盐胁迫下不同葡萄砧木的渗透调节及抗氧化能力
2023-01-04陈泽平史晓敏王振平
陈泽平,史晓敏,王 瑞,吴 轩,王 宁,王振平
(宁夏大学 农学院,银川 750021)
葡萄(Vitisvinifera)具有重要的营养价值、药用价值、经济价值,已成为中国种植最广泛的果树树种之一。中国西北的干旱、半干旱地区是葡萄发展的重要产区,是酿酒葡萄生长的黄金地带。据统计,中国盐碱化土地面积约为1亿 hm2,这严重制约着葡萄产业的发展[1]。付晴晴等[2]研究表明对于葡萄而言,选择耐盐性强的砧木嫁接栽培品种,能够提高葡萄砧穗组合的耐盐性。
邵红雨等[3]报道,高盐度土壤会影响植物生长,导致渗透胁迫、离子失衡和细胞氧化损伤。盐胁迫下渗透调节物质的积累是非盐生植物对盐胁迫的一种有效生理响应机制。植物体中主要的渗透调节物质有脯氨酸、可溶性糖及可溶性蛋白等。樊怀福等[4]研究表明在盐胁迫下植物体内活性氧(ROS)
宁夏贺兰山东麓葡萄栽培气候区年土壤水分蒸发量大于年降雨量,其盐渍化问题尤为严重[7]。在众多的生物措施中,种植耐盐碱植物是改良盐碱地的最佳措施之一。目前,葡萄砧木嫁接苗的抗旱性、抗寒性等方面已经得到深入研究,但关于盐胁迫对不同葡萄砧木影响的研究较少,为初步解释葡萄砧木耐盐性机理,本试验研究了盐胁迫对6种葡萄砧木渗透调节及抗氧化能力的影响,并利用隶属函数法综合评价砧木的耐盐性,筛选出适宜西北盐渍化地区栽培的品种,为葡萄的嫁接栽培提供参考依据。
1 材料和方法
1.1 试验材料及处理
试验于2020年5月到9月在宁夏永宁县玉泉营农场国家葡萄产业体系水分生理与节水栽培岗位试验基地的避雨大棚中进行。试验材料为13种一年生葡萄砧木:‘贝达’、‘5BB’、‘101-14’、‘110R’、‘188-08’、‘3309C’、‘140R’、‘Valiant’、‘山河2号’、‘B.R.NO.2’、‘Dogridge’、‘香槟尼’、‘5C’。
本试验为盆栽试验,于2020年5月9日将供试砧木留两芽修剪,根系剪留5 cm左右蘸适量生根粉,定植于底径200 mm、顶径280 mm、高200 mm的塑料花盆中。生长基质由蛭石、珍珠岩和草木灰混合而成(V蛭石∶V珍珠岩∶V草木灰为1∶1∶1),每盆1株,基质含量为8 L,花盆底下放置托盘以防溶液外渗,污染环境。定植后灌透水,砧木发芽后每隔3 d浇灌1/2 Hoagland’s营养液。待砧木长至20片完全展开叶后,选取9株长势一致的苗木进行盐胁迫处理,每3株为一个生物学重复。
处理方法参照付晴晴[8],在中度盐胁迫(100 mmol/L NaCl)处理6 d后表现出盐害症状,因此用含100 mmol/L NaCl的1/2 Hoagland’s营养液进行浇灌处理 (SALT),记为实验处理的第1天,随后每3 d浇灌1次,每次每盆浇1 L。对照浇灌相同体积的不含NaCl的1/2 Hoagland’s营养液(CK),持续处理6次后各砧木品种表现差异明显。于盐胁迫开始3 d、9 d、15 d后取各砧木品种7~9节完全展开叶及根系,用去离子水洗净、滤纸擦干、液氮速冻,于-80 ℃ 冰箱中保存备用。试验期间的平均最低和最高气温分别为24.2 ℃ 和37.5 ℃,平均最低和最高相对湿度分别为32.5%和55.2%,平均日照时间为12.6 h。
1.2 测定项目及方法
1.2.1 相对电导率葡萄砧木叶片的相对电导率(RC)参照徐新娟等[9]方法进行测定。于盐胁迫处理的第3、9、15天取各砧木叶片。取样后用蒸馏水洗净擦干叶片,然后用打孔器避开主脉取直径5 mm的叶片,每个品种取20片小圆片,将打好的叶片放入有20 mL蒸馏水的离心管中,浸泡4 h。浸泡结束后用雷磁DDS-307A型电导率仪测定溶液的电导率,记为R1;然后将离心管放入沸水浴中加热15 min,待溶液冷却至室温时测定溶液电导率,记为R2;每样品测定3个重复。最后计算相对电导率(RC=R1/R2×100%)。
1.2.2 丙二醛含量丙二醛(MDA)含量的测定参照赵世杰等[10]方法。称取新鲜叶片和根系 0.2 g(FW),用3 mL 0.1%(w/v)三氯乙酸研磨成匀浆,定容至10 mL。提取液用离心机离心,转速为5 000 g/min,离心10 min。吸取1.5 mL上清液,加入1.5 mL 5%硫代巴比妥酸溶液,混匀后100 ℃水浴反应20 min后迅速冷却,5 000 g/min 离心10 min后取上清,分别于450、532和600 nm下比色测定吸光度值OD450、OD532和OD600。依据公式计算MDA含量(C)。C (μmol·L-1) = [6.452×(OD532- OD600) - 0.559×OD450]×10/(1.5×FW)。
1.2.3 超氧阴离子产生速率和过氧化氢含量超
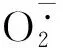
1.2.4 渗透调节物质含量葡萄砧木根系和叶片的可溶性蛋白含量参照高俊凤《植物生理学实验指导》[11]进行测定,可溶性糖含量的测定方法参照Rim Mzid[12]蒽酮硫酸法,脯氨酸含量的测定参照M. FOZOUNI[13]方法。
1.2.5 SOD和POD活性根系和叶片的SOD活性测定按南京建成试剂盒说明书进行(试剂盒货号A001-1),POD活性测定按李成龙等[14]方法进行。
1.3 葡萄砧木耐盐性综合评价采用隶属函数法、选取盐胁迫第15天各项生理指标进行葡萄砧木耐盐性综合评价。隶属函数值计算公式:Uij=(Xij-Xjmin)/(Xjmax-Xjmin)(正相关),Uij=1-(Xij-Xjmin)/(Xjmax-Xjmin)(负相关),式中:Uij表示i品种j指标的隶属函数值;Xij表示i品种j指标的测定值;Xjmin和Xjmax分别表示品种中j指标的最小值和最大值;i表示品种,j表示指标。根据上述公式先分别计算出各砧木8项指标的隶属度。然后取同种砧木8项指标的算术平均数作为平均隶属度使用。参照夏思哲等[15]的方法,按照平均隶属度将耐盐性分为3级:0.6~1.0为高耐盐(HR),0.4~0.6为中耐盐(MR),0~0.4为低耐盐(LR)。
1.4 数据处理
采用 Excel 2019 软件进行数据整理和作图,SPSS17.0 软件进行差异显著性与相关性分析。
2 结果与分析
2.1 盐胁迫下葡萄砧木叶片表观形态比较
在100 mmol/L NaCl 胁迫过程中,从叶片外观形态表现(图1)可以看出,各葡萄砧木的耐盐性差异明显。首先,各砧木品种叶片出现盐害的时间不同。其中,‘3309C’、‘140R’、‘山河2号’在第9天叶片出现褐斑、叶缘卷枯现象,在第15天叶片全面卷枯,几乎脱落;‘贝达’、‘5C’在第9~15天叶片出现褐斑,15 d后逐渐卷枯现象;‘101-14’、‘110R’在第15天叶片出现少量褐斑。其次,各砧木品种叶片遭受盐害程度不同。在盐胁迫处理15 d时,砧木品种‘188-08’、‘3309C’、‘140R’、‘Valiant’的叶片近乎完全脱落,且砧木生长受到显著抑制,说明其抗盐性极弱;砧木品种‘贝达’、‘5BB’、‘B.R.No.2’、‘Dogridge’有少部分叶片脱落,大部分叶片出现褐斑、叶缘卷枯现象,耐盐性较弱;砧木品种‘香槟尼’、‘5C’有较少的叶片出现叶缘卷枯现象,耐盐性较好;砧木品种‘101-14’、‘110R’除生长量较CK有所减少外,叶枯现象最少,说明其耐盐性较强,抗盐性最好。通过形态观察获得各品种砧木的实际耐盐性差异后,选择不同耐盐性水平的代表性品种‘贝达’、‘101-14’、‘110R’、‘3309C’、‘140R’、‘5C’进行后续研究,考察不同耐盐性品种相关抗逆性指标变化特征,探讨其耐盐机制。
2.2 盐胁迫对6种葡萄砧木相对电导率和MDA含量的影响
叶片相对电导率和MDA含量在一定程度上反映了外界环境条件胁迫下细胞质膜受损伤程度。随着盐胁迫处理天数的增加,各葡萄砧木叶片相对电导率呈逐渐增加的变化趋势(图2)。其中,在胁迫处理3 d时,对照组和胁迫组各品种叶片相对电导率分别在26.39%~37.49%和27.22%~42.44%之间,而‘贝达’、‘110R’、‘3309C’表现为胁迫组显著低于对照组,‘140R’则为胁迫组显著高于对照组39.58%,‘5C’、‘101-14’在两组之间均无显著差异。在胁迫处理9 d时,对照组和胁迫组各品种叶片相对电导率分别在25.49%~45.56%和27.62%~48.38%之间,仅‘5C’表现为胁迫组显著高于对照组,其余品种在两组间无显著差异。在胁迫处理15 d时,对照组和胁迫组各品种叶片相对电导率分别在38.56%~54.23%和42.48%~89.19%之间,除‘贝达’在两组间无显著差异外,其余品种均表现为胁迫组显著高于对照组,且‘3309C’、‘140R’的叶片相对电导率高达80%以上,表明其叶片细胞膜已受到损害,细胞液渗透严重。在胁迫处理18 d时,胁迫组各品种的相对电导率均显著高于对照组,其中的‘3309C’、‘140R’的相对电导率已达90%以上,叶片已经枯萎,其耐盐性极差;而此时‘110R’的电导率较15 d时降低,可能是该品种对盐胁迫适应后细胞膜有所修复,膜透性降低所致。由此可见,葡萄砧木品种‘贝达’、‘3309C’、‘140R’耐盐性较差,而‘101-14’、‘110R’耐盐性较好。

小写字母表示处理与对照间具有显著性差异(P<0.05),下同图2 盐胁迫下葡萄砧木叶片相对电导率的变化The different normal letters indicate significant difference between treatment and control.(P < 0.05), the same as belowFig.2 The relative electrical conductivity in leaves of grapevine rootstocks under salt stress
同时,盐胁迫下各品种砧木叶片和根系的MDA含量均随胁迫处理天数的增加而逐渐升高(图3)。

图3 盐胁迫下砧木叶片和根系丙二醛含量的变化Fig.3 The MDA content in leaves and roots of grapevine rootstocks under salt stress
在胁迫处理3 d时,胁迫组叶片MDA含量在‘贝达’、‘5C’中分别显著高于对照组32.67%、41.03%,其余品种与对照组均无显著差异;在胁迫处理9 d时,胁迫组MDA含量在‘140R’、‘贝达’中分别显著高于对照组68.75%、37.51%,在‘101-14’和‘3309C’中与对照组均无显著差异,在其余品种中显著低于对照组;在胁迫处理15 d时,胁迫组叶片MDA含量除‘3309C’与对照组无显著差异外,其余品种均比对照显著增加,在‘贝达’、‘110R’、‘5C’中增幅分别达到39.29%、52.99%、65.15%。同时,在胁迫处理3 d时,胁迫组根系MDA含量在‘贝达’、‘110R’、‘140R’中均较对照组显著增加,前两者增幅分别达到73.64%、61.10%,在‘5C’和‘3309C’中均显著低于对照组,而在‘101-14’中无显著差异;在胁迫处理9 d时,各品种胁迫组根系MDA含量均不同程度高于对照组,但仅‘5C’的增幅(83.27%)达到显著水平;在胁迫处理15 d时,各品种胁迫组根系MDA含量均较对照组显著增加,其中‘110R’和‘140R’增幅较大,分别达到67.70%、125.72%,而‘101-14’和‘3309C’增加较少,分别仅为15.42%和15.31%。
2.3 盐胁迫对6种葡萄砧木叶片和根系渗透调节物质含量的影响
2.3.1 脯氨酸含量在各盐胁迫处理阶段,各葡萄砧木间叶片和根系脯氨酸含量差异显著(表1)。其中,在盐胁迫处理3 d时,胁迫组砧木叶片脯氨酸含量仅‘贝达’和‘101-14’比对照组显著升高,升幅分别为34.58%和28.21%,其余变化均不显著;在胁迫处理9 d时,胁迫组脯氨酸含量在‘101-14’、‘贝达’、‘140R’中分别比对照组显著升高49.10%、39.00%和31.41%,在‘110R’和‘5C’中则比对照组显著降低;在胁迫处理15 d时,各品种胁迫组叶片脯氨酸含量均不同程度高于对照组,且除‘贝达’和‘101-14’外升幅均达到显著水平,并以‘5C’升幅最大(83.50%),其次是‘140R’(74.88%)。同时,在胁迫处理3 d时,胁迫组根系脯氨酸含量除‘贝达’和‘101-14’分别较对照组显著增加28.25%、40.96%外,其余品种均不同程度降低,但降幅均不显著;在胁迫处理9 d时,胁迫组脯氨酸含量除‘140R’和‘110R’分别比对照组显著升高219.42%、91.20%外,其余均不同程度降低,且‘5C’显著降低了52.17%;在胁迫处理15 d时,各品种胁迫组根系脯氨酸含量均高于对照组,且除‘110R’外均达到显著水平,其中的‘140R’、‘3309C’分别显著升高了106.70%、69.61%。
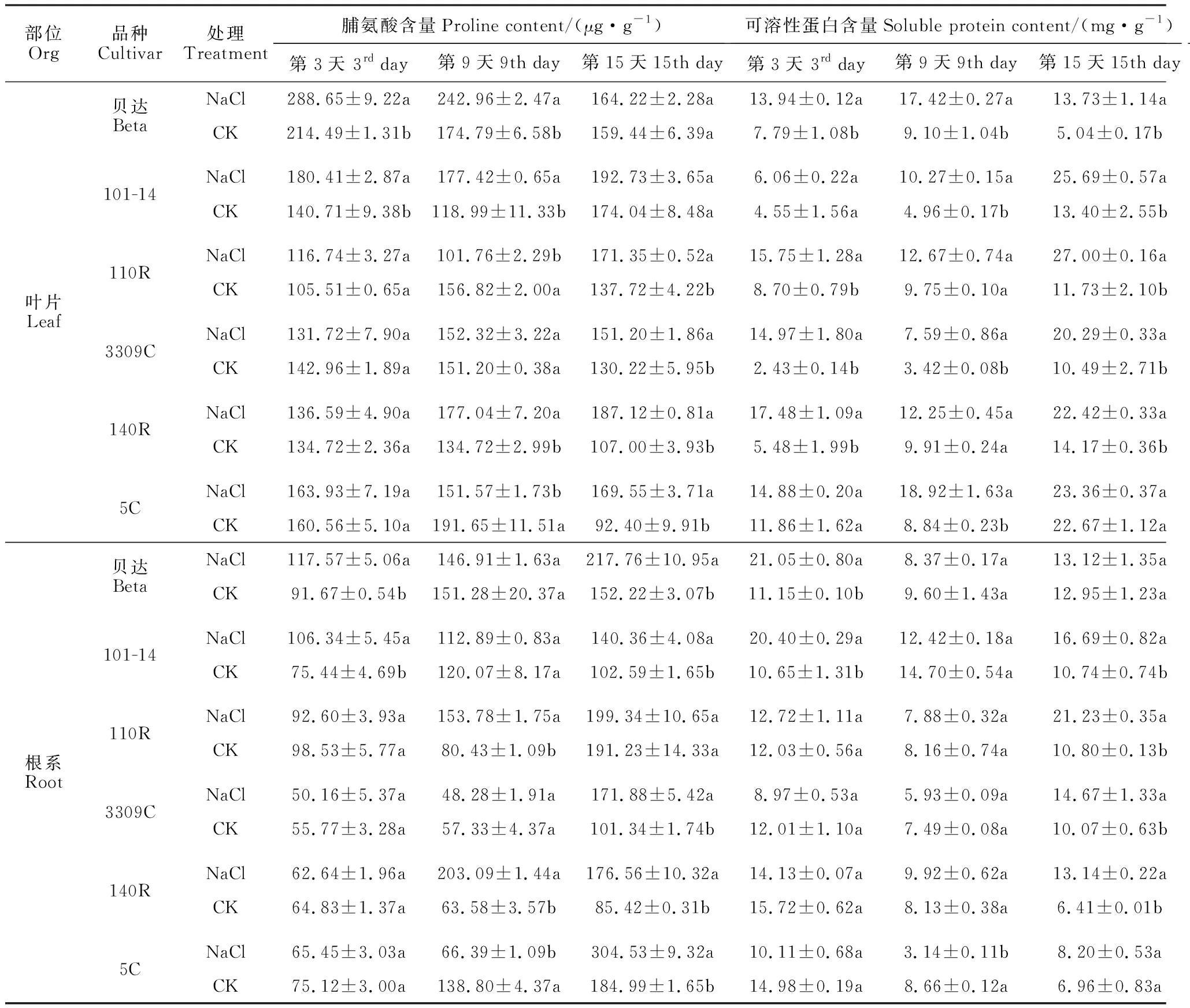
表1 盐胁迫下不同葡萄砧木叶片和根系渗透调节物质含量的变化
2.3.2 可溶性蛋白含量在各盐胁迫处理阶段,各葡萄砧木间叶片和根系可溶性蛋白含量表现出不同的增加趋势(表1)。各品种胁迫组砧木叶片可溶性蛋白含量均不同程度高于对照组,在胁迫处理3 d时,除‘101-14’和‘5C’外升幅均达到显著水平,并以‘3309C’升幅最大(516.05%),其次是‘140R’(218.98%);在胁迫处理9 d时,除‘110R’和‘140R’外升幅均达到显著水平,并以‘3309C’升幅最大(121.93%),其次是‘5C’(114.03%);在胁迫处理15d时,除‘5C’增加不显著,仅为3.04%外,其余品种均达到显著水平,以‘贝达’升幅最大(172.42%),其次是‘110R’(130.18%)。各砧木根系可溶性蛋白含量与叶片变化趋势不同,在胁迫处理3 d时,胁迫组根系可溶性蛋白含量显著增加的品种有‘101-14’、‘贝达’,增幅分别为91.55%、88.79%,而‘110R’增幅为5.74%,其余品种均不同程度降低;在胁迫处理9 d时,胁迫组根系可溶性蛋白含量仅‘140R’小幅升高22.02%外,其余品种均不同程度降低,且‘5C’显著降低了63.74%;在胁迫处理15 d时,胁迫组根系可溶性蛋白含量均有所增加,除‘贝达’和‘5C’外升幅均达到显著水平,以‘140R’升幅最大(104.99%)。其中叶片可溶性蛋白含量较高的品种,根系含量较少,说明各品种对盐胁迫存在时空特异性。
2.3.3 可溶性糖含量在各盐胁迫处理阶段,各葡萄砧木间叶片和根系可溶性糖含量差异显著(表1),且叶片和根系表现出不同的变化趋势。在胁迫处理3 d时,胁迫组砧木叶片可溶性糖含量除‘3309C’比对照组降低外,其余品种均比对照组不同程度升高,‘101-14’比对照组显著升高,升幅为15.18%;在胁迫处理9 d时,‘贝达’、‘110R’、‘5C’比对照组显著升高,升幅分别为31.00%、14.17%、20.24%,仅‘101-14’可溶性糖含量比对照组显著降低了25.49%,其余变化不显著;在胁迫处理15 d,胁迫组叶片可溶性糖含量均有所增加,显著增加的品种有‘101-14’,升幅最大(72.43%),其次是‘110R’(25.56%)。同时,胁迫组各砧木品种根系可溶性糖含量差异显著,在胁迫处理3 d时,除‘3309C’、‘5C’比对照组降低外,其余品种均比对照组显著升高,并以‘贝达’升幅最大(80.12%),其次是‘110R’(59.26%);在胁迫处理9 d时,胁迫组根系可溶性糖含量除‘101-14’比对照组增加不显著外,其余均显著增加,以‘140R’升幅最大(91.71%),其次是‘5C’(72.76%);在胁迫处理15 d时,除‘贝达’和‘3309C’比对照组降低外,其余品种均比对照组显著增加,并以‘140R’升幅最大(45.10%),其次是‘5C’(42.39%)。6个品种中只有‘110R’、‘101-14’叶片与根系表现出相对一致的增加趋势,其余品种地上部和地下部均有较大的差异性。
2.4 盐胁迫对6种葡萄砧木叶片和根系抗氧化系统的影响


图4 盐胁迫下砧木叶片和根系产生速率的变化Fig.4 The production rate in leaves and roots of grapevine rootstock under salt stress
2.4.2 H2O2含量图5显示,盐胁迫下各品种砧木叶片和根系的H2O2含量均大于相应对照,增幅大多达到显著水平,且叶片H2O2含量大于根系。其中,在胁迫处理3 d时,胁迫组叶片H2O2含量以‘5C’、‘101-14’增幅较大,分别较对照升高了122.76%、80.95%,而‘110R’增幅较小(14.71%);胁迫组根系H2O2含量以‘3309C’增幅最大(146.91%);在胁迫处理9 d和15 d时,各品种胁迫组叶片H2O2含量分别较对照组增加10.86%~66.58%和21.83%~60.45%,以‘5C’增幅较大,‘101-14’和‘140R’增幅较小;在胁迫处理15 d时,胁迫组‘101-14’、‘5C’和‘3309C’根系H2O2含量均显著高于对照组,前两者增幅分别为88.32%、74.51%,而胁迫组‘110R’和‘140R’则与对照组无显著性差异。可见,盐胁迫后H2O2含量变化在品种间表现出明显差异,大部分品种在盐胁迫后显著增加且H2O2含量处于相似的水平,个别品种具有特异性。
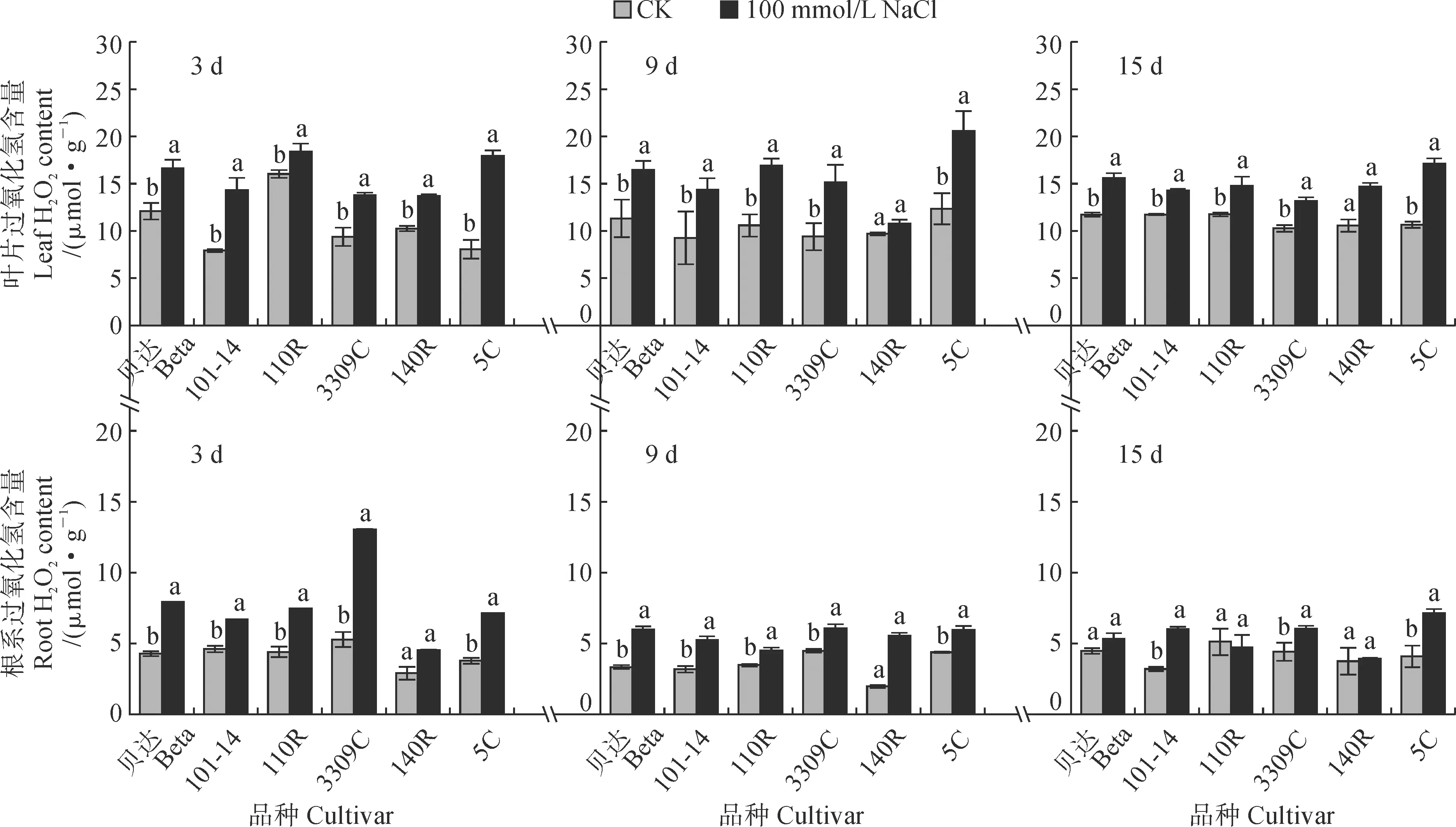
图5 盐胁迫下砧木叶片和根系H2O2含量的变化Fig.5 The H2O2 content in leaves and roots of grapevine rootstocks under salt stress
2.4 3 SOD活性SOD能够催化超氧阴离子自由基歧化生成氧和过氧化氢。图6显示,随着盐胁迫处理天数的增加,各品种砧木叶片的SOD活性总体表现为胁迫组高于对照组且大多达到显著水平,并整体呈先升高后降低的趋势,同期各个品种之间差异较小。其中,在胁迫处理3 d时,砧木叶片SOD活性较高的是‘贝达’、‘5C’,胁迫组较对照组分别显著增加了10.37%、22.82%;在胁迫处理9 d时,胁迫组砧木叶片SOD活性较大的是‘5C’,较对照组显著升高了23.66%,酶活性较低的仍然是‘3309C’和‘140R’,分别较对照组升高了6.76%和9.40%。
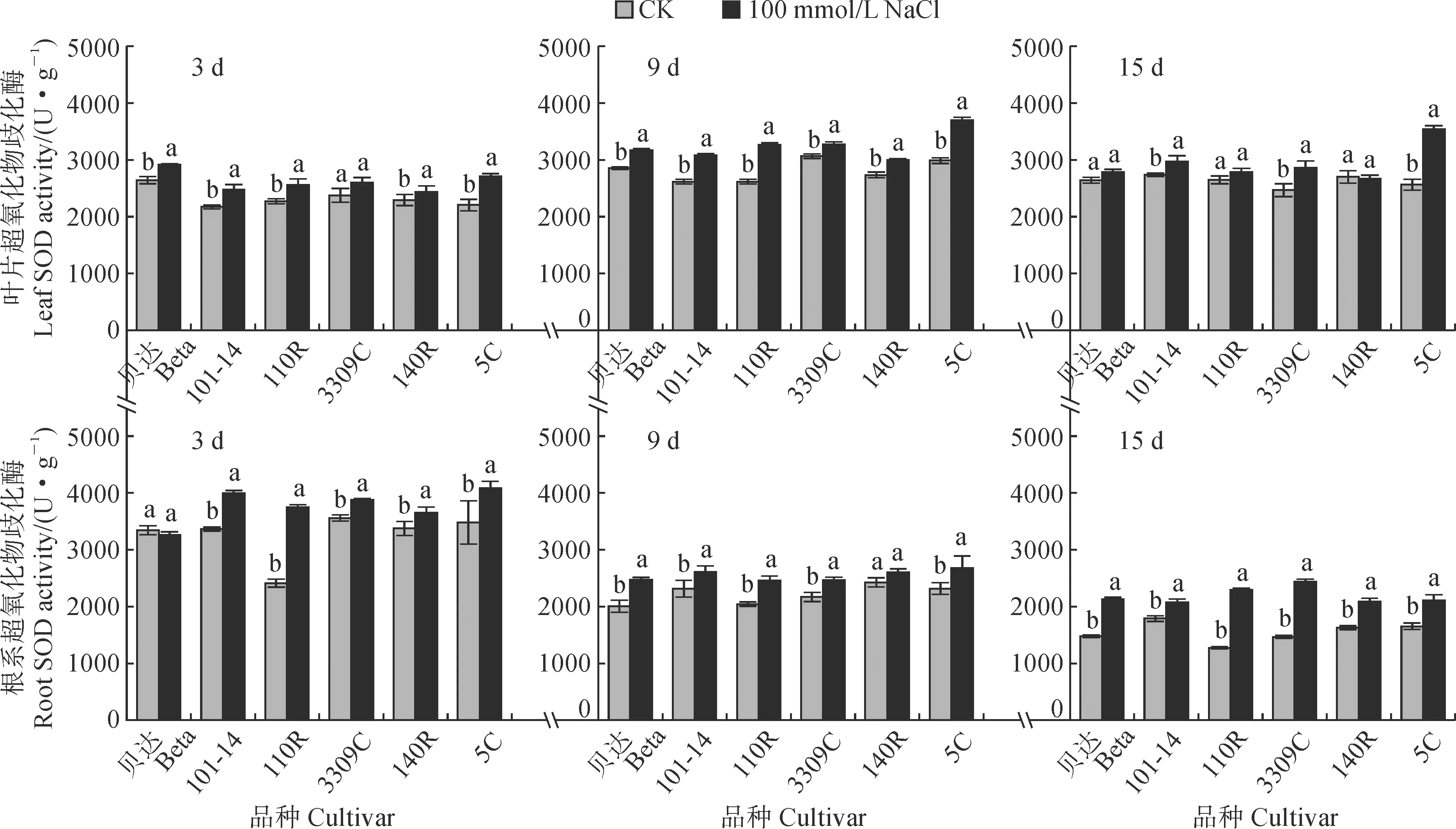
图6 盐胁迫下葡萄砧木叶片和根系SOD活性的变化Fig.6 The SOD activity in leaves and roots of grapevine rootstocks under salt stress
在胁迫处理15 d时,胁迫组叶片SOD活性仅有‘5C’保持了较高水平,较对照组显著增加38.02%,而‘贝达’、‘110R’、‘140R’叶片SOD活性均与对照组无显著差异。同时,各砧木品种根系的SOD活性与叶片表现出不同的变化趋势,均随胁迫时间呈逐渐降低,但是胁迫组大多仍比对照组显著增加。其中,在胁迫处理15 d时,‘110R’、‘3309C’、‘贝达’的根系SOD活性显著高于对照组,增幅分别增加到79.69%、65.90%、44.00%,而‘101-14’的酶活性则相对较弱,为15.72%。根系SOD活性在胁迫处理15 d时下降原因可能是盐胁迫时间过长,导致抗氧化酶系统受到了严重伤害。
2.4.4 POD活性POD是清除H2O2的重要保护酶。与SOD活性变化趋势不同,盐胁迫下各砧木叶片POD活性随胁迫天数的增加呈先增加后降低的变化趋势,且个别品种表现出较高的酶活性水平,其中品种‘101-14’的POD活性较高,但随胁迫时间的增加逐渐降低(图7)。在胁迫处理3 d时,除‘101-14’显著升高43.75%外,其余各砧木叶片POD活性与对照组差异不显著;在胁迫处理9 d时,各品种POD活性均比3 d时有所增加,且胁迫组大多显著高于对照组;在胁迫处理15 d时,胁迫组‘贝达’、‘110R’叶片酶活性较对照组分别显著降低63.64%、51.02%,其余品种无显著变化。同时,与叶片相比,根系POD活性在品种间相近,随着胁迫时间基本呈先升高后降低的变化趋势。在胁迫处理3 d和9 d时,除‘3309C’外,其余各砧木品种根系POD活性大多比对照组显著增加;在胁迫处理15 d时,各品种砧木根系POD活性除‘110R’、‘5C’比9 d时增加外,其余品种酶活性下降,且胁迫处理过程中,胁迫组‘101-14’、‘110R’、‘5C’根系的POD活性均显著高于对照组。结果表明不同品种砧木对盐胁迫的耐受性与响应程度存在较大差异,耐盐性较强品种的酶活性较高且维持时间较长。
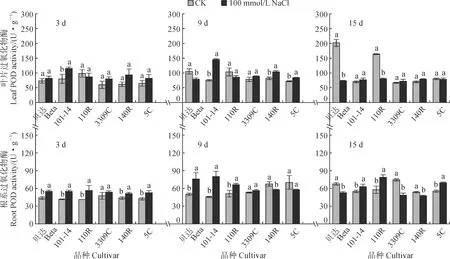
图7 盐胁迫下葡萄砧木叶片和根系POD活性的变化Fig.7 The POD activity in leaves and roots of grapevine rootstocks under salt stress
2.5 不同葡萄砧木耐盐性综合评价
依据盐胁迫第 15 天时各葡萄砧木叶片丙二醛含量、超氧阴离子产生速率、过氧化氢含量、可溶性蛋白含量、可溶性糖含量、脯氨酸含量、SOD活性和POD活性等 8 项生理指标,采用隶属函数法对6个葡萄砧木进行耐盐性综合评价(表2),依据各砧木的平均隶属度大小得出它们耐盐性排序为‘110R’>‘101-14’>‘贝达’>‘5C’>‘3309C’>‘140R’。与实际形态观察结果相比,高耐盐、中耐盐和低耐盐性品种的综合评价结果与其相符。

表2 盐胁迫第15天不同葡萄砧木耐盐性的隶属函数值
3 讨 论
葡萄作为宁夏地区重要的经济果树之一,生长过程中受到各种胁迫的影响,盐胁迫是限制其正常生长的非生物胁迫因子之一。丙二醛(MDA)是衡量氧化胁迫程度的常用指标之一,能反映植物膜脂过氧化的程度[16],因此,丙二醛含量也可以指示植物在盐胁迫下的耐盐性。本研究发现在盐胁迫处理下各葡萄砧木叶片丙二醛和相对电导率都显著增加,这与Liang[17]、Mahmoud[18]等研究结果一致。本研究中各砧木叶片相对电导率差异显著,耐盐性弱的砧木品种‘3309C’、‘140R’在胁迫初表现出较高的相对电导率,说明两者叶片细胞在盐胁迫开始就遭受破坏,且随着盐胁迫天数增加,相对电导率值逐渐升高,盐胁迫后期耐盐性弱的品种相对电导率值接近90%。植物在遭受盐胁迫后产生的这些膜脂过氧化产物通过对蛋白质、植物细胞膜系、植物细胞光合作用、植物呼吸作用等的伤害[19],进而影响植物的正常生长发育。
同时,盐胁迫引起的渗透胁迫会限制植物生长,植物遭受胁迫后其脯氨酸、可溶性糖及可溶性蛋白等可以通过保持细胞内外渗透势防止脱水,缓解胁迫过程中植物的衰老程度。田晓艳等[20]发现,牧草幼苗叶片可溶性糖、可溶性蛋白含量随着盐胁迫时间的增加而显著增加,从而平衡植株细胞渗透势,保持植株在胁迫条件下与外界交换水分,本研究结果与之一致。本试验中砧木根系脯氨酸含量在胁迫处理后期显著增加,这可能是由于盐胁迫造成了膜蛋白水解,从而使可溶性蛋白含量增加,说明胁迫后期细胞膜严重受损。本试验中葡萄砧木品种‘3309C’、‘140R’可溶性蛋白含量在胁迫开始时较高,出现盐害的时间最早,也是最早枯死的品种,说明其耐盐性较弱。另外,试验盐胁迫下各品种砧木脯氨酸含量较对照均有显著增加,这与Fozouni等[13]和罗巧玉等[21]研究结果一致。
另外,细胞膜具有重要的生理功能,它既维持稳定代谢的胞内环境,又能调节和选择物质进出细胞。细胞膜能接收外界信号的刺激使细胞做出反应,同时满足植物生理活动的需要[22]。氧气是植物生长发育过程中不可缺少的物质之一,它参与新陈代谢、线粒体呼吸和氧化磷酸化等代谢过程并产生能量。然而,氧在代谢过程中可能被激活成活性氧(ROS),ROS具有很强的氧化能力,可导致细胞质膜损伤、不可逆代谢功能障碍和细胞死亡[23],因此ROS含量的变化可以一定程度上反映植物伤害情况。本试验发现,耐盐能力差的葡萄砧木品种叶片ROS含量积累较多,而耐盐性强的品种则产生较少,其有害的超氧阴离子含量也较少。抗氧化酶可有效清除叶片中的ROS,从而缓解超氧阴离子过量带来的伤害[24]。抗氧化酶活性的提高有利于植物适应盐渍环境,SOD是ROS
耐盐性是评价砧木抗逆性的一个重要指标,它与盐胁迫下葡萄砧木叶片和根系中活性氧代谢、渗透调节物质和抗氧化酶活性密切相关,但采用单一指标很难做出准确判断。本试验依据盐胁迫下各葡萄砧木各项抗逆生理指标,利用隶属函数法综合评价砧木的耐盐性,结果表明砧木品种‘110R’、‘101-14’耐盐性强,这与袁军伟等[30]的研究结果一致,而与吴梦晓等[31]的砧木抗逆性鉴定结果不同,猜测可能是取样时间、部位不同所致,也可能是盐处理、栽培方式等因素引起的,今后需进一步研究证实。
综上所述,在中度盐胁迫条件下,葡萄砧木叶片细胞液外渗,细胞膜透性增加。‘110R’、‘101-14’等耐盐性较强的品种能产生较多的可溶性蛋白、可溶性糖、脯氨酸等渗透调节物质和提高自身SOD、POD等抗氧化酶活性,维持细胞内稳态和细胞膜的稳定性,从而表现出一定的耐盐胁迫能力。在本研究含100 mmol/L NaCl的1/2 Hoagland’s营养液胁迫条件下,‘110R’和‘101-14’砧木品种耐盐性较强,可作为高盐土壤葡萄砧木使用,适合在西北地区的推广与应用。
