伊犁马PFKM基因表达及酶活性分析
2023-01-10曾亚琦王建文吐尔逊江吾木尔艾力王川坤袁鑫鑫王彤亮姚新奎
王 洁,孟 军,2,曾亚琦,2,王建文,2,吐尔逊江·吾木尔艾力,王川坤,袁鑫鑫,王彤亮,姚新奎,2
(1.新疆农业大学动物科学学院/新疆农业大学马产业研究院,乌鲁木齐 830052;2.新疆马繁育与运动生理重点实验室,乌鲁木齐 830052;3.新疆伊犁哈萨克自治州畜牧总站,新疆伊宁 835000)
0 引 言
【研究意义】伊犁马环境适应性和抗病力极强,耐粗饲、耐寒,拥有力量与速度并存点,探究伊犁马糖酵解具有深刻的研究意义[1-3]。在葡萄糖分子转化利用的过程中,PFKM基因对磷酸果糖激酶的有重要影响,而磷酸果糖激酶是糖酵解过程的起点[4]。糖酵解途径中有3个重要的限速酶,即己糖激酶(HK,hexokinases)、磷酸果糖激酶-1(PFK1,phosphofructokinase1)以及丙酮酸激酶(PK,pyruvate kinase)。在其反应中,磷酸果糖激酶(PFK)是主要的调节酶,是糖酵解途径的起博器[5-6]。其中6-磷酸果糖激酶(6-PFK)是糖酵解过程中将ATP分解的关键酶[7-8]。而PFK1有3种同工酶:PFKM为肌肉型、PFKP为血小板型以及PFKL肝脏型,以四聚体或二聚体的形式参与不同组织部位的糖酵解。【前人研究进展】磷酸果糖激酶(PFKM)被证实可调节葡萄糖代谢及肌肉维持等多种生物过程,是糖酵解的一个关键限速酶,也是葡萄糖代谢中起决定作用的限速酶。这种酶催化Mg+、ATP依赖的果糖-6-磷酸磷酸化,并形成ADP和果糖-1,6-二磷酸以及刺激6-磷酸果糖激酶的活性来控制机体的糖酵解代谢[9]。研究证实PFKM基因的相对表达量直接影响糖酵解的反应速率,在家畜家禽和猪中已证明PFKM基因表达变化与肉质有直接的关联性[10-12]。【本研究切入点】目前,在国内外未见关于伊犁马组织内PFKM基因表达及酶活方面的有关研究。需研究测定PFKM基因在伊犁马不同组织部位中的差异表达。【拟解决的关键问题】以伊犁马为试验动物,利用实时荧光定量PCR技术结合酶联免疫吸附(Elisa)检测手段,分析心脏、肝脏、夹肌、斜方肌、背阔肌、臀中肌、半腱肌以及腹外斜肌8种组织中PFKM基因mRNA的表达水平差异及不同组织中6-PFK的酶活规律,研究糖酵解途径中PFKM基因以及相关酶在伊犁马生命活动中的变化规律,为其糖酵解过程中提供理论基础。
1 材料与方法
1.1 材 料
1.1.1 试验动物
挑选年龄、体况相近的8匹伊犁马作为试验对象,屠宰后1 h内分别采集心脏、肝脏、夹肌、斜方肌、背阔肌、臀中肌、半腱肌以及腹外斜肌8种组织样品,用自封袋包装置于液氮罐-196℃保存。
1.1.2 主要试剂
Elisa试剂盒(6-磷酸果糖激酶);C2H5OH(分析醇)国药集团化学试剂有限公司、Trizol、氯仿北京化工厂;6×Loading Buffer北京索莱宝科技有限公司;RNase-free H2O北京天根生化科技有限公司;HyperScript III RT SuperMix for qPCR with gDNA Remover、2x S6 Universal SYBR qPCR Mix等均购自新贝(上海)生物科技有限公司。
1.2 方 法
1.2.1 引物设计与合成
根据NCBI数据库已发表的PFKM基因序列,利用prime 5软件设计PFKM荧光定量PCR引物,其中GAPDH为其内参基因。引物由新疆康普森有限公司合成。引物使用去离子水稀释,存于-20℃冰箱内保存。表1

表1 引物信息
1.2.2 RNA提取和cDNA合成
运用Trizol法得到伊犁马不同组织部位中RNA,采用1%的琼脂糖凝胶电泳验证RNA的完整程度。根据反转录试剂盒(HyperScript III RT SuperMix for qPCR with gDNA Remover)的操作步骤反转录合成cDNA;反转录反应体系:RNA Template100ng-2 μg,5×RT SuperMix 4 mL,RNase-free ddH2O补充至20 mL ,DTT(0.1mol /L)2 mL;反应条件:37℃ 15 min,85℃ 5s;接下来进行离心,利用超微量紫外分光光度计检测cDNA浓度,然后用无酶水稀5倍,-20℃保存于冰箱备用。
1.2.3 目的基因实时荧光定量检测
利用实时荧光定量PCR技术测试不同部位样本中PFKM基因mRNA表达量;反应体系:S6 UniversaL SYBR qPCR Mix 5mL,Primer-F(10 mM)0.2 mL,Primer-R(10 mM)0.2 mL,将cDNA模板(500 ng/mL)1 mL,以及RNase-free ddH2O添加至10 mL;反应条件:在95℃时预变性30s,95℃时变性10s,60℃时退火30s,72℃延伸至45s,总共40个循环。
1.3 数据处理
荧光定量PCR的数据用2-△△Ct法检测PFKM基因在伊犁马不同部位中差异表达量。第一步计算伊犁马不同组织部位目的基因和内参基因的Ct差值即目的基因与内参基因的差值,接下来利用Excel计算不同部位的△Ct平均值,肝脏部位为对照组,其他部位△Ct减去肝脏的△Ct,即△△Ct也为目的基因减去内参基因的差值,再用试验组-△△Ct(目的基因与内参基因的差值)对照组[13];PFKM基因mRNA相对表达量数据用SPSS Statistics26.0软件ANOVA方差分析,并且在Excel中对数据进行作图。
2 结果与分析
2.1 伊犁马不同组织部位提取RNA完整性检测条带
研究表明,伊犁马不同组织部位中提取的RNA利用1%琼脂糖凝胶电泳检测,条带十分清晰且整齐,亮度明显,无特别明显的扩散现象,结果质量好。图1
2.2 实时荧光定量RT-PCR产物的特异性
研究表明,扩增效率较高,斜率较高,分别为95.5%和96.4%,因其斜率较高,拐点鲜明,均呈“S”型,线条平缓,两者溶解曲线显示引物特异性和重复性较好,无引物二聚体出现及非特异性扩增产物产生,扩增曲线为人类明显的单峰,无明显扩增杂峰,所设计引物特异性良好,符合试验的质量要求。图2、图3
注:1:夹肌;2:斜方肌;3:背阔肌;4:臀中肌;5:半腱肌;6:肝脏;7:心脏;8:腹外斜肌

图2 PFKM基因qRT-PCR 溶解曲线和扩增曲线

图3 GAPDH基因qRT-PCR溶解曲线和扩增曲线
2.3 PFKM基因mRNA在不同组织的相对表达
研究表明,伊犁马PFKM基因mRNA在心脏组织中的相对表达量最高为22.88,极显著高于其他组织(P<0.01);夹肌组织、斜方肌组织、背阔肌组织、臀中肌组织、半腱肌组织及腹外斜肌组织中PFKM的表达量极显著高于肝脏组织(P<0.01);心脏组织的表达量最高,其次是半腱肌、背阔肌、腹外斜肌、臀中肌、夹肌、斜方肌;在肝脏组织中最低。图4
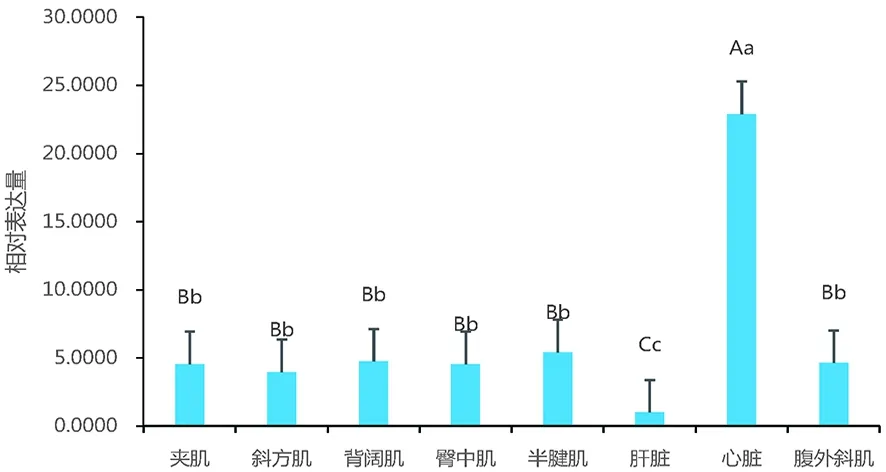
图4 伊犁马PFKM基因mRNA在不同组织中的相对表达量
2.4 6-磷酸果糖激酶在不同组织的含量
研究表明,伊犁马不同组织中6-PFK在心脏组织中含量为1.049 mg/L极显著高于背阔肌、肝脏、臀中肌、半腱肌、腹外斜肌、斜方肌、夹肌(P<0.01);而背阔肌中的含量也极显著高于肝脏、臀中肌、腹外斜肌、半腱肌、斜方肌、夹肌(P<0.01);臀中肌中的含量极显著高于腹外斜肌(P<0.01);其中臀中肌中的6-PFK显著高于夹肌、斜方肌和肝脏(P<0.05);半腱肌中的含量显著高于腹外斜肌(P<0.05);其他各组织之间含量无差异。表2

表2 6-PFK在不同组织的含量
3 讨 论
在糖酵解过程中,肌肉型磷酸果糖激酶(PFKM,phosphofructokinase-muscle)已被确认是其中之一的主要调控酶,将6-磷酸果糖激酶(6-PFK)转化为1,6-双磷酸果糖,此过程是不可逆的。6-磷酸果糖激酶(6-PFK)主要通过PFKM基因影响糖酵解的速率,并且PFKM基因的相对表达量直接影响糖酵解的反应速率[10]。研究结果显示,在伊犁马的不同组织部位中,PFKM基因在心脏组织中高度表达,且6-磷酸果糖激酶的活性也最高,在伊犁马的糖酵解代谢过程中,6-磷酸果糖激酶(6-PFK)的含量可能是PFKM基因的标志代谢物。PFKM基因在伊犁马的糖酵解过程中造成的生理及生化的具体规律尚未明晰。对PFKM基因利用荧光定量qRT-PCR检测,开展相应的研究,证实PFKM基因对6-磷酸果糖具有相对较高的亲和力[14]。试验研究与以上相关实验结果一致,通过荧光定量qRT-PCR对伊犁马的PFKM基因的表达与6-磷酸果糖激酶(6-PFK)进行检测,发现PFKM基因和6-PFK的含量极显著高于其他组织。在生命活动中,PFKM基因的异常表达,也造成了葡萄糖的代谢过程的改变,尤其是增加了组织细胞的糖酵解速率,在一定水平上可以利用降低PFKM基因的表达及相关的酶活性的方法可以影响不同组织的发育水平,进而降低细胞的生长速度[15-17]。因此在PFKM基因和6-PFK在心脏组织中高度表达,其心脏在生命体能量代谢以及糖酵解途径中,通过酶活性调节代谢的方式为机体提供能量,而磷酸果糖激酶活性的调控代谢可能在协同PFKM基因调控糖酵解代谢[18]。
采用荧光定量的方法分析PFKM基因在长白猪、梅山猪等猪肌肉部位中分布,PFKM基因在猪的心肌以及骨骼肌中具有较高的相对表达量,在卵巢、子宫、脂肪组织和肾脏表达程度中等,但在其肝脏、睾丸、脾脏和肺中表达量相对较低,在小肠和胃中没有发现PFKM基因的表达。Wang Jun等[19]运用荧光定量PCR技术得出PFKM基因在猪不同部位的表达,尤其在心肌中,PFKM基因的表达量相对较高。与以上结果相似,PFKM基因在伊犁马的心脏组织中表达显著高于肝脏等其它组织。并且在伊犁马的肝脏组织中PFKM基因的表达要显著低于臀中肌、腹外斜肌、半腱肌、斜方肌、背阔肌、夹肌等组织。PFKM基因可能在伊犁马的糖酵解过程中,通过心脏组织参与PFKM基因的重要调控机制。孙慧等[20]通过生物信息学网站,针对综合质量评分最高的微小RNA进行进一步的检测以及验证,预测可能调控PFKM基因的微小RNA,并在PFKM基因的3’-UTR区域,推测出miR-141-3p可能是影响PFKM基因在不同组织中的的表达量进而改变生命细胞糖酵解的过程,控制其不同组织的发育水平。在转基因小鼠的骨骼肌中发现PFKM基因在低表达时检测到ATP能量水平很低、糖原含量高,致使小鼠的大部分组织的肌肉纤维改变甚至坏死,并且致使PFKM基因相关酶活性降低了大概50%左右,使红细胞渗透脆性增大,结果表明6-磷酸葡萄糖在肌肉中的积累,与人类GSDVⅡ的肌病和代偿性溶血特征具有相似性[21]。试验中伊犁马的PFKM基因在心脏组织中高表达,可能是伊犁马具有强大的心肌纤维活力,在6-磷酸果糖激酶的作用下,调控PFKM基因的正常表达,更好的分解、利用ATP,为其生命活动提供能量。Gibb等[22]利用大鼠原代的心肌细胞作为试验对象,并用双CMV启动子的磷酸酶缺陷型的病毒分别感染心肌,根据同位素示踪法得到,在PFKM活性较高的细胞中可以间接的限制线粒体分解以及细胞呼吸作用来增强糖酵解活动。在糖酵解过程中,PFKM的表达与动物的心肌细胞反应密切相关,该基因可能对心肌细胞进行糖酵解有重要的影响。
4 结 论
PFKM基因在伊犁马的不同组织中均有表达,且存在一定的差异性,其中心脏表达高达22.88。6-PFK的含量在伊犁马不同组织的含量也存在显著差异性,在心脏组织中的含量约为1.049 mg/L也显著高于其他组织。PFKM基因和6-PFK可能在糖酵解过程中起着重要作用,PFKM基因可能通过影响6-PFK的酶活,对心脏组织进行糖酵解有重要影响,PFKM基因和6-PFK在马匹的糖酵解过程中有重要的作用。
参考文献(References)
[1] 哈尼巴提·沙布开. 发挥优势凝心聚力让新疆现代马业走向世界—在2016年“中国·天山论马”论坛上的讲话[J]. 新疆畜牧业, 2016,(9): 4-7.
Hanibati Shabukai. Play the advantages to make Xinjiang modern horse industry to the world—Speech at the "China Tianshan Mountain on the Horse" Forum in 2016 [J].XinjiangXumuye, 2016,(9): 4-7.
[2]王建文, 李林玲, 王镜力, 等. 伊犁马ACTN3基因外显子多态性研究[J].畜牧兽医学报, 2018, 49(6): 1299-1306.
WANG Jianwen, LI Linling, WANG Jingli, et al. Analysis on ACTN3 Gene Exon Polymorphism of Yili Horse [J].ActaVeterinariaEtZootechnicaSinica, 2018,49(6): 1299-1306.
[3]叶再华, 姚新奎, 潘小东, 等. 伊犁马屠宰性能测定及适宜屠宰年龄研究[J]. 新疆农业大学学报, 2008,(3): 13-15.
YE Zaihua, YAO Xinkui, PAN Xiaodong, et al. Determination of slaughter performance and study on suitable slaughter age of Yili horse [J].JournalofXinjiangAgriculturalUniversity, 2008,(3): 13-15.
[4] Gibb A A, Epstein P N, Uchida S, et al. Exercise-Induced Changes in Glucose Metabolism Promote Physiological Cardiac Growth[J].Circulation, 2017, 136(22): 2144-2157.
[5] Ausina P, Silva D, Majerowicz D, et al. Insulin specifically regulates expression of liver and muscle phosphofructokinase isoforms[J].Biomedicine&Pharmacotherapy=Biomedecine&Pharmacotherapie, 2018, 103(4):228-233.
[6] Sun S, Li H, Chen J, Qian Q, Lactic acid: no longer an inert and end-product of glycolysis [J].Physiology(Bethesda) 2017, 32(6): 453-463.
[7] Webb B, Forouhar F, Szu F, et al. Structures of human phosphofructokinase-1 and atomic basis of cancer-associated mutations [J].Nature, 2015, 523(7558): 111-114.
[8]胡萍, 韦芳, 顾青, 等. 6-磷酸果糖激酶-2/果糖-2,6-二磷酸酶(PFKFB3)抑制剂3PO对高糖环境下人脐静脉血管内皮细胞新生血管形成的影响[J]. 眼科新进展, 2019, 39(5): 428-432.
HU Ping, WEI Fang, GU Qing, et al. Effect of 6-phosphofructokinase-2/fructose-2,6-bisphosphatase (PFKFB3) inhibitor 3PO on angiogenesis of human umbilical vein endothelial cells in high glucose environment[J].Newadvancesinophthalmology, 2019,39(5): 428-432.
[9] Wang J, Qin L, Feng Y, et al. Molecular Characterization, Expression Profile, and Association Study with Meat Quality Traits of PorcinePFKMGene[J].AppliedBiochemistry&Biotechnology, 2014, 173(7): 1640-1651.
[10]Matarneh S K, Yen C N, Elgin J M, et al. Phosphofructokinase and mitochondria partially explain the high ultimate pH of broiler pectoralis major muscle [J].PoultryScience, 2018, 97(5): 1808-1817.
[11]Wang J, Qin L, Feng Y, et al. Molecular Characterization, Expression Profile, and Association Study with Meat Quality Traits of PorcinePFKMGene[J].AppliedBiochemistry&Biotechnology, 2014, 173(7): 1640-1651.
[12]Wenting W, Hongxiang L, Jiamei X, et al. Aqueous Extract of Black Maca Prevents Metabolism Disorder via Regulating the Glycolysis/Gluconeogenesis-TCA Cycle and PPARα Signaling Activation in Golden Hamsters Fed a High-Fat, High-Fructose Diet[J].FrontiersinPharmacology, 2018, 9(4): 333-345.
[13]赵一萍,康德措,杜明,等.蒙古马免疫组织NF-KB信号通路相关基因的表达研究[J].中国畜牧杂志,2022,58(2):78-81.
ZHAO yiping, KANG decao, DU Min, et al. Study on expression of NF-kb signaling pathway related genes in Mongolian horse[J].ChineseJournalofAnimalScience, 2022, 58(2): 78-81.
[14] Moreno-Sánchez R, Marín-Hernández A, Gallardo-Pérez J, et al. Phospho fructokinase type 1 kinetics, isoform expression, and gene polymorphisms in cancer cells[J].JournalofCellularBiochemistry, 2012, 113(5):1692-1703.
[15] Xiong X, Ke X, Wang L, et al. Splice variant of growth hormone-releasing hormone receptor drives esophageal squamous cell carcinoma conferring a therapeutic target [J].ProceedingsoftheNationalAcademyofSciences, 2020, 117(12): 13433-13447.
[16] Ahsan H, Halpern J, Kibriya M G, et al. A genome-wide association study of early-onset breast cancer identifiesPFKMas a novel breast cancer gene and supports a common genetic spectrum for breast cancer at any age [J].CancerEpidemiologyBiomarkers&Prevention, 2014,(4): 23-24.
[17] Tang H, Lee M, Sharpe O, et al. Oxidative stress-responsive microRNA-320 regulates glycolysis in diverse biological systems [J].FasebJournalOfficialPublicationoftheFederationofAmericanSocietiesforExperimentalBiology, 2012, 26(11):4710-4721.
[18]卢建雄,陈粉粉,杨公社.调控糖酵解和生脂的重要转录因子:碳水化合物反应元件结合蛋白[J].生理科学进展,2006,(3):266-269.
LU Jianxiong, CHEN Fanfen, YANG Gongshe. Important transcription factors regulating glycolysis and lipid formation: carbohydrate response element binding protein [J].AdvancesinPhysiologicalScience, 2006,(3): 266-269.
[19] Wang J, Qin L, Feng Y, et al. Molecular Characterization, Expression Profile, and Association Study with Meat Quality Traits of PorcinePFKMGene[J].AppliedBiochemistry&Biotechnology, 2014, 173(7): 1640-1651.
[20]孙慧.miR-141-3p靶向PFKM影响细胞糖代谢参与调控间充质干细胞衰老的作用研究[D].长春:吉林大学,2021.
SUN Hui.MIR-141-3PtargetingPFKMaffectscellglucosemetabolisminregulationofMesenchymalstemcellaging[D]. Changchun: Jilin University, 2021.
[21]M García, Pujol A, Ruzo A, et al. Phosphofructo-1-Kinase Deficiency Leads to a Severe Cardiac and Hematological Disorder in Addition to Skeletal Muscle Glycogenosis [J].PlosGenetics, 2009, 5(8): 36-42.
[22] Gibb A A, Lorkie P K, Zheng Y T, et al. Integration of flux measurements to resolve changes in anabolic and catabolic metabolism in cardiac myocytes [J].BiochemicalJournal, 2017, 474(16): 2785-2801.
