珍稀濒危植物阜康阿魏的遗传多样性及遗传结构
2023-01-11曹秋梅李文军
杨 磊 曹秋梅 冯 缨 李文军*
(1. 荒漠与绿洲生态国家重点实验室,中国科学院新疆生态与地理研究所,乌鲁木齐 830011;2. 中国-塔吉克斯坦生物资源保育与利用联合实验室,中国科学院新疆生态与地理研究所,乌鲁木齐 830011;3. 资源与环境学院,中国科学院大学,北京 100049)
阜康阿魏(Ferula fukanensis)是伞形科(Apiaceae)多年生一次结果的草本植物,全株有强烈的葱蒜样臭味。该种为新疆特有种,珍稀濒危的国家二级保护野生植物[1-2],仅见于新疆阜康和乌鲁木齐有黏质土壤的冲沟和前山带,是典型的小种群野生植物。阜康阿魏分泌的具有特殊葱蒜样臭味的油胶树脂,具有重要药用价值,被作为药用阿魏的基原植物收载于《中华人民共和国药典》[3-4]。近年来,滥采盗挖、过度放牧和土地开发等因素对阜康阿魏的原生生境造成破坏,该种种群数量快速下降,濒临灭绝[5-6]。目前有关阜康阿魏的研究主要集中在资源调查[5-7],鲜见有关阜康阿魏遗传多样性的报道,这一定程度上限制了针对该种有效的保护管理措施的制定。
遗传多样性评价是对珍稀濒危物种自然种群管理的关键性步骤之一[8-10]。野生物种的遗传分析有助于掌握物种的遗传变异,预测种群栖息地丧失和破碎化[9,11]。理论上认为:广布物种比种群规模小、分布范围窄的稀有濒危物种的遗传多样性更大[12-16]。小种群由于近亲繁殖概率增加、种群结构不稳定和遗传漂变等因素加速了遗传多样性的丧失,进而导致其适应环境变化的能力和进化能力下降[8,13,16]。但最近的一些研究发现,种群规模较小的濒危物种有时也存在相对较高的遗传多样性[17-21]。阜康阿魏作为濒危的小种群野生植物,很有必要对其现有种群开展遗传多样性评价,以更好地对其开展保护。
本研究采用Li[20]等开发的阿魏属的微卫星引物(Simple sequence repeat,SSR),对采集于现存的3 个阜康阿魏居群的87 个个体进行了遗传多样性和遗传结构检测,以期厘清该种的遗传多样性和遗传结构,提出针对小种群阜康阿魏的保护和恢复建议,为有效保护和管理这种珍稀濒危的药用植物提供支撑。
1 材料与方法
1.1 试验材料
根据标本馆馆藏标本资料和野外实地调查,在现有分布居群阜康西泉(XQ,44°20′38.06″N,88° 25′2.66″E)、乌 鲁 木 齐 赤 焰 岭(CYL,43°49′16.01″N,87°25′38.41″E)和阜康小红沟(XHG,44°6′46.86″N,87°54′27.51″E)分别采集了30 株、22 株和35 株个体的新鲜健康叶片,在采样的过程中保证2 个个体间间隔不少于20 m,取每个个体20 g 以上新鲜叶片单独装袋,硅胶干燥后带回实验室备用。凭证标本保存在中国科学院新疆生态与地理研究所标本馆(XJBI)。
1.2 DNA提取、引物筛选及PCR扩增
每个样本取2 g 左右的干燥叶片,经粉碎机粉碎后,利用天根生化科技(北京)有限公司生产的新型植物基因组提取试剂盒(DP320)进行DNA提取,提取方法参照随试剂盒携带的操作指南。筛选出10 对高多态性引物对所有DNA 样本进行PCR 扩增,扩增体系为20 μL:模板DNA 1.0 μL 30 ng),上下游引物各0.5 μL(10 pmol·L-1),10×PCR Buffer 2 μL,dNTPs 0.5 μL(0.5 mmol·L-1),TaqDNA 聚合酶0.2 μL(0.5 U),DMSO 0.5 μL,H2O 14.8 μL。扩增反应程序:95 ℃预热5 min,之后分两个阶段,第一阶段10 个循环(95 ℃变性30 s,58 ℃复性30 s,72 ℃延伸30 s),第二阶段32 个循环(95 ℃变性30 s,55 ℃复性30 s,72 ℃延伸30 s),最后一次循环结束后72 ℃延伸5 min[20-21]。扩增产物利用QIAxcel全自动电泳系统进行毛细管电泳分型。电泳结果输出在QI Axcel Biocalculator 进行片段长度读取和预处理。PCR 产物在ABI 3730 自动测序仪上进行基因分型,使用混合分子大小标记(10 μL LIZ500:1 000 ulhi-di,Applied Biosystems,USA)。
1.3 数据处理
利用CERVUS v3.0.7[22]、POPGENE v1.32[23]和GenALEx 6.5[24]统计出每个位点水平和居群水平的等位基因数(Na)、有效等位基因数(Ne)、观测杂合度(Ho)、期望杂合度(He)、Shannon信息指数(I)、多态信息含量(PIC)、多态位点百分率(PPB)、近交系数或固定指数(F)、基因分化指数(Fst)。Nei’s基因多样性指数(h)通过FSTAT v1.2[25]计算。Hardy-Weinberg平衡通过1 000个随机分组进行评估,通过GENEPOP 网络版中的连续Bonferroni 校正调整P值[26-27]。采用GenAlEx 6.5[24]软件进行分子方差分析(AMOVA),并基于遗传距离矩阵对所有居群的遗传关系进行主成分分析(PCoA)。遗传结构采用Structure v2.3.4和Structure HARVESTER[28]软件进行分析,K分别设置为1~20,每个K值重复运算20 次。100 000 次的马尔科夫链蒙特卡罗(MCMC)重复之后进行30 000 次burn-in。使用CLUMPP v2.0[29]合 并 运 算 结 果,采 用Distruct v1.1[30]软件绘制居群遗传结构图。通过IBD v1.53对遗传距离与地理距离进行Mantel 相关性检验[31],采用BOTTLENECK v1.2.02[32]对阜康阿魏居群进行瓶颈效应检测分析。
2 结果与分析
2.1 遗传多样性分析
各位点以及3 个居群遗传多样性指数见表2~3。来自3个居群的87个个体共有39个等位基因,每个位点的等位基因(Na)从3 个(FS9、FS41、FS45)到6个(FS6)不等,平均等位基因为3.900,居群XQ、CYL 和XHG 的等位基因(Na)分别为3.100、2.900、3.700。平均有效等位基因(Ne)为2.205,各位点从1.115(FS2)到2.902(FS18)不等,XQ、CYL 和XHG 分别为2.250、2.118、2.247。平均观测杂合度(Ho)为0.881,高于平均期望杂合度(He)0.512,且各居群的观测杂合度也都高于期望杂合度(见表2~3)。3个居群多态位点百分率(PPB)均为100.00%。1个位点(FS2)多态信息含量比较低(PIC<0.2),5 个位点(FS1、FS9、FS21、FS41 和FS45)多态信息含量中等(0.2≤PIC<0.5),4 个位点(FS6、FS12、FS18 和FS36)多态信息含量比较高(PIC≥0.5)。除FS2外所有位点经Bonferroni 校正后显著偏离哈温平衡。居群间Nei’s 基因多样性指数(hS)和总的Nei’s 基因多样性指数(hT)分别为0.514 和0.516。3 个居群的近交系数均为负值,平均值为-0.693。
2.2 遗传分化和遗传结构分析
3 个居群的居群内分化系数(Fst)为0.007(见表2),平均基因流非常高(Nm=5.874)。AMOVA 分析显示,群体间变异仅占总遗传变异的4.1%,而群体内变异占95.9%。居群间基因分化系数(PhiPT)为0.041(见表4)。贝叶斯聚类分析结果如图1 所示,选择K=2作为最优K值,因为K=2时ΔK的值最大。Mantel 相关性检验也表明,不同种群间的地理距离和遗传距离(R2=0.136,P=0.669)无显著相关关系(P>0.05)。PCoA 分析的第2、第3主成分分别解释累计总变量的30.43%和43.37%(见图2)。
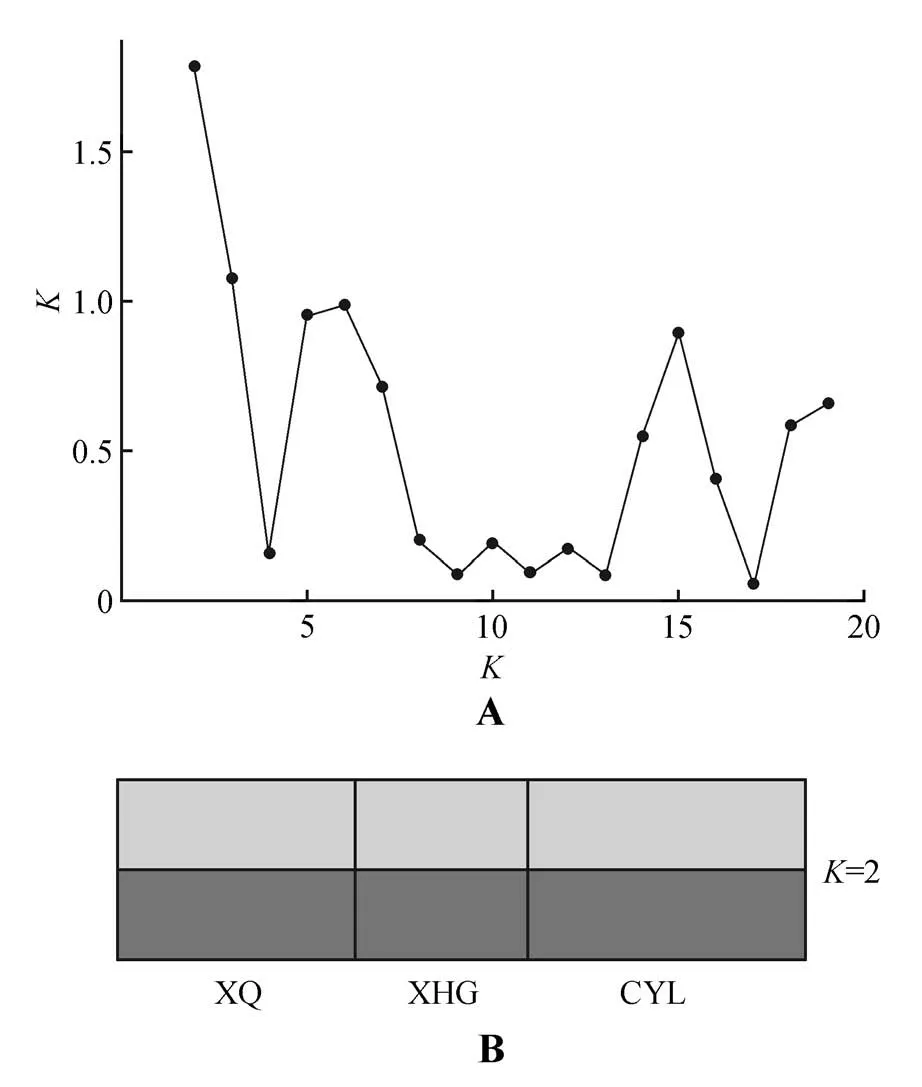
图1 阜康阿魏遗传结构分析A.基于20 次重复的Delta K 的对数似然函数值;B.根据10 个SSR位点变异推断3个居群的遗传结构(任何一个K值显示的3个种群都是完全相同,此处仅展示K=2)Fig.1 Genetic structure analysis of F.fukanensis A.Log-likelihood function values of Delta K based on 20 replicates;B.The genetic structure of based on the variation of 10 SSR loc(iAny value of K showed that the three populations were exactly the same,and only K=2 was shown here)
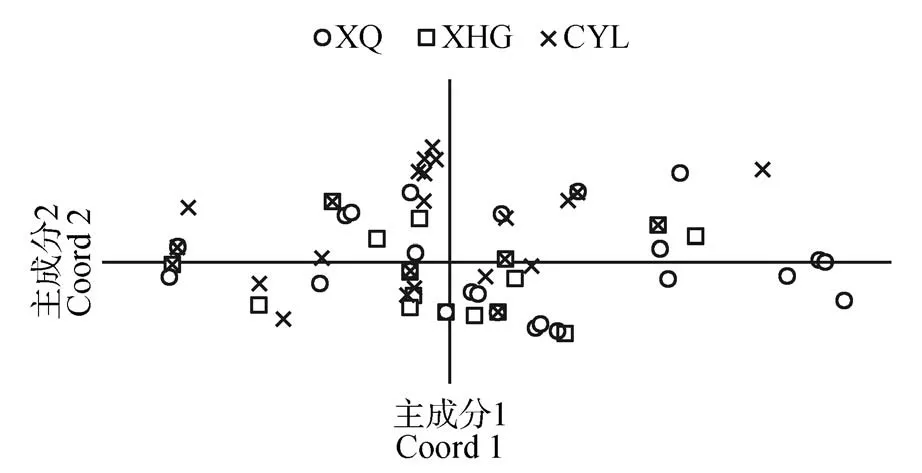
图2 阜康阿魏遗传距离的主成分聚类分析Fig.2 PCoA clustering analysis of paired genetic distance of F.fukanensis
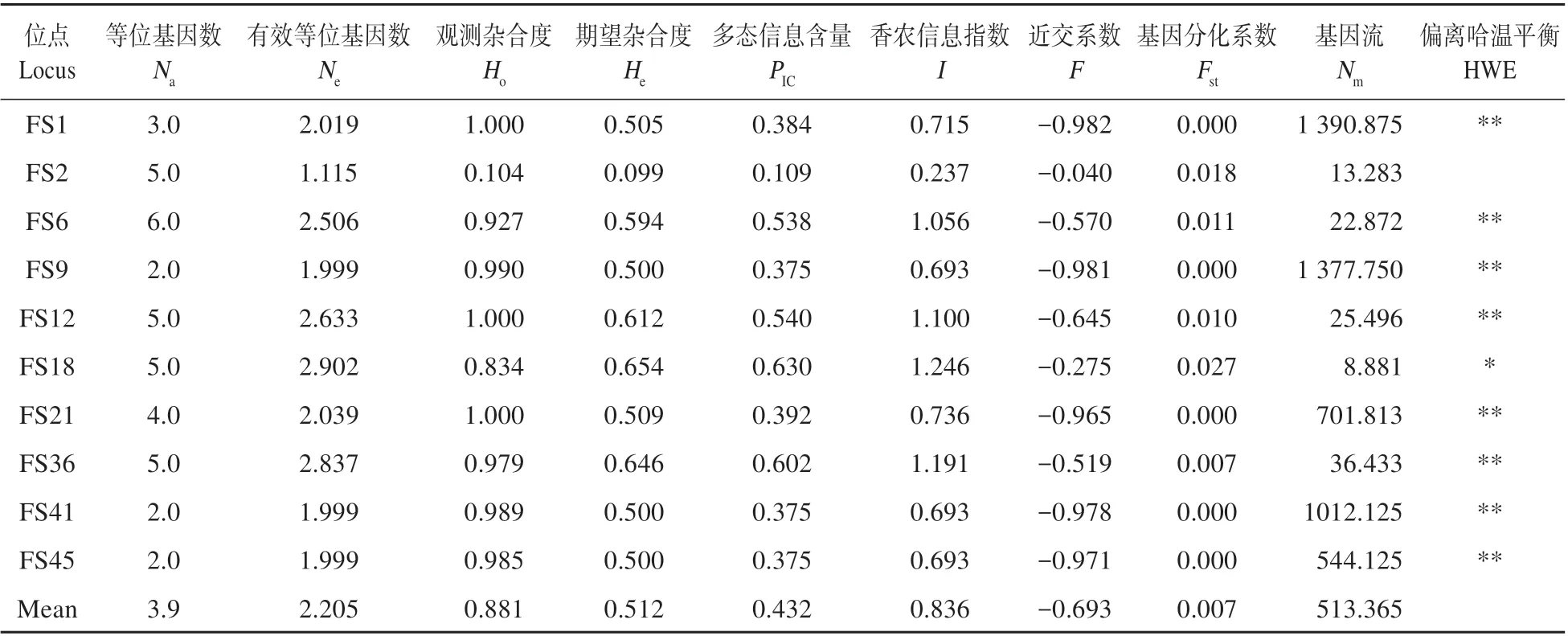
表2 各位点遗传多样性指数Table 2 Index of genetic diversity for all loci
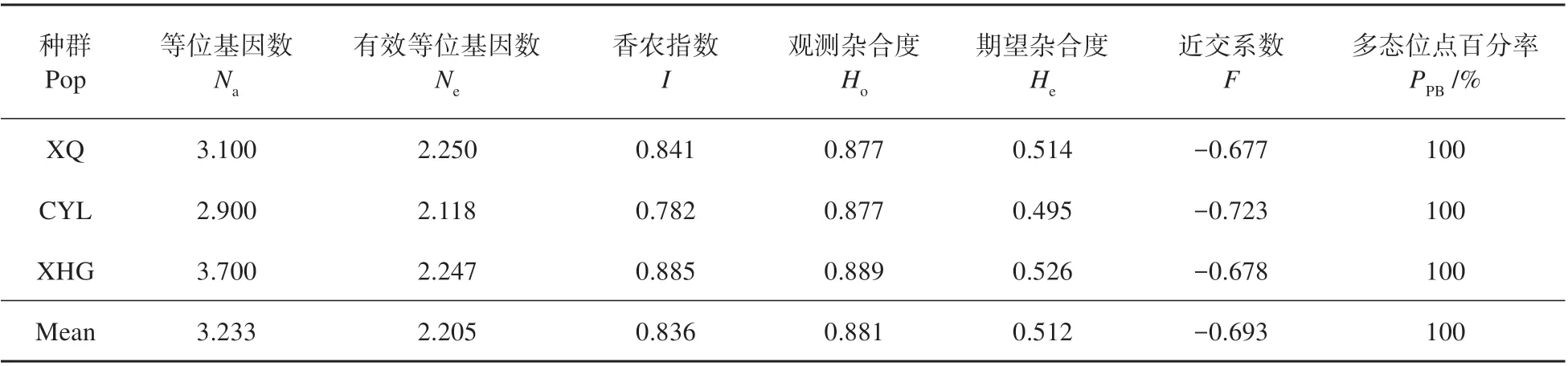
表3 各居群遗传多样性指数Table 3 Index of genetic diversity of F.fukanensis populations
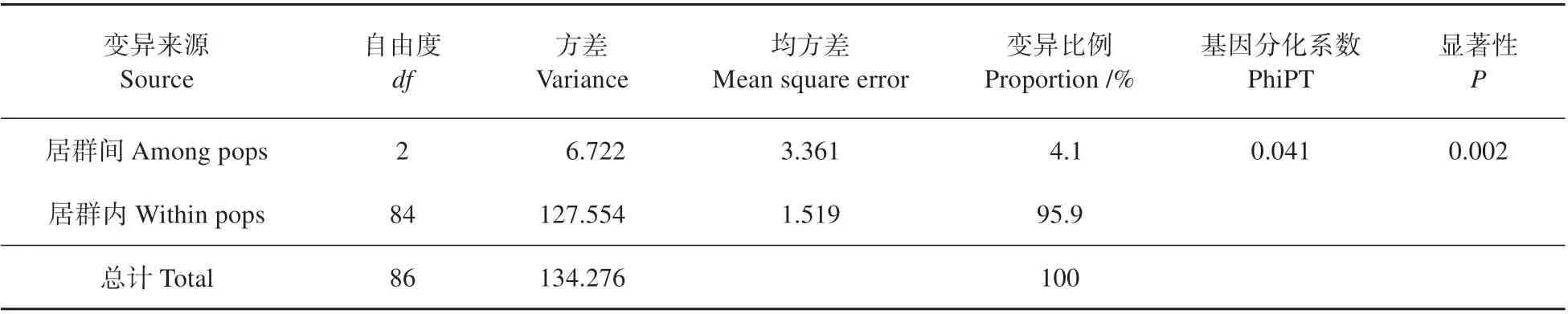
表4 居群间和居群内的分子方差分析Table 4 Results of AMOVA between and within populations of F.fukanensis
2.3 瓶颈效应检测
3 个居群在无限等位基因模型(IAM)和双相突变模型(TPM)均显示存在显著的杂合度过剩(P<0.05),在逐步突变模型(SMM)下不显著(P>0.05),只有XHG居群呈L形分布(见表5)。
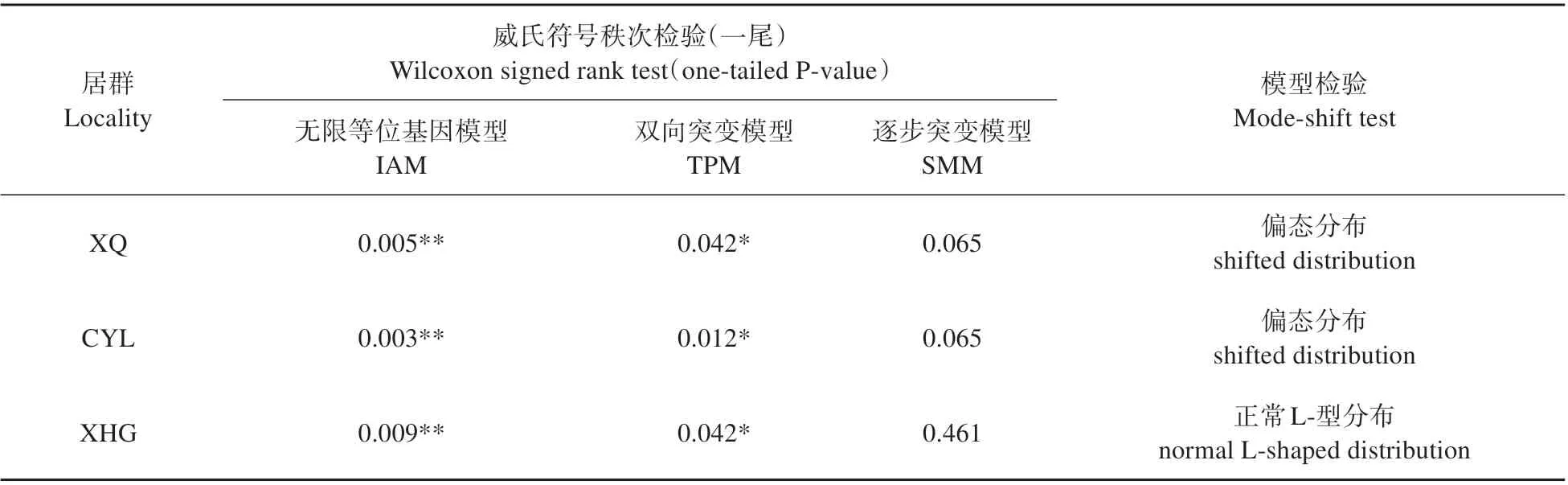
表5 阜康阿魏瓶颈效应检测Table 5 Detection of bottleneck effect for F.fukanensis
3 讨论
3.1 阜康阿魏遗传多样性
本研究通过前期开发的10对SSR引物对阜康阿魏进行遗传多样性评价,结果发现阜康阿魏表现出较高的遗传多样性(hS=0.514,hT=0.516,Ho=0.881,He=0.512,I=0.836,PPB=100%)。低于广布物种的平均值(He=0.58),却高于其他濒危物种的平均值(He=0.42)[16]。相较阿魏属其他物种的遗传多样 性 也 要 高 很 多,如F. loscosii(He=0.125,hT=0.152)[33],F. communis(Hpop=0.320)[34],F. communiscomplex(Hw=0.263,hT=0.317)[35],F. arrigonii(Hw=0.317,hT=0.336)[36]和F. asafoetida(hT=0.34,I=0.51)[37]。与其他濒危物种相比,如Nuphar submersa(He=0.42)[38],Ottelia acuminatavar.jingxiensis(He=0.441,I=0.781)[39],Ruta oreojasme(He=0.687)[40],Vincetoxicum atratum(He=0.67)[41],Ammi seubertianum(He=0.66,I=1.28)和A. trifoliatum(He=0.67,I=1.35)[42],Tapiscia sinensis(He=0.690 4,I=1.436 8)[43],也具有中等的遗传多样性。理论上,由于近交衰退和基因漂变的原因,濒危物种与广布物种相比有较低的遗传多样性[44-45],而阜康阿魏相对较高水平遗传多样性的结果明显不符合小种群、狭窄分布物种表现出低水平遗传多样性的普遍假设[14,46-48]。
影响珍稀濒危植物遗传多样性的因素主要包括物种自身因素(繁育系统、瓶颈效应、生活史等)和人为因素(自然资源过度开发、栖息地退化、土地利用等)[12,17,47,49-51]。理论上认为异交物种通常比自交物种具有更高的遗传多样性水平[15,52]。关于阿魏属植物繁育系统和传粉生物学的相关研究表明,异交可能是阿魏属植物的主要交配方式[53-54]。阿魏属植物异交为主的繁育方式可能是保留较高水平遗传多样性的主要因素。本研究中涉及的3 个居群具有非常高的平均基因流(Nm=5.874)也一定程度上验证了居群间存在广泛的交流。阜康阿魏较高水平的遗传多样性很大程度是由于短时间内对阜康阿魏过度破坏(滥采盗挖、过度放牧等因素)导致种群数量减少的速率远高于其遗传多样性降低的速率,现存的阜康阿魏继承了原来大种群的部分遗传变异。但现有小种群可能会增加阜康阿魏近亲繁殖和遗传漂变的概率,并增加衰退风险[55]。
3.2 阜康阿魏遗传结构和遗传分化
本研究发现3 个居群间的遗传变异和分化程度较低(Fst=0.007),PCoA、AMOVA 和Structure 分析也支持了这一结论。AMOVA 分析显示阜康阿魏95.9%的遗传变异发生在居群内,仅4.1%的变异发生在居群间,可能是由于巨大的基因流(Nm=5.874)抵消了遗传漂变的影响,减少了居群间的遗传变异[20-21]。Structure 结果显示,当K=2 时,ΔK值最大,3 个居群划分了2 个基因库,随着K值的增加,仍有多个拐点出现,说明出现了新的基因库,表明阜康阿魏杂合性很高,遗传背景复杂[56]。Structure 未能将居群分组,Mantel 相关性检验也显示了不同种群间的地理距离和遗传距离(R2=0.136,P=0.669)无显著相关关系(P>0.05)。基因瓶颈效应检测显示,XHG 居群为正常L 分布,还未经历遗传瓶颈,而XQ 和CYL 居群,都为shifted 分布,说明它们最近经历了遗传瓶颈,该结果可能也是由于对阜康阿魏种群的过度破坏造成的。
3.3 保育策略
研究结果表明阜康阿魏具有相对可观的遗传多样性和进化潜力,遗传因素并不是该种濒危的主要原因。通过野外调查推测,人为滥采和土地开发使得阜康阿魏物种栖息地破碎、缩减,种群数量急剧下降,可能是阜康阿魏当前面临的最大挑战。考虑到小种群会增加近亲繁殖和遗传漂变的风险,建议开展就地保护和迁地保护工作等加强对阜康阿魏的保护:(1)建立阜康阿魏保护区,保护阜康阿魏的原生境;(2)加强监管力度,防范原生境农田开垦和盗采盗挖等人为活动破坏种群更新;(3)采集种子或幼苗进行种质资源的保存和迁地保育;(4)加强种群动态监测,并开展相关基础研究工作,辅助阜康阿魏种群复壮;(5)加强宣传教育,提高公众的保护意识,形成全民参与的社区保护模式。
