上海海洋大学 水产科学国家级实验教学示范中心,上海 201306)
自2007年起,中国黄海海域浒苔绿潮连年大量暴发,江苏省如东海区鉴定出附着和漂浮的浒苔属优势种为浒苔Ulvaprolifera、扁浒苔Ulvacompressa、缘管浒苔Ulvalinza和曲浒苔Ulvaflexuosa[1-2]。其中,缘管浒苔作为绿潮暴发早期的重要物种,广泛分布于中国渤海、黄海和东海海域[3]。光合作用对藻类的生长和繁殖至关重要,其反应过程复杂[4],与温度、pH及营养盐等环境因素密切相关[5-6],其中,温度和光照是最主要的制约因素[7]。由于每种绿潮藻的最适温度和光照不同[8],黄海漂浮绿潮藻会随季节出现演替现象,优势种由扁浒苔、缘管浒苔和曲浒苔逐渐演变为浒苔[1-2]。
在不同温度下,藻类通过调整色素含量和光合色素/光保护色素的比例[9]等一系列机制来调整光合作用。汤文仲等[10]研究发现,缘管浒苔光合作用和生长的适宜温度为 15~25 ℃,当温度达到5、35 ℃时,缘管浒苔的光化学效率(Fv/Fm)最小。藻体对光照变化也有不同的响应,低光照下藻体光合作用变弱,净光合效率降低[11],而高光照时容易造成藻体光抑制[12],且随高光照处理时间延长,缘管浒苔的Fv/Fm值逐渐降低[13]。当缘管浒苔短期处于低光照下,其叶绿素含量显著高于高、中光照下的含量,但净光合作用速率在低光照下显著降低[14]。6、7月黄海地区雨天光照低于30 μmol/(m2·s),晴天时高于300 μmol/(m2·s),当光照度为18 μmol/(m2·s)时缘管浒苔的Fv/Fm值更小[10]。
应用组学技术研究藻体,可以了解浒苔属对环境的响应。Liu等[15]证明了浒苔的碳固定途径,除了典型的C3循环外,还涉及C4循环及CO2浓缩机制。许建方等[16]研究发现, 丙酮酸磷酸双激酶活性的升高会使C4代谢途径功能加强,提升了缘管浒苔耐受高盐度、低盐度胁迫的能力。Fan等[17]通过分析养分转运蛋白在不同温度和营养物浓度下的基因表达情况,对缘管浒苔的环境广适应性进行了解释。利用转录组学研究基因表达量随环境条件的改变,短期效应更明显。Guan等[18]通过强光和干旱的急性胁迫研究发现,缘管浒苔的光捕获复合体蛋白基因ElipL1、ElipL2、Cbrx和OHP具有一定的光保护作用,LHCSR、Elip-like1和Elip-like2等蛋白基因也参与缘管浒苔的光保护机制[19-20]。因此,研究缘管浒苔不同光照和温度作用下光合作用的基因表达情况,对于进一步了解光合作用机制具有重要意义。本研究中,利用转录组学测序技术,从光合作用的角度分析了缘管浒苔对环境光照和温度变化的短期响应,以期为分析绿潮藻演替机制提供科学参考。
1 材料与方法
1.1 材料
试验用缘管浒苔于2008 年7月采自山东青岛胶州湾海区。鉴种后, 在实验室内20 ℃、100 μmol/(m2·s)最适条件[1]下进行纯种传代培养, 光照周期(D∶L)为 12∶12,采用VSE培养液,每隔7 d更换一次培养液。在该条件下长期培养理论上不会对藻体的生理响应产生驯化。
1.2 方法
1.2.1 试验设计 试验设置30 ℃(高温)、20 ℃(最适温度,对照)和4 ℃(低温)3个温度组,在每个温度下设置400 μmol/(m2·s)(高光照)、100 μmol/(m2·s)(最适光照,对照)和20 μmol/(m2·s)(低光照)3个光照度(表1)。将藻体分为9组,每组设置3个生物学重复,每个重复组取10根同一批放散后生长的缘管浒苔藻体(长度为5 cm,鲜质量为0.02 g),光照周期不变,同时处理24 h。

表1 缘管浒苔温度、光照试验设计Tab.1 Experimental design of effects of temperature and light intensity on green alga Ulva linza
1.2.2 光合作用参数测定 采用PHYTO-PAM荧光分析仪(Walz,德国)测定叶绿素荧光参数。以最适温度和最适光照为对照组,每组设置3个平行试验,处理24 h后取样,暗适应20 min后测定光合诱导曲线,并记录最大光化学效率等参数。
1.2.3 总RNA提取及转录组测序 取不同光照和温度处理24 h后的藻体,经液氮处理后,于-80 ℃超低温冰箱中保存备用。采用生工生物工程(上海)股份有限公司的RNA提取试剂盒提取总RNA。采用紫外分光光度法检测其浓度和含量,采用琼脂糖凝胶电泳检测RNA的纯度和完整性。检测合格的藻体总RNA送至华大基因公司进行文库构建和测序。采用磁珠法富集样品mRNA,经反转录后合成双链cDNA,纯化后进行末端修复、接头连接,经过产物纯化和片段大小分选,最终PCR扩增获得文库,并在Illumina HiSeqTM2000平台上完成RNA-Seq测序分析。
1.2.4 组装及功能注释 通过Trinity软件对筛选后的clean reads进行组装得到Unigene序列。随后将Unigene序列与外源蛋白数据库NR、GO、SwissProt、KEGG和InterPro进行Blastx比对(E-value<0.000 01),根据比对结果获得Unigene的蛋白功能注释信息。
1.2.5 生物信息学分析 采用FPKM法计算Unigene表达量。根据P<0.05且|log2(FoldChange)| ≥1筛选得到显著表达的Unigene。通过基因注释进一步筛选得到光合作用不同阶段起重要作用的Unigene基因,使用NCBI Blast+(http://bioinformatics. psb.ugent.be/blast/moderated/?project=orcae_Ulvmu)将每条基因与易变石莼Ulvamutabilis基因组进行比对[21],总体认为比对率为70%以上的基因与浒苔具有同源性。
1.3 数据处理
试验数据均以平均数±标准差(mean±S.D.)表示,采用SPSS 16.0软件进行单因素方差分析(one-way ANOVA)和Tukey多重比较,使用GraphPad Prism等软件绘制图表。显著性水平设为0.05。
2 结果与分析
2.1 藻体的光合特征
从图1可见:在不同光照下,缘管浒苔受低温(4 ℃)影响时,Fv/Fm值均显著降低(P<0.05);在最适温度(20 ℃)下,光照对藻体Fv/Fm的影响不显著(P>0.05),仅高光照下略有影响;在高温/低温与光照的协同作用下,高光照下藻体Fv/Fm值均显著低于低光照(P<0.05),且低温低光照下藻体Fv/Fm值显著高于单一低温。由此推测,温度和光照对缘管浒苔的交互作用影响大于单一因素,低光照缓解了低温对藻体的胁迫。
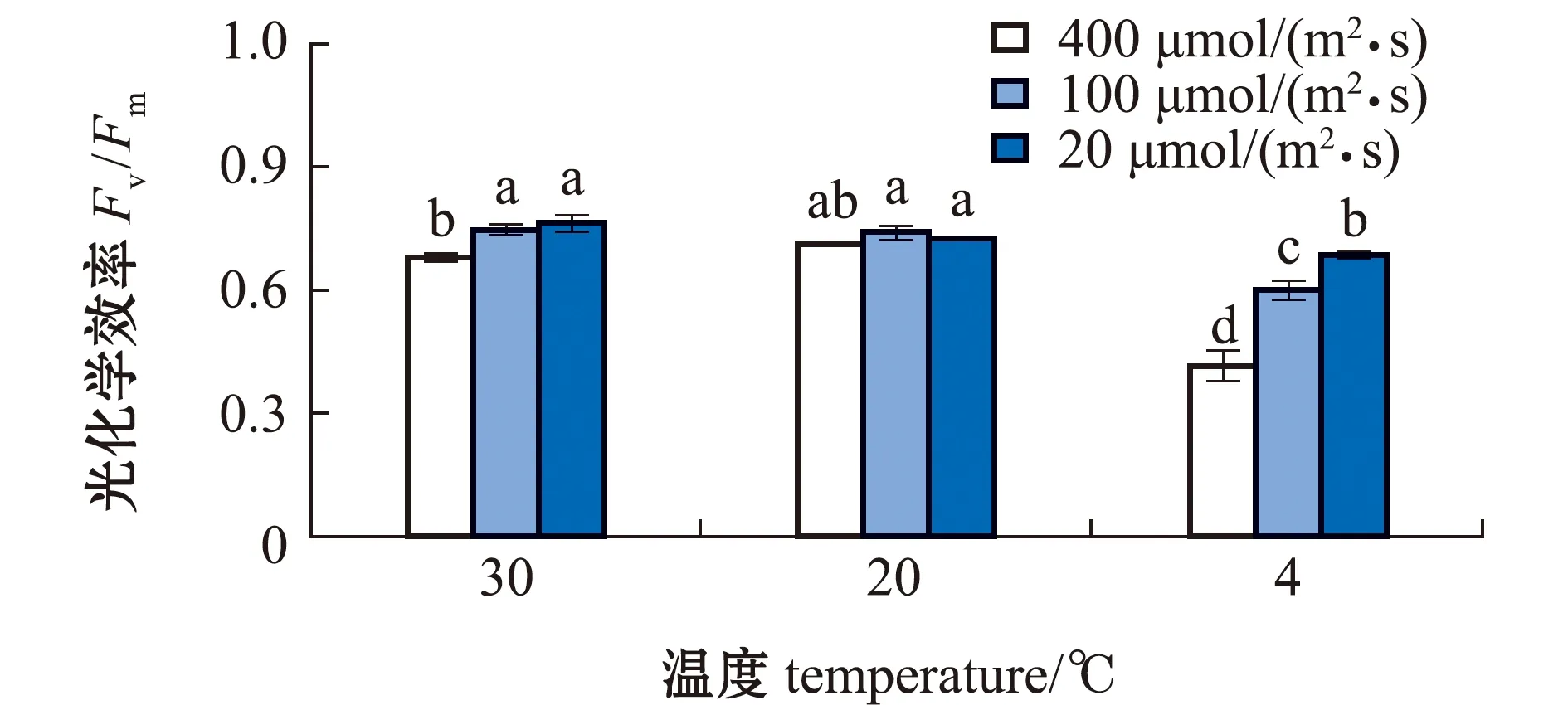
标有不同字母者表示组间有显著性差异(P<0.05),标有相同字母者表示组间无显著性差异(P>0.05)。The means with different letters are significantly different in the groups at the 0.05 probability level, and the means with the same letter are not significant differences.图1 不同处理下缘管浒苔的光化学效率Fig.1 Photochemical efficiency of green alga Ulva linza under different treatments
2.2 差异表达基因分析
通过Illumina HiSeqTM2000平台测序,总计产出42 984 321 300 nt数据。组装的总Unigene有101 056个,总长209 052 506 nt,平均长度2 069 nt,N50达到3 489 nt。Unigene功能注释得到NR、NT、Swiss-Prot、KEGG、COG和GO库的Unigene数量分别为60 840、11 036、42 042、45 915、 9 915 和26 451,去除重复后总计为61 580个。根据差异表达分析,不同处理下差异表达的基因数量如表2所示,缘管浒苔在单一的温度处理(Y2、Y8)后产生的差异表达基因数量大于单一光照(Y4、Y6)处理后。在温度和光照的交互影响下,高温高光照(Y1)和低温低光照(Y9)处理组显著差异表达的基因数量较多。
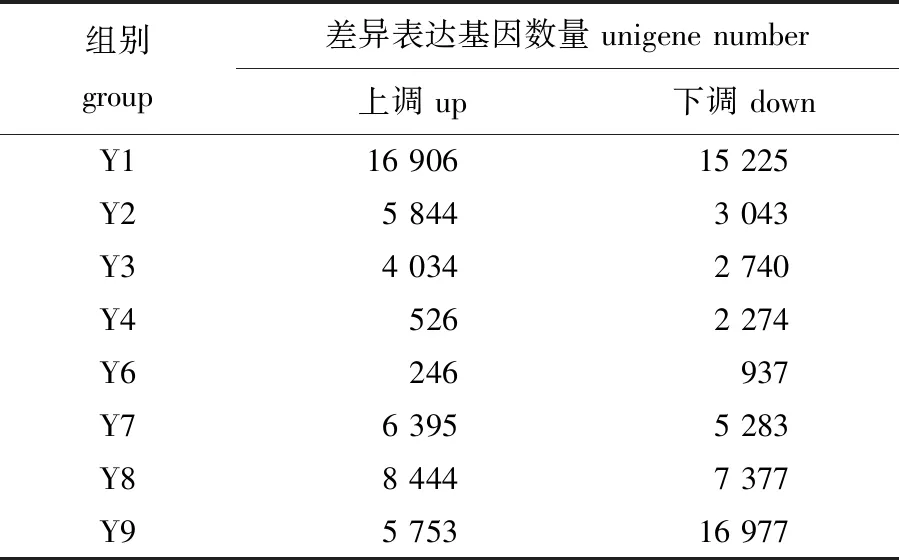
表2 所有基因表达差异比较结果Tab.2 Comparison of all differentially expressed genes
2.3 低温低光照(Y9)处理下特异表达的基因
相比其他处理条件,缘管浒苔在响应低温与低光照的短期作用时有104个Unigene特异高表达(其他情况下这些Unigene的FPKM几乎为0),其中,有36个Unigene特异基因可以注释到KEGG图上(表3),这些基因在应对温度或光照的单一胁迫时其表达量无显著性变化,且涉及翻译过程的核糖体、RNA转运、氨基酸代谢、运输过程的吞噬体及有防御功能的细胞壁蛋白。通过Uniport对这36个特异表达基因进行定位查询,发现40S核糖体蛋白S24-1、S8-1及60S核糖体蛋白L12-1这3个基因位于叶绿体中。
2.4 捕光色素蛋白基因
光合作用初期,天线色素分子吸收和传递光能。本研究中,捕光色素蛋白复合物基因表达量在不同温度和光照条件下表现出较大差异,结果如表4所示。与光系统 Ⅰ(SPⅠ)相比,光系统 Ⅱ(SPⅡ)的捕光色素蛋白受外界影响较小,捕光色素蛋白基因Lhcb6和Lhcb7在各处理条件下均未发生显著变化。捕光色素蛋白对单一温度处理(Y2/Y8)的响应大于单一光照处理(Y4/Y6)的响应;在低温条件下(Y7/Y8/Y9),捕光色素蛋白受低光照影响,低光照(Y9)处理组差异基因除Lhca1和Lhca2外,其他基因的变化趋势与低光照(Y7)和中光照(Y8)处理组相反,藻体受低温和低光照协同作用时,上调捕光色素蛋白可能用来抵御胁迫。
2.5 电子传递链相关基因
电子传递链是光合作用形成同化力的核心环节。本研究中,缘管浒苔的电子传递链相关基因响应较小,结果如表5所示。低温中光照(Y8)和低温低光照(Y9)处理组部分基因呈现上调趋势,其中,低温低光照(Y9)处理组差异表达基因的表达趋势与捕光色素一致,捕获的光能需要更多的载体进行传递。除此之外,还发现D1蛋白基因的表达量在高温高光照(Y1)、低温中光照(Y8)和低温低光照(Y9)处理组发生显著上调(P<0.05)。由此推测,在高温高光照条件下,缘管浒苔启动了SPⅡ中D1蛋白的可逆失活来耗散多余的能量,而在低温中光照和低温低光照条件下D1蛋白上调,可能是由于较低温度下酶活性不高,藻体通过提高表达量来应对低温环境。
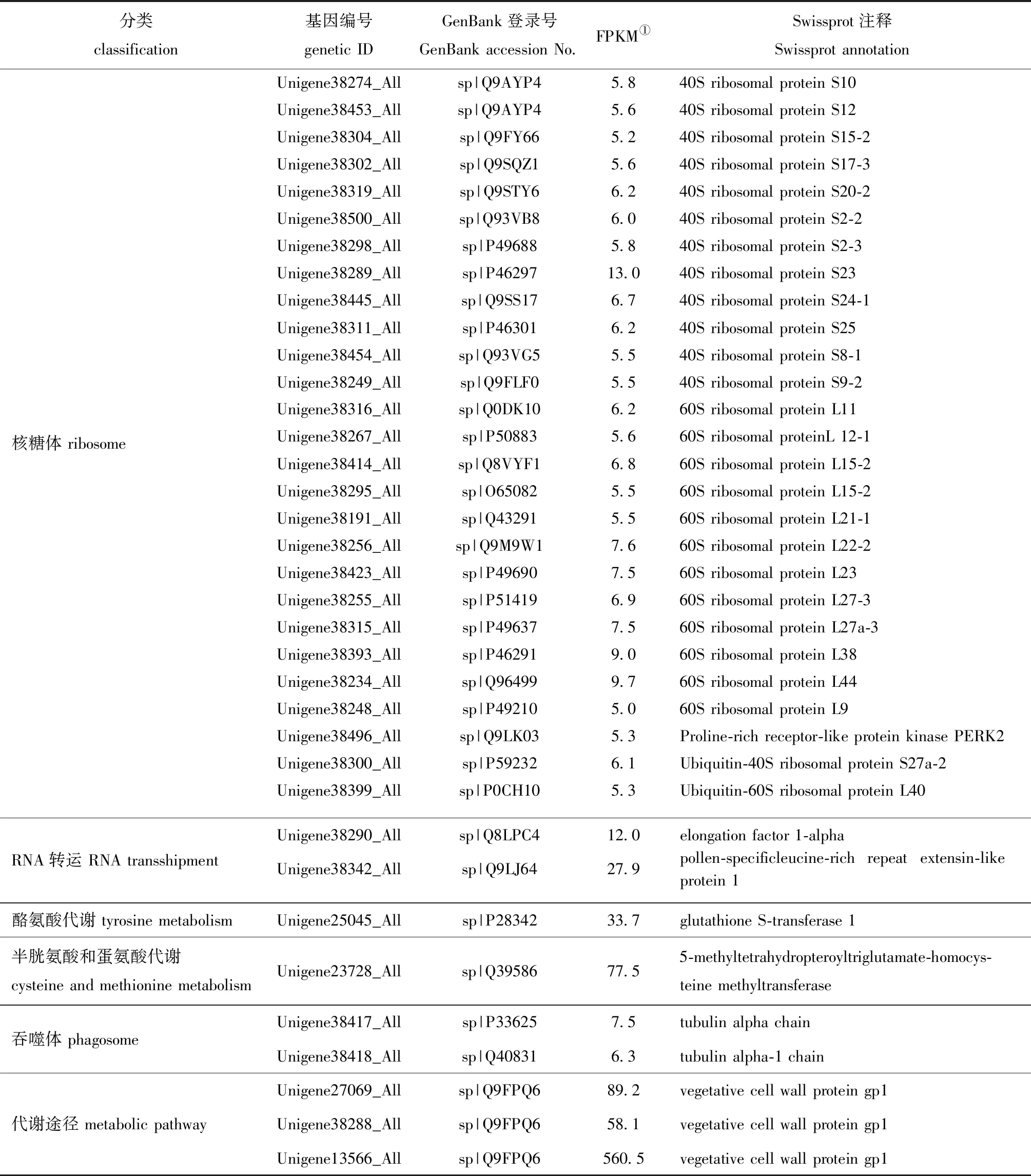
表3 低温低光照处理下特异表达的UnigenesTab.3 Unigenes specifically expressed under low temperature and low light interactions
2.6 光合碳同化相关酶基因
光合固碳C3和C4途径的相关基因检测结果如表6所示。不同处理条件下藻体的碳同化能力受到不同程度的影响,适温(Y4/Y6)条件下,碳同化相关酶基因表达情况受光照影响较小,缘管浒苔固碳反应受温度的影响大于光照;高温高光照(Y1)处理下,C3和C4途径差异表达基因均显著下调(P<0.05);低温(Y7/Y8/Y9)条件下,藻体C3和C4途径的CO2受体再生阶段酶的基因均显著下调(P<0.05)。
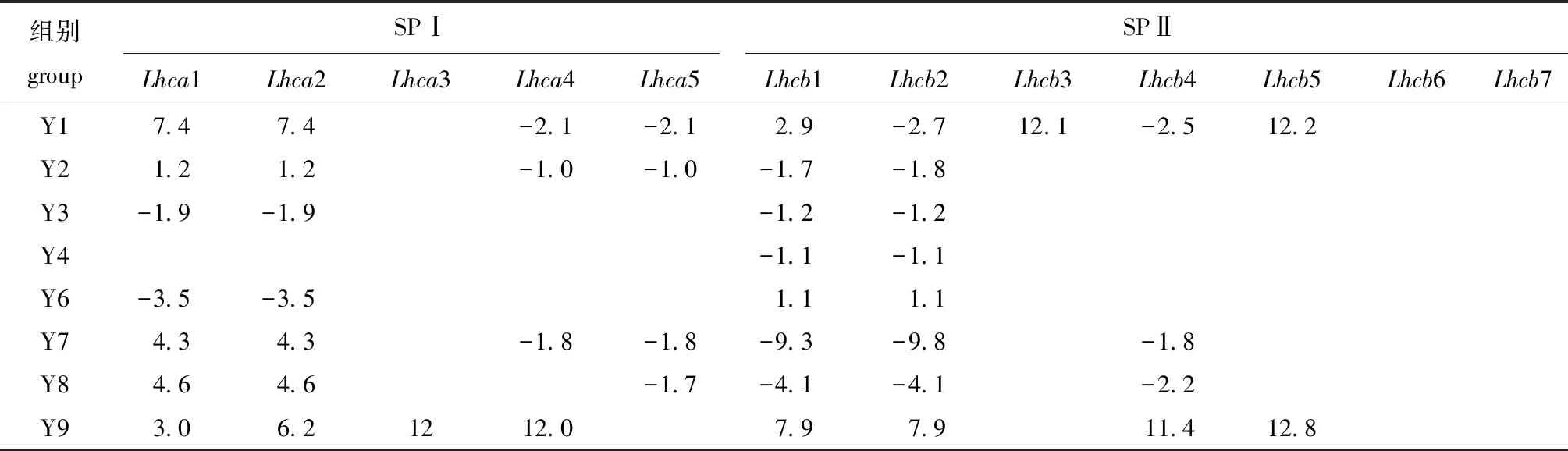
表4 不同温度和光照条件下捕光色素蛋白基因相对表达量的变化Tab.4 Relative expression level of genes related to light harvesting pigment protein under different temperature and light intensities
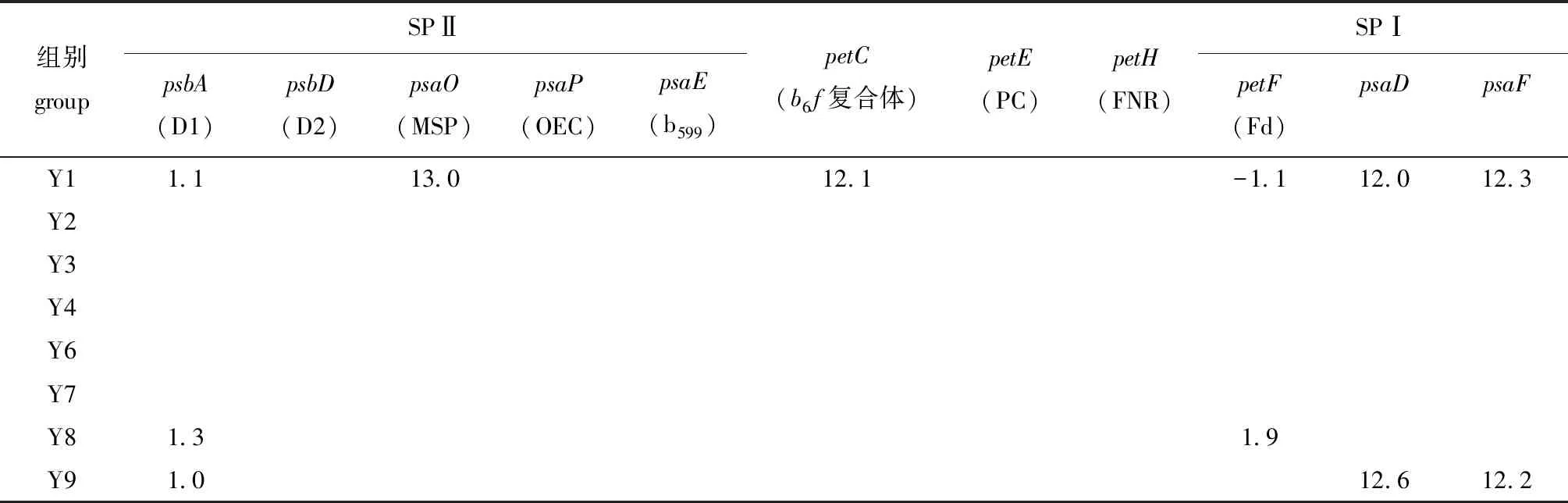
表5 不同温度和光照条件下电子传递链相关基因相对表达量的变化Tab.5 Relative expression level of genes related to electron transport chain under different temperature and light intensities

表6 不同温度和光照条件下光合碳同化相关酶基因相对表达量的变化Tab.6 Relative expression level of genes related to photosynthetic carbon assimilation under different temperature and light intensities
3 讨论
3.1 缘管浒苔的演替
大量微观繁殖体在春末海区温度回升时迅速萌发,在漂移过程中,随着温度和光照升高缘管浒苔逐渐消失[22]。王艺等[22]通过对绿潮发生过程进行全程监视,于2015年3—6月在江苏如东海区发现了缘管浒苔繁殖体。宋伟[23]通过对苏北浅滩紫菜养殖筏架上的定生绿藻进行连续跟踪,于2012年7月在青岛海域漂浮藻中发现缘管浒苔,而华梁等[24]于2012和2013年的8月在江苏紫菜筏架区绿潮藻中并未鉴定到缘管浒苔,这表明随着海水温度升高和绿潮藻群逐渐向北漂移,缘管浒苔逐渐消失,优势种逐渐由扁浒苔、缘管浒苔和浒苔演替成主要由浒苔构成的漂浮绿潮藻种群[25]。本研究中,通过24 h短期温度与光照处理,分析了缘管浒苔在适应环境变化时对光合作用的响应,根据结果推测,冬季温度较低时,藻体上调部分捕光色素蛋白基因和电子传递链基因的表达,但光合固碳反应的CO2受体再生阶段受到抑制,生长缓慢;夏季随着温度升高,藻体下调C3和C4途径的基因表达,抑制光合固碳反应来适应白天高光照环境。
3.2 藻体光合特性
光化学效率是研究环境对藻体光合作用效应的重要指标,常被用来表征植物被胁迫下的光系统变化。短期的高光照条件会引起浒苔光合能力下降。付倩倩等[14]在短期高光照(400 μmol/(m2·s))处理缘管浒苔后,其Fv/Fm值低于中光照(200 μmol/(m2·s))、低光照(90 μmol/(m2·s))处理。张珺等[26]用高光照(400 μmol/(m2·s))处理浒苔第3天后,其Fv/Fm值和实际光化学量子效率显著低于其他光照(40~160 μmol/(m2·s))处理。本研究中发现,缘管浒苔在单一高光照 (400 μmol/(m2·s))短期(24 h)处理后,其Fv/Fm值略有下降,但在高温(30 ℃)或低温(4 ℃)的协同作用下,Fv/Fm值显著下降,且藻体的差异表达基因数量也增多;在高温与高光照协同作用下,藻体光合固碳、电子传递和补光色素蛋白相关基因大量差异表达,其中光合固碳基因的表达均显著下调,这表明,缘管浒苔对温度和光照的协同作用响应更大。Jiang等[27]研究发现,高光照 (400 μmol/(m2·s))处理5 d,可以缓解高温(30 ℃)对浒苔生理参数的抑制作用,但本研究结果与之相反,这可能是本研究中缘管浒苔高光照下未能在短期内缓解受到的高温胁迫,而在较高光照下长期培养后,对温度的耐受性会增加,表现出一定的适应性。
3.3 低温低光照下缘管浒苔的响应分析
低温低光照条件是藻类越冬及复苏经历的极端环境条件。刘峰[28]研究发现,绿潮藻显微繁殖体在低温环境中可以存活3~4个月;同时Schories[29]研究认为,越冬孢子是绿潮藻显微繁殖体的重要来源,其能在黑暗条件下存活至少10个月。在大型海藻的液体保存法中,低温低光照也是一种实用的种质保存方法[30]。本研究中发现,缘管浒苔在短期(24 h)的低温低光照处理下已经产生了较强的适应性。相较于单一低温条件,低温低光照处理下的藻体Fv/Fm值反而显著增加(图1),低光照提高了缘管浒苔对低温的耐受性。此外,差异表达的捕光色素蛋白和电子传递链相关基因在低温低光照下表达量均显著上调,这可能是由于低光照诱导植物产生了更多的光合色素[11,31],捕获更多的光能,低温和低光照协同作用时藻体通过储藏更多的能量抵御胁迫。值得注意的是,藻体在低温低光照下特异高表达大量基因(表3),其中,S24-1、S8-1和L12-1 3种位于叶绿体上的核糖体蛋白基因表达极显著上调(P<0.01且|log2(FoldChange)|>2),推测这些蛋白可能参与修饰光合作用中大量上调的基因,从而使低温低光照条件下的光合作用参数Fv/Fm值高于低温高光照和低温中光照处理。由此可见,低温和低光照协同作用下,缘管浒苔存在特殊的适应机制。
4 结论
1)低温低光照下,缘管浒苔上调捕光色素蛋白和电子传递链相关基因,并特异表达部分叶绿体基因,说明藻体在冬季极端环境下具有特殊适应机制。
2)高温高光照下,藻体光合固碳作用被全面抑制,推测缘管浒苔不适应夏季极端环境。
3)温度和光照的交互影响下,藻体的光合作用参数和基因表达情况均异于单一条件下,说明温度和光照对藻体具有协同作用。
