鸡白痢沙门菌感染雏鸡的骨髓miRNA表达谱分析
2023-01-03潘孝成沈学怀张丹俊王洁茹
尹 磊,潘孝成*,沈学怀,张丹俊,戴 银,王洁茹
(1.安徽省农科院畜牧兽医研究所 安徽省畜禽疫病研究中心,合肥 230031;2.畜禽产品安全工程安徽省重点实验室,合肥 230031)
鸡白痢沙门菌(S. Pullorum)是禽类的重要病原体,对家禽业构成严重威胁。鸡白痢沙门菌可利用大囊泡(Salmonellacontaining vacuole, SCV)有效逃避宿主免疫系统,使宿主长期保持持续或隐性感染,造成机体持续带菌且细菌长期定植于脾巨噬细胞或生殖道内,通过血液循环进入全身各组织器官,进而导致免疫抑制和免疫系统损害,引起严重的系统性疾病。相较于其他种属沙门菌,鸡白痢沙门菌具有高度的宿主特异性,鸡最为易感,其次是火鸡。它能引起雏鸡白痢并引发急性败血症,造成较高的发病率和死亡率[1]。尽管S. Pullorum病在发达国家的商业鸡群中已得到了很好的控制,但在发展中国家,包括中国,仍然是一个主要问题[2]。因此,更全面地了解鸡对S. Pullorum的反应将有助于改进防控策略,并为有效控制和预防S.Pullorum病提供新的理论基础。
骨髓是家禽中最重要的器官之一[3],它在抵抗和抑制细菌及病毒等各种病原体中发挥着重要作用。骨髓不仅是多能造血干细胞的重要来源,而且也是两大类白细胞,即淋巴系和骨髓系的储备库。其中,淋巴系分化为B细胞、T细胞和自然杀伤细胞,而骨髓系则发展为巨噬细胞、粒细胞、肥大细胞和树突状细胞(DCs)。所有这些细胞在先天性和适应性免疫反应中发挥关键作用[4-5]。目前,通过组学技术对免疫器官或免疫细胞在感染期间进行基因差异表达分析,有助于理解S. Pullorum致病机制和免疫调控水平。
微小核糖核酸(miRNA)是一类20至23个核苷酸的非编码小RNA,作为生物过程的转录后调节器,在调控发育、分化、器官形成、生长控制和细胞凋亡等生命活动中发挥重要作用[6]。研究表明,miRNA在微生物感染宿主过程中发挥着重要作用,如禽致病性大肠杆菌感染鸡后,脾miRNA的表达失调与免疫细胞的发育分化及炎症反应密切相关[7]。骨髓提供的原始细胞,即两大类白细胞,不受淋巴器官中存在的发育细胞因子和其他因素的影响[8],因此,骨髓可以用来识别感染S. Pullorum的新的候选调控分子和网络。本研究对感染和未感染鸡骨髓中不同的miRNA表达进行分析,以确定骨髓对S. Pullorum感染的免疫反应机制。
1 材料与方法
1.1 实验动物与样品采集
无特异性病原体(SPF)的来航鸡的胚胎蛋购自北京梅里亚实验动物技术有限公司。SPF鸡在封闭环境中孵化,并在负压隔离器中饲养,以保证动物实验中无沙门菌。在7日龄时,30只SPF鸡口服1 mL 108cfu的S. Pullorum (CVCC 2216)作为感染组,另外15只鸡口服1 mL PBS作为对照组,感染组与对照组分别单独隔离饲养。在感染后24 h内,对鸡静脉注射巴比妥钠安乐死。用解剖刀去除鸡腿骨上的结缔组织,然后用刀将骨剖开,用刮刀将其中的骨髓取出,最后用1.05 mm的尼龙网过滤,得到纯骨髓,快速冷冻并保存在-80 ℃。
1.2 总RNA提取及样品准备
使用TRIzol试剂(Invitrogen)和Polyacryl Carrier(MRC)从样品中提取总RNA。在1%琼脂糖凝胶上监测RNA的降解和污染情况。使用NanoPhotometer分光光度计(IMPLEN)、Aglient Bioanalyzer 2100系统的RNA Nano 6000检测试剂盒(Aligent Technologies)和Qubit 2.0 Flurometer的Qubit RNA检测试剂盒(Life Technologies)测量RNA纯度、完整性和浓度。只有吸光度(260/280 nm)之比>1.8和RNA完整性编号(RINs)>7的RNA样品被用于RNA分析。
1.3 miRNA测序及生物学功能分析
由Oebiotech公司(中国上海,http://www.oebiotech.com)对样品进行microRNA分析。提取的RNA被标记并杂交到Agilent-070154 RatmiRNA Microarray V21.0 8×15K(Agilent)。采用Genespring软件(13.1版,Agilent Technologies)对原始数据进行标准化,并识别差异表达的miRNAs(DEmiRNAs)。上调或下调的基因的阈值是倍数变化≥2和P≤0.05。通过检查两个数据库(Targetscan和microRNAorg)的重叠交集来选择DEmiRNAs的目标基因。使用基因本体论(GO)和京都基因和基因组百科全书(KEGG)对推测的基因进行了功能和途径富集分析。GO和KEGG分析的意义阈值被定义为P≤0.05。miRNA和靶基因之间的潜在调节关系使用Cytoscape软件(http://www.cytoscape.org/)进行分析[9]。
1.4 qRT-PCR分析miRNAs
使用ABI Step One热循环仪(Applied Biosystems, CA, USA)和miRcute miRNA qPCR SYBR Green检测试剂盒(Vazyme, Nanjing, China)进行qRT-PCR。本研究使用的miRNA特异性正向引物见表1。U6 snRNA被用来作为内部标准。每个基因都使用了3个独立的生物重复。每个miRNA的相对表达水平通过2-ΔΔct方法计算[10]。

表1 本研究所用引物序列
2 结 果
2.1 骨髓中差异表达的miRNA分析
对转录组数据通过P值≤0.05和差异倍数变化≥2的阈值进行过滤。根据这些标准,在鸡白痢沙门菌感染后的骨髓中,与对照组相比,发现了20个差异表达的已知miRNAs。其中11个上调,9个下调,通过对样本进行差异化处理的聚类热图,结果如图1所示。

图1 差异表达miRNAs聚类热图Fig.1 Heat map of differentially expressed miRNAs clustering
2.2 通过qRT-PCR验证miRNAs
为了验证miRNA测序结果的可靠性,随机选取6个关键差异显著的miRNAs进行验证,以U6作为内参基因,根据NCBI中的基因序列设计引物,通过qRT-PCR检测,结果如图2所示,与未感染组相比,3个miRNAs(gga-miR-6643-5p、gga-miR-1466和gga-miR-2954)表达上调,3个miRNAs(gga-miR-762、gga-miR-6690-5p和gga-miR-1647)表达下调,测序结果与qRT-PCR趋势一致,表明转录组数据可信。
2.3 miRNA靶基因分析
利用miRanda算法预测差异表达miRNAs的潜在靶基因,并进行生物信息学分析。通过差异表达miRNAs预测靶基因并进行GO功能注释和富集分析,确定miRNA靶基因的功能。从图3可以看出,预测的靶基因主要注释到膜运输、信号转导、免疫系统、碳水化合物代谢、糖类的生物合成和代谢等等分子功能上(图3)。KEGG信号通路分析发现,差异表达miRNA的靶基因主要富集于Notch信号通路、Hedgehog信号通路、PPAR信号通路、AMPK信号通路、Hippo信号通路等信号通路(图4)。
2.4 与免疫相关的miRNA-mRNA调控网络互作分析
利用Cytoscape软件构建与免疫过程相关的miRNA-mRNA网络互作图。图5结果显示,有11个与免疫相关的差异表达的miRNAs,结合miRNAs与mRNA的连接性及所拥有的mRNA靶基因数量,发现gga-miR-1466和gga-miR-6643-5p可能是参与免疫过程相关的关键miRNA。

图2 qRT-PCR结果与miRNA测序结果的比较Fig.2 Comparison of the qRT-PCR results with the miRNA sequencing results
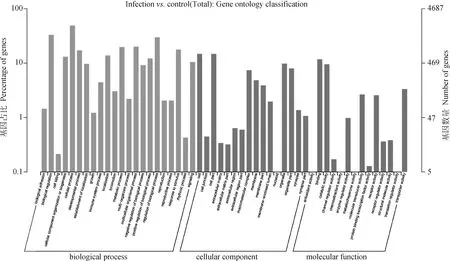
图3 差异表达miRNA靶基因GO分析Fig.3 GO analysis of differentially expressed miRNA target genes

图5 miRNA-mRNA互作网络图Fig.5 miRNA-mRNA interactions network map
3 讨 论
先天免疫是抵御病原体入侵的第一道防线,当病原体相关分子模式(PAMPs)被模式识别受体(PRRs)识别时就会被激活[11]。骨髓作为未成熟免疫细胞的储存库,它在抵抗病原菌感染过程中发挥着重要作用。鸡白痢沙门菌作为一种重要的宿主特异性致病菌,只在鸡中引起疾病。目前,关于骨髓细胞在应对鸡白痢沙门菌感染过程中的作用及机制未见报道。因此,本研究利用miRNA-seq技术获得了感染S. Pullorum与未感染鸡的骨髓转录表达谱测序数据。随后,基于对骨髓中miRNA和潜在靶标mRNA表达谱的联合分析,目的是更好地了解宿主在抵抗S. Pullorum感染过程中的免疫机制。
miRNAs已被证明与病原体入侵和宿主抵抗力密切相关[12-13]。因此,有必要对鸡感染S. Pullorum等病原体免疫反应中的关键miRNAs进行鉴定和定性,以了解发病机制,改善动物福利,减少家禽生产中的损失,保证食品安全。本研究中miRNA表达谱数据显示,共获得20个已知的差异表达miRNAs,其中11个表达上调,9个表达下调,其中与免疫相关的差异表达miRNAs有11个,包括gaa-miR-762、gaa-miR-2954、gaa-miR-1647等。研究表明,gaa-miR-762的表达变化与免疫系统和炎性损伤密切相关,在病原体感染宿主过程中,LPS诱导的骨髓衍生巨噬细胞诱导特定的miRNAs上调,这其中包括gaa-miR-762,进而导致炎性炎症细胞因子(IL-6、IL-12和TNF)的产生减少,但它们增加了IL-10的释放[14-15];Zhou等[16]通过注射地塞米松(Dex)建立了7日龄鸡的免疫抑制模型,分析了鸡胸腺中miRNA表达谱特征,发现gaa-miR-2954高度富集,通过影响Toll样受体信号通路、Jak-STAT信号通路等免疫信号通路进而影响鸡的胸腺免疫功能;gaa-miR-1647参与禽巨噬细胞的表达分化,在应对病原体时能够成功激活相关通路从而发挥重要作用[17]。总之,本试验结果表明,这些差异表达的miRNAs可能参与了宿主骨髓细胞与S. Pullorum的相互作用,具体机制有待后续进一步的研究。
通过预测miRNAs的靶点和注释其生物功能,对预测miRNAs的功能和构建调控网络很有帮助。本研究中,11个与免疫有关的生物过程被显著富集,同时,发现富集的基因主要集中在Notch信号通路、Hedgehog信号通路、PPAR信号通路、AMPK信号通路、Hippo信号通路上。Notch信号通路在控制淋巴细胞介导的炎症反应中发挥着重要作用[18];Hedgehog信号通路相关基因的表达失调会导致巨噬细胞的组织特异性炎症[19];PPAR信号通路可以被Genipin激活,引导并维持巨噬细胞向M2亚型的极化,从而缓解炎症反应[20];AMPK信号通路作为能量应激和钙诱导自噬的关键调节因子,它的激活对宿主在炎症反应下的损伤有保护作用[21];Hippo通路作为先天免疫中最原始的信号通路,其信号成分LATS2通过PQBP1-cGAS途径增强先天免疫,从而抑制病原微生物的感染[22]。本试验结果突出了差异表达的miRNAs在宿主对S. Pullorum感染反应中的潜在功能。
当前,鸡白痢沙门菌所造成的经济损失对中国养禽业仍是一个重大的挑战,需要高效精准的防控解决方案。miRNA在S. Pullorum感染致病过程中发挥着重要作用,从miRNA水平解析S. Pullorum的致病机制,有助于发现防控S. Pullorum的新途径。本研究发现,gaa-miR-1647在S. Pullorum感染过程中显著差异表达,gaa-miR-1647作为激活免疫细胞应对病原感染的关键因子[17],可以考虑利用gaa-miR-1647作为诊断鸡白痢沙门菌感染的生物标志物,有待后续进一步的验证与研究。同时,miRNA靶向宿主基因作为新型治疗靶标的研究方向,通过将聚合物纳米技术与miRNA调控技术相结合,开发出一种靶向聚合物纳米颗粒[23],将包裹着gaa-miR-762和gaa-miR-2954共同递送到鸡体内,通过调节Toll样受体信号通路、Jak-STAT信号通路和炎症信号通路,实现对S. Pullorum的防控。
4 结 论
本试验通过miRNA测序来筛选和识别S. Pullorum感染鸡骨髓的差异表达miRNAs,这些差异表达的miRNAs与免疫和炎症反应的信号通路有关,本研究为阐明S. Pullorum致病机制和病原与宿主的相互作用提供了基础,也为防控S. Pullorum提供了新的策略。
