不同种源蒙古栎种子及其苗期生长性状变异研究
2023-01-12李祉漪张家榕雷鸣雷杨立东李亚亚陆秀君张晓林
李祉漪,战 昊,张家榕,雷鸣雷,杨立东,李亚亚,陆秀君,张晓林*
(1.沈阳农业大学 林学院,辽宁 沈阳 110161;2.朝阳市森林病虫害防治检疫站,辽宁 朝阳 122000; 3.清原满族自治县国有杨树崴林场,辽宁 抚顺 113300)
蒙古栎(Quercusmongolica),又名蒙栎,柞栎,柞树,属壳斗科(Fagaceae)栎属(Quercus),是国家二级珍贵树种,在我国华北、东北10个省、市、自治区均有分布[1]。其木材坚硬耐腐,可作为建筑材料,叶片和树皮有很高药用价值[2],是用途广泛的经济型树种。同树种不同分布区的遗传特性有很大差异,这种与地理分布相联系的变异,称之为地理变异[3]。地理变异表现在植物的表型性状和生理性状等方面,种源试验则是通过观察,发现优良性状与地理种源的变异规律,从而做到林木遗传性状改良[4]。表型性状观测是研究植物遗传多样性最直观简单的方法[5],生理性状需要借助特定仪器进行测量分析。关于蒙古栎的种源试验,我国已有学者在吉林省不同生态区11个种源采种,对树高、地径等生长指标进行测量,最终选出各生态区的优良种源[6]。在不同种源蒙古栎生理特征差异方面,有学者选取5个蒙古栎种源,对其苗木叶片的气体交换特征、栅栏组织厚度和光合色素进行测定,发现不同种源叶片结构和光合特性存在差异,并选出江密峰种源应用潜力最大[7]。也有对其他树种如胡桃楸(Juglansmandshurica)[8]、元宝槭(Acertruncatum)[9]、东北杏(Prunusmandshurica)[10]、山桐子(Idesiapolycarpa)[11]、红栎(Quercusrubra)[12]、文冠果(Xanthocerassorbifolium)[13]等进行了种源变异的相关研究。目前,天然蒙古栎林已被过度采伐,资源的枯竭导致森林生态被破坏,相关产业资源短缺,开展蒙古栎人工林培育迫在眉睫。有针对性地推广蒙古栎人工林培育成为蒙古栎研究热点[14]。本研究通过对辽宁地区7个不同地理种源的蒙古栎种子和幼苗的表型性状试验,测量了种子长径、短径、千粒重(质量,下同)、苗高、地径、侧枝数、叶脉数、顶芽体积、生物量一系列指标,采用方差分析法处理数据,综合种子和苗期的表型性,状探索不同种源变异规律,从而为筛选生长发育良好的蒙古栎优质种源提供理论支持。
1 材料与方法
1.1 试验地及种源地概况
试验在沈阳农业大学营造的蒙古栎种源地内进行(41.481 18°N、123.253 12°E),海拔41.45 m,属于温带半湿润大陆性气候,年平均气温6.2~9.7 ℃,年降水量600~800 mm。该试验地于2019年播种,采取随机区组设计,株行距0.1 m×0.2 m,试验区四周设有同种树0.5 m宽保护行,防止边行效应,提高试验精度。
在辽宁省蒙古栎分布区收集了7个种源。种源地理分布跨度为120.303 37°-125.169 61°E、40.136 73°-42.738 35°N。分别是丹东市,西丰县,弯甸子镇,碱厂镇,草河口镇,朝阳县和普乐堡镇(表1)。
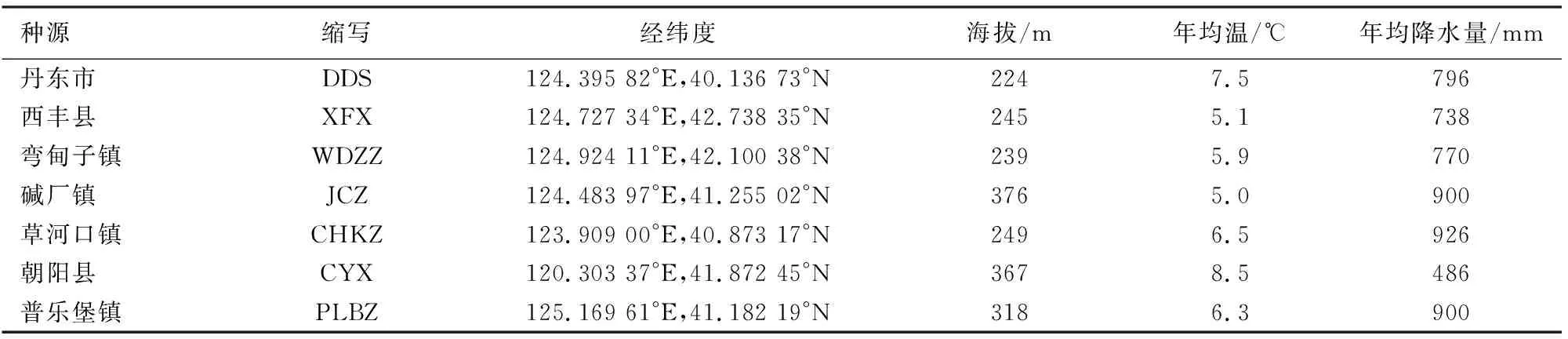
表1 各种源地名称缩写及地理位置信息
1.2 研究方法
1.2.1 种子千粒重和表型性状测定 用游标卡尺测量种子最长处和最宽处,精确到0.01 mm,作为种子长和宽数据。将各种源蒙古栎种子充分混合均匀,随机取出100粒,重复3次取平均值,折算成千粒重。
1.2.2 幼苗生长指标测定 2020年10-12月,各种源中随机选取100株长势一致的幼苗,共计700株,标号,测量苗高、地径、侧枝数、叶脉数、顶芽体积及生物量。苗高和地径分别用卷尺和游标卡尺测量,读数分别精确到0.1 cm和0.01 mm。数侧枝数。取每株幼苗顶端叶片2~4片,数叶脉数, 取平均值。借助imagej软件测算顶芽的茎和高,用圆锥体体积公式计算顶芽体积。取样后苗木用清水洗净,分为叶、茎、根与裁剪,后装入信封。放进烘箱,105 ℃下杀青30 min,70 ℃烘干至恒重。
1.2.3 数据处理 采用S-N-K多重比较法、Duncan、LSD、变异系数分析、方差分析等方法处理种子和苗期所有数据。综合评价用隶属函数综合分析法,将种源按其隶属度大小排序,选出辽宁地区优良种源。
统计软件:SPSS 22.0、Excel 2019;作图软件:SigmaPlot14.0。
变异系数计算公式:变异系数C·V=(标准偏差SD/平均值Mean)×100%。
隶属度计算公式:隶属度U(Xi)=(测定指标Xi-最小值Xmin)/(最大值Xmax-最小值Xmin)。
2 结果与分析
2.1 蒙古栎种子表型性状变异分析
对7个种源蒙古栎种子表型性状进行方差分析(表2),种间P均小于0.01,说明不同种源间蒙古栎种子各表型性状差异极显著。
对7个种源蒙古栎种子表型性状进行多重比较分析。其中,JCZ种源地长径值最大20.27 mm,与其他种源差异性显著(P<0.05)。其次为DDS种源地19.53 mm,最小值为PLBZ种源地17.63 mm。短径最大值为XFX种源地15.79 mm,较其他种源达到显著差异(P<0.05)。其次为CYX和JCZ种源地,分别为15.65 mm和15.32 mm。DDS种源地种子长宽比值最大,达到1.34,较总均值高出5.55%,较最小值种源高出12.2%。DDS、JCZ、WDZZ种源地种子长宽比显著高于其他4个种源地(P<0.05)。XFX种源地种子长宽比为最小值1.19。7个种源间千粒质量差异均显著(P<0.05),JCZ种源地值最大3 074.05 g,较千粒质量总均值高11.9%,较千粒质量最小值种源高27.4%。CHKZ种源地千粒质量最小值,2 412.66 g。
2.2 蒙古栎种子表型性状变异系数分析
对各种源蒙古栎种子表型性状进行变异系数分析(表3)。在7个种源间,长径变异系数在8.13%~13.14%,最小在XFX,最大在PLBZ。短径变异系数在8.26%~12.145%,最小在XFX,最大在WDZZ。千粒重变异系数在0.09%~0.27%,种间变异不大。总体上,XFX种源地蒙古栎种子性状变异系数最小6.07%,其次是JCZ种源地7.68%,说明2个种源的种子表型性状较为平均。CHKZ种子的种间变异系数最大18.17%,说明CHKZ蒙古栎种子存在丰富的变异。

表2 不同种源蒙古栎种子表型性状多重比较分析

表3 不同种源蒙古栎种子表型性状变异系数分析
种源内部,长径和短径的变异系数相近,而长宽比变异系数最大达到17.46%。千粒重变异系数最小0.18%,说明各种源内部千粒相对稳定。
2.3 蒙古栎苗期表型性状变异分析
对7个种源蒙古栎的苗高、地径、侧枝数、叶脉数、顶芽体积进行多重比较分析(图1-图5)。其中,XFX种源地苗高最大,达到56.88 cm,较苗高总均值高27.2%,较苗高最小种源高51.7%,说明6个种源之中XFX种源地蒙古栎生长普遍较高。其次为JCZ,苗高48.80 cm。DDS种源地苗高最小37.49 cm,说明DDS蒙古栎生长普遍较矮。CYX种源地地径最大,达到10.35 mm,较地径总均值高9.9%,较地径最小值种源高23.9%。其次为JCZ和XFX种源地,分别为10.11 mm和10.11 mm。CYX、JCZ和XFX这3个种源地苗木的地径与其他种源的地径呈现显著差异,这3个种源蒙古栎生长普遍更为粗壮。DDS种源地地径最小8.36 mm,说明DDS蒙古栎苗木生长较细。XFX种源地侧枝数最大,达到6.03个,较侧枝数总均值高18.8%,较侧枝数最小值种源高53.5%。其次为CYX、JCZ、PLBZ种源地,分别为5.85、5.73和5.38个。XFX、CYX、JCZ和PLBZ这4个种源地苗木的侧枝数显著多于其他种源地。WDZZ种源地侧枝数最小3.93个,说明WDZZ种源地蒙古栎侧枝较少。WDZZ种源地蒙古栎叶脉数最大10.26个,显著高于其他6个种源。其次为DDS种源地,平均叶脉数10.15个。CYX种源地叶脉数最小9.1个。DDS种源地顶芽体积最大18.89 mm3,较顶芽体积总均值高22.7%,较顶芽体积最小值种源高47.9%。其次为JCZ种源地,顶芽体积16.56 mm3。CYX种源地顶芽体积最小12.77 mm3。XFX种源地叶生物量均值最大达到2.49 g,显著高于其他种源,较叶生物量总均值高65.23%。其次为JCZ种源地,叶生物量均值1.96 g。JCZ种源地茎生物量和根生物量均最大,分别达到69.06 g和90.33 g,茎生物量显著高于其他种源,根生物量显著高于除XFX外的5个种源。
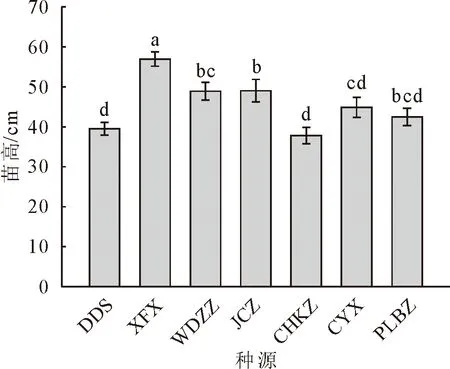
注:图中同一指标字母相同表示差异不显著,不同则表示差异极显著,下同。

图2 不同种源蒙古栎地径多重比较分析
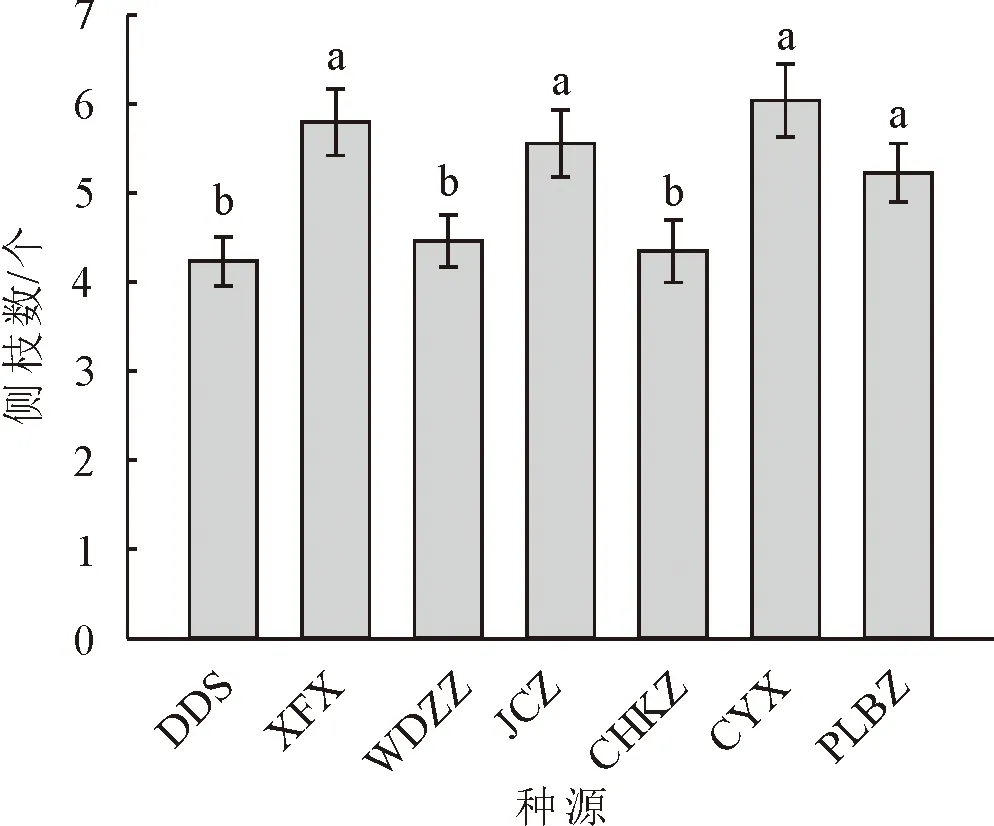
图3 不同种源蒙古栎侧枝数多重比较分析
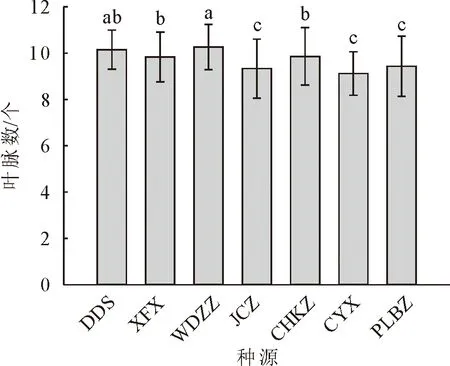
图4 不同种源蒙古栎叶脉数多重比较分析
2.4 蒙古栎苗期表型性状变异系数分析
对7个种源蒙古栎苗期表型性状进行变异系数分析(表4)。种源之间苗高变异明显,变异系数在25.96%~48.97%,XFX种源地较CYX相差近2倍,表明CYX种源地蒙古栎苗高差别较大,而XFX种源内苗高较均匀。各种源地径变异系数在23.42%~29.88%,变异不明显。CHKZ种源地侧枝数变异系数70.88%,高于其他种源。叶脉数变异系数在8.32%~13.80%,最小为DDS种源地,最大为PLBZ种源地。顶芽体积变异系数在67.36%~82.76%,种间差异较大。叶生物量、茎生物量、根生物量变异系数分别在6.05%~16.14%、11.31%~58.86%和12.07%~40.29%。总体上,JCZ种源地苗期性状变异系数最小32.37%,说明JCZ蒙古栎幼苗长势平均。而CHZK种源地变异系数最大40.48%,说明该种源蒙古栎幼苗变异明显。

图5 不同种源蒙古栎顶芽体积多重比较分析
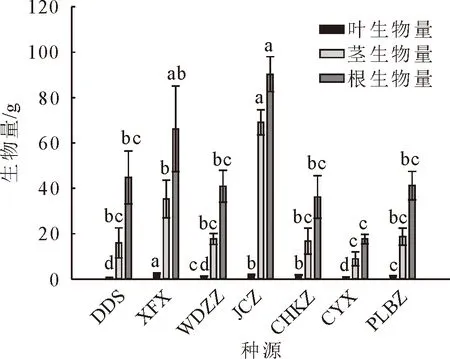
图6 不同种源蒙古栎生物量多重比较分析
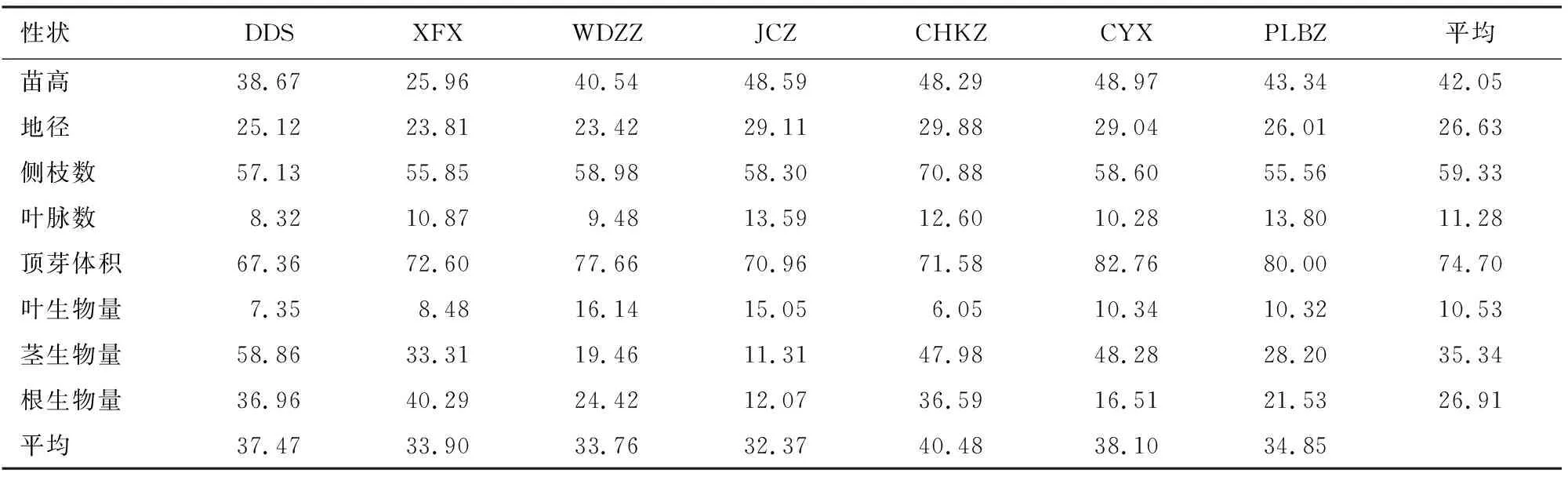
表4 不同种源蒙古栎苗期表型性状变异系数分析
种源内,所有观测性状变异系数均>5%,叶生物量平均变异系数最小10.53%,最大为顶芽体积74.70%。由此可见,蒙古栎的苗期表型性状的变异丰富,叶生物量是相对稳定的生长指标,顶芽体积变异最多。
2.5 不同种源蒙古栎种子及苗期表型性状与地理因子相关性分析
通过相关性分析可见(表5),苗高和地径2个生长指标与纬度均显著正相关,说明蒙古栎对高纬度地区气候适应性强,对低纬度地区气候适应性弱,故表现出纬度越高的种源地区,苗高地径的平均生长指标越大。叶脉数和顶芽体积与经度呈正相关,与纬度、海拔呈负相关,且叶脉数与海拔呈极显著的负相关(P<0.01),即海拔高的地区蒙古栎幼苗叶片平均叶脉数越少。叶生物量与海拔呈负相关,与经度、纬度呈正相关,但均不显著。茎生物量、根生物量与各地理因子均呈不显著正相关。种子表型性状指标与各地理因子均呈正相关,但均不显著。
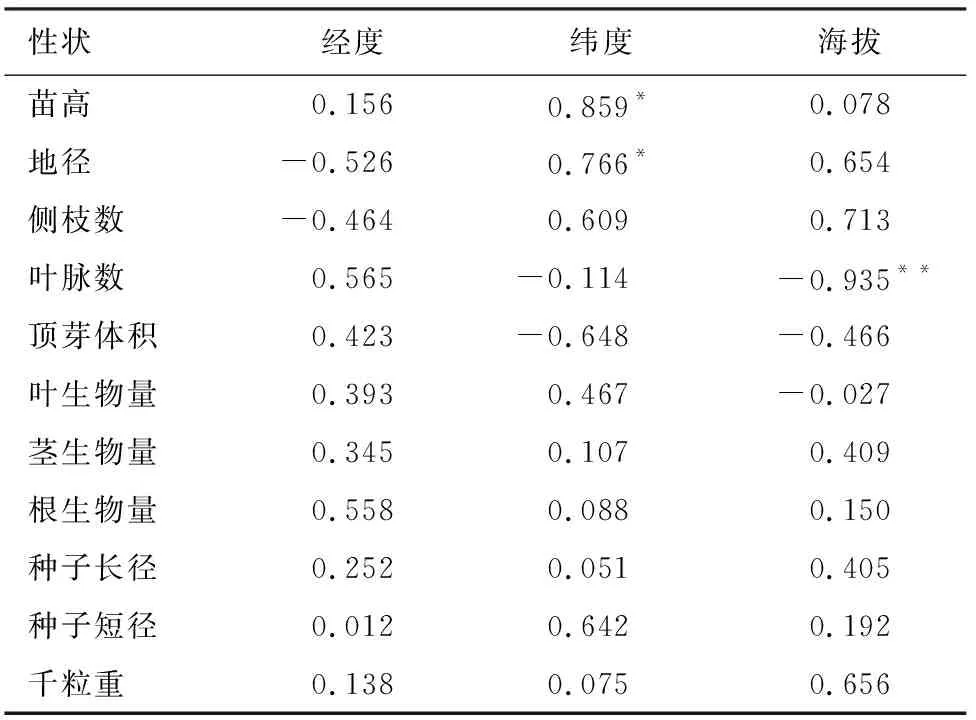
表5 不同种源蒙古栎种子及苗期表型性状与 地理因子相关性分析
2.6 综合分析
对不同种源蒙古栎各表型性状采用隶属函数综合评价法进行综合分析(表6)。JCZ种源地蒙古栎各项表型性状表现最优,其次为XFX,CHKZ表现最差。各种源按平均隶属度综合排序为:JCZ>XFX>WDZZ>CYX>DDS>PLBZ>CHKZ。

表6 不同种源蒙古栎综合评价
3 结论与讨论
通过对辽宁省7个种源的蒙古栎种子和苗期表型性状进行分析,在蒙古栎优良种源选择中,主要参考指标为苗高、地径、生物量、种子千粒重。通过多重比较分析得出,3种生长指标显著高于其他种源的种源地分别为:XFX、XFX和JCZ、XFX和JCZ、JCZ。
变异系数分析发现,在种子生长指标中变异系数最小的种源为XFX,苗期生长指标中变异系数最小的种源为JCZ。综上可见,XFX和JCZ种源地蒙古栎幼苗长势较好,种子平均千粒重较高,种源内部变异系数较小。因此,XFX和JCZ种源地入选为辽宁地区优良蒙古栎种源。
经过长期的地理隔离和自然选择,一些树种为适应当地的气候环境产生了相应的遗传变异,并能稳定遗传给下一代[15]。根据这一现象,许多研究通过分析同一树种在不同分布地区的生长差异,开展种源试验,选择出适应该树种生长的优良种源。为林木遗传改良提供重要途径和基础工作[16],是缩短育种周期、提高林木遗传改良效益的有效途径之一[17]。有关蒙古栎优良种源选择的其他试验中,屈红军等[18]对带岭试验地25个不同种源1年生及6年生蒙古栎的树高、地径进行多重比较分析,发现各生长性状存在着显著性差异,并选出苇河、集安种源可初步作为带岭试验地的优良种源应用于林业生产。有学者以辽宁12个蒙古栎种源为研究材料,对苗高和地径进行测量及方差分析,发现这2种性状差异均达到显著水平,最终筛选出3个地区作为优良种源[19]。通过变异系数分析发现,不同种源种子性状变异系数由小到大的变化趋势为千粒重、短径、长径、长宽比,比梁德洋等[20]对蒙古栎种子的研究结果偏低,且发现千粒重较长径和短径变异系数更小,稳定性更高。
通过地理因子相关性分析,发现一些地理因子对苗木某些生长指标产生了规律性的显著影响。本研究发现纬度高的地区蒙古栎幼苗长势好,本试验中的优良种源地XFX,纬度达到42.73835°N,为纬度最高的种源地。这种地理因子影响产生的变异规律在对花榈木(Ormosiahenryi)[21]的种源变异研究中也有体现,试验发现经度对花榈木苗高和茎生物量呈显著负相关,纬度对根尖数和茎生物量呈显著负相关,海拔对苗高和千粒重呈极其显著正相关,对茎生物量呈显著正相关。因此,在种源试验中应考虑种源与当地环境相互作用对每个种源个体生长表现的影响程度[22]。不同植物与地理因子相关性规律并不相同,同种植物不同种源其地理变异规律也不尽相同。
