巨噬细胞源性外泌体调控骨改建的研究进展
2023-01-10刘艺刘奕
刘艺 刘奕
中国医科大学附属口腔医院正畸科 沈阳 110013
巨噬细胞是人体固有免疫的主要效应细胞,参与伤口愈合和组织再生等多种生理病理过程[1-2]。外泌体是细胞在生理、病理条件下分泌的一种直径50~150 nm的微小囊泡,因其从细胞中主动释放的特定机制、参与细胞间相互作用及作为疾病标志物而成为热点[3]。骨改建过程受到多种信号通路调节,现普遍认为炎症细胞和骨细胞(成骨细胞、破骨细胞和骨细胞)之间的相互作用是骨改建的关键[4]。巨噬细胞与骨组织关系密切,外泌体也在骨骼疾病中起着至关重要的作用[5-6]。大量学者对不同状态下巨噬细胞来源的外泌体功能进行了研究。阐明巨噬细胞外泌体与骨骼改建相互作用的机制有助于控制炎症反应及干预疾病,开发新的治疗策略。本文就巨噬细胞源性外泌体作用于骨改建的研究进展做一综述。
1 巨噬细胞与骨改建
巨噬细胞起源于骨髓和卵黄囊[7-8],是一类高度异质性的细胞。不同的刺激信号可使巨噬细胞展现出不同的表型,即巨噬细胞的极化。根据表面标记物、细胞因子和功能活性不同,极化巨噬细胞大致可分为2种亚型:经典激活或炎症型巨噬细胞(M1) 和交替激活或抗炎症型巨噬细胞(M2)[8]。与之相对应的未激活巨噬细胞通常标记为M0。M1型巨噬细胞通常由干扰素-γ、细菌脂多糖识别诱导[9],产生促炎为主的细胞因子如肿瘤坏死因子(tumor necrosis factor,TNF)-α、白细胞介素(interleukin,IL)-1β,具有较强的抗微生物活性[10]。M2型巨噬细胞由IL-4、IL-10等诱导生成,产生IL-10、TGF-β等,主要吞噬碎片、抑制炎症反应。M2巨噬细胞可进一步区分为4个亚群,即M2a、M2b、M2c、M2d[8,11]。M1、M2只是巨噬细胞极化的两个“极端”,主导的表型改变时,其炎性细胞因子的分泌和基因表达也会发生变化。活化的巨噬细胞通过细胞间直接接触和/或释放分泌体(细胞因子、细胞外泌体)与各种类型的靶细胞进行通讯,以发挥其免疫调节作用。
骨改建包括了初期清除受损或老化的骨组织和后期骨形成的动态过程[12]。酶、激素、细胞因子、趋化因子等信号都可调节这一过程。巨噬细胞与骨组织细胞起源相近,共同与骨组织浸润在骨髓微环境中,与骨组织关系非常密切。用药物耗竭小鼠体内的巨噬细胞,截骨处骨痂及基质生成均降低[13]。作为炎症的调节细胞,巨噬细胞参与骨愈合的全过程,并在不同阶段表现出不同的极化类型,是启动愈合级联和正确引导骨痂形成的关键。许多学者[6,14-16]发现骨损伤后早期M1巨噬细胞及破骨细胞浸润,晚期转变为M2细胞为主。但长期的促炎反应(M1表型)对骨愈合有负面影响[17]。
2 巨噬细胞源性外泌体
细胞外囊泡(extracellular vesicles,EVs)是指细胞分泌的一种双层脂质结构小泡,是外泌体、微囊泡和凋亡小体等结构的通称[18]。大部分文献采用“外泌体”(exosomes) 一词指代囊泡结构[19]。外泌体携带其亲本细胞的蛋白质、代谢物、核酸和脂质等内容物,具有与其亲本细胞相似的作用[20-21]。它能够通过表面标志直接与受体细胞结合,还可以通过旁分泌及膜融合(内化)的方式进入胞质[22],将内含物从来源细胞转移到受体细胞,进而调节受体细胞的功能。越来越多的学者提出巨噬细胞外泌体在调控炎症或骨病进展中起着关键作用[5]。研究[23-25]证实,极化的巨噬细胞外泌体内容物及含量并不相同。阐明这些外泌体调控骨微环境的机制有利于针对骨炎症性疾病的治疗或干预骨改建的进程,达到动态平衡。
3 巨噬细胞源性外泌体调控骨改建
3.1 M1巨噬细胞外泌体对骨改建的作用
一般认为,M1极化的巨噬细胞促进趋化因子和炎症介质的释放,有助于初始破骨细胞激活、骨折部位碎片吸收[26-28]。MiRNAs是一种高度保守的非编码小RNA分子,通过调节基因表达的转录后控制来实现其生物学功能,是外泌体的重要内容物之一[29]。M1及M2极化的巨噬细胞富集的mi-RNA有所不同。Kang等[24]发现,M1外泌体富含miR-155,它能够负调控骨形态发生蛋白(bone morphogenetic protein,BMP)信号级联,特异性降低BMP2、BMP9和Runt相关转录因子(Runt-related transcription factor, Runx) 2 的 表 达 , 而BMP2是成骨细胞分化必不可少的因子[12],从而抑制骨再生。Ge等[25]则发现M1外泌体的miR-155能够靶向抑制下游细胞因子信号转导抑制因子6(SOCS6),提高p65 蛋白含量从而激活核因子(nuclear factor,NF)-κB信号通路,加重炎症反应。miR-222同样在M1型巨噬细胞中高表达[30]。研究[31]发现,M1外泌体富集的miR-222可以通过抑制Bcl-2蛋白诱导骨髓间充质干细胞(bone marrow mesenchymal stem cells,BMMSCs) 凋亡。Yu等[32]报告M1外泌体miR-98靶向MC3T3-E1细胞中的DUSP1从而抑制成骨分化。研究[33]表明,M1-外泌体富含miR-1246,通过降低GSK3β和Axin2的表达激活Wnt途径,导致软骨炎症和降解。这些miRNA大多结合生物信息学、测序等方式证实。由此可见,M1 巨噬细胞外泌体富集miR-155、miR-222、miR-98、miR-1246等,能够作用于下游信号通路从而促进炎症、抑制骨修复。miRNA的含量影响其对下游信号的作用,并靶向信号传导网络中关键节点,而外泌体可以作为特定的载体,这使得巨噬细胞外泌体miRNA成为一类有前景的可调控骨改建的生物标志物。然而,目前对巨噬细胞外泌体的内容物了解仍不够透彻,关于极化对miRNA含量改变的影响以及miRNA与下游通路的作用机制仍需要进一步的探索。
NF-κB是5种转录因子的统称,可在多种因素刺激下激活,是典型的促炎型号通路[34]。NF-κB通路激活后能促进IL-1β和TNF-α等细胞因子的表达,而它们既是促进巨噬细胞M1极化的分子,也是NF-κB通路的反馈激活因子[26,35],进一步导致促炎环境。McDonald等[3]在小鼠炎症疼痛模型中证明脂多糖刺激的巨噬细胞(即M1巨噬细胞)外泌体中检测到丰富的细胞因子并导致NF-κB的转录激活,而未进行刺激的巨噬细胞(即M0巨噬细胞)不能激活NF-κB通路。Wang等[36]进一步证实脂多糖诱导的巨噬细胞外泌体激活NF-κB通路,促进细胞因子的释放,建立局部炎症环境。炎性细胞因子在调节骨骼破坏和成骨细胞生成方面起着重要作用。组织损伤后,损伤组织和凋亡细胞大量存在,必须清除感染后才能启动改建过程。巨噬细胞转化到促愈合表型,释放细胞因子,趋化因子和其他促间充质干细胞、成骨细胞系细胞归巢的因子促进组织修复和重新建立组织稳态。这些特征表明M1外泌体通过作用于NF-κB治疗炎症相关骨疾病和通过促进炎症而调节骨转换具有巨大潜力。此外,miRNA虽然不是直接的NF-κB靶点,但也可以调节NF-κB信号[29]。
许多学者对巨噬细胞外泌体如何影响BMMSCs的分化能力进行了研究。He等[37]提取M1巨噬细胞产生的条件培养基(conditioned medium,CM)进行研究,发现其促进BMMSCs增殖、成脂分化及细胞外基质的沉积。进一步提取CM中的外泌体,发现促进干细胞增殖、成骨、成脂分化的是M1外泌体而不是M2外泌体,Xia等[38]得出同样结论。但Kang等[24]将M1极化的巨噬细胞与小鼠间充质干细胞(mesenchymal stem cells , MSCs)共培养后,MSCs的BMP2和BMP9的表达显著降低,表明M1巨噬细胞外泌体损害了BMP-2的骨诱导作用以及MSCs的成骨诱导基因的表达。M1巨噬细胞主要通过外泌体促进早期和中期成骨,并对BMMSCs的增殖、成骨和成脂分化具有更强的作用。其对间充质细胞成骨的负性调节很可能与受体细胞来源不同有关。由于巨噬外泌体是从特殊的CM(无外泌体血清培养基与巨噬细胞共培养一段时间)中提取的,其他原因如巨噬细胞的成熟度、共培养条件、CM中其他细胞因子的影响、细胞谱系甚至技术差异都可能影响实验结果[27,38]。BMMSCs的成骨和成脂平衡是维持骨量的关键。探索调节BMSSCs成骨和成脂平衡的机制可能有助于临床疾病机制的探索及靶向药物应用。M1外泌体对骨改建信号通路的调节如图1所示。
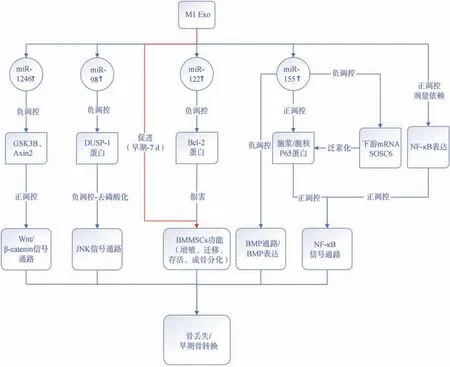
图1 M1巨噬细胞外泌体对骨改建信号通路的调节Fig 1 The regulation of M1 macrophages-derived exosomes on the signal pathway of bone remodeling
3.2 M2巨噬细胞外泌体对骨改建的作用
骨愈合的一个关键因素是从最初的促炎M1型巨噬细胞转变为抗炎M2型[39]。M2型细胞在骨愈合后期建立抗炎环境,可促进BMMSCs成骨。Li等[5]认为,外泌体是M2巨噬细胞诱导成骨过程中的重要介质。
BMPs是内源性调控成骨的重要信号通路。BMP2和BMP4的丢失会导致严重的成骨障碍,BMP还可通过激活Smad蛋白并上调Runx2的基因表达,而Runx2是诱导间充质干细胞向成骨细胞分化的主要转录因子[40-41]。Kang等[24]通过体内骨缺损实验证实M2外泌体能够促进骨再生,进一步分析得知M2外泌体富含miR-378a,通过调节BMP信号通路促进骨诱导,导致小鼠颅骨骨再生增强。研究[42]发现,IL-4刺激的巨噬细胞(M2)外泌体富集的miR-99a-5p、miR-146b-5p或miR-378-3p都通过下调TNF-α和NF-κB信号通路来抑制炎症。M2巨噬细胞外泌体通过miR-690、IRS-1和TAZ促进BMMSCs的成骨分化,但抑制成脂分化[5]。Yu等[43]则发现,M2外泌体富集的miR-690还可通过抑制NF-κB p65蛋白的翻译,上调成骨分化。此外,有报道称M2巨噬细胞来源的外泌体高表达miR-221-5p,通过与E2F2的30UTR结合负调控其表达,促进组织修复再生[44]。这些研究都表明,M2巨噬细胞外泌体浸润在骨髓微环境中,并将miRNAs稳定地转移到BMMSCs,有助于成骨分化和骨折愈合[45-46]。这些miRNA(miR-378a、miR-690、miR-99a-5p等)可能是成骨的靶点,并可调节BMP等通路促进成骨。然而,M2外泌体miRNA在骨形成和破骨细胞形成中的作用仍然不清楚[47]。
越来越多的证据表明,BMMSCs和M2巨噬细胞外信号的相互作用参与了骨的形成。但由于细胞来源不同、共培养条件不同以及巨噬细胞的状态不同,这种相互作用的机制和细节仍然存在争议。有学者[48]提出,M0或M2巨噬细胞产生的CM在成骨的后期阶段有利于BMMSCs的矿化。但是也有学者得出了相反结论。Xia等[38]的研究得出,M2巨噬细胞外泌体损害了BMMSCs的增殖。M2外泌体对BMMSCs的成骨、成脂相关基因的表达没有明显影响,甚至对软骨分化有抑制作用。M2外泌体对骨改建信号通路的调节如图2所示。
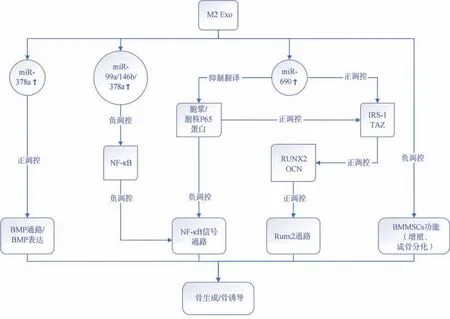
图2 M2外泌体对骨改建信号通路的调节Fig 2 The regulation of M2 macrophages-derived exosomes on the signal pathway of bone remodeling
4 结语
巨噬细胞是先天免疫系统的关键介质,巨噬细胞源性外泌体可发挥促炎、促进早期成骨或抗炎、促进骨修复的作用。M1和M2亚型巨噬细胞外泌体对骨改建的作用不同或相反,这大部分取决于其各自高表达的miRNA与下游信号通路的作用,但与它们的来源细胞功能一致。二者对NF-κB通路的调节尤其相反,M1巨噬细胞外泌体富集的miR-155能够直接靶向下游SOSC6,减少其对细胞p65蛋白的降解和泛素化,从而提高p65含量,促进NF-κB通路和介导炎症。而M2巨噬细胞外泌体通过其富集的miR-690直接抑制p65蛋白的翻译过程,减少其含量而抑制NF-κB通路。
良好控制的炎症反应可能有益于骨修复,但当炎症反应过度,可能会对骨组织造成破坏[49-50]。巨噬细胞外泌体为纳米级结构的流体(悬液),在无细胞治疗的发展中有着广阔的前景,由于它可将蛋白质、miRNA等转运到受体细胞,针对极化巨噬细胞的外泌体治疗可能成为炎症性骨病或调节炎症反应的治疗方式。外泌体与微环境之间相互作用的机制、外泌体与生物材料的整合都是值得探讨的问题。今后的工作中,有必要将巨噬细胞外泌体应用于动物,进一步验证对骨改建的效果、机制和治疗剂量。
利益冲突声明:作者声明本文无利益冲突。
