人参内生菌的研究进展*
2022-12-29凡贞洁徐静雅侯典云胡灏禹肖水明
凡贞洁,徐静雅,侯典云,胡灏禹,肖水明**,徐 江
(1.中国中医科学院中药研究所 北京 100700;2.河南科技大学农学院 洛阳 471000)
内生菌最早由De Bary在1866年提出[1],用来定义生活在植物组织内的微生物。广泛使用的内生菌定义是由Bacon和White提出的:定居在植物的活体内部组织而不会造成任何直接的、明显的病害症状的微生物[2]。植物内生菌种类丰富,分布广[1-3]。由于它们与宿主植物协同进化[4-5],有了一些特殊的生物学功能,如间接促进植物生长[6-7]、产生与宿主相似或相同的次生代谢产物[6-8]。
人参(Panax ginsengC.A.Mey.)是人参属多年生草本植物[9],生长环境潮湿阴凉,主产于中国东北地区,在朝鲜北部、日本等地也少有分布[10]。《神农本草经》中早有记载,人参可治疗肺虚喘促、脾虚泄泻等症,具有益血生津,宁神益智等功效,被誉为百草之王[11-12]。人参含有人参皂苷、多糖、甾醇、氨基酸等多种化学成分,具有抑菌、抗肿瘤、抗氧化、抗过敏等多种药理作用[13-15];目前,人参基因组已进行测序和分析,为研究不同药理活性成分的生物合成途径奠定基础[16]。病虫害、根际微生物影响、生长周期长等会制约人参的产量和品质[17-20];而植物内生菌具有促进植物生长、提高植物抗逆性的作用,可以提高植物产量[21-22]。因此,内生菌在农业生产中具有重要的应用潜力。
文章主要对人参内生菌国内外的最新研究进行综述概括,并对未来人参内生菌的研究发展做出展望,以期为今后人参在农业生产、医药治疗方面提供依据,为今后药用植物微生物的研究方向以及中医药的发展提供参考。
1 人参内生菌分离与鉴定研究方法
研究内生菌的方法包括传统分离鉴定的培养法和基于分子生物学技术的非培养法。内生菌具有一定程度的组织偏好性,应选取植物的不同组织进行内生菌研究[3]。若采用组织培养法分离内生菌,不同类型的内生菌所用培养基一般不同,内生细菌分离一般采用营养琼脂(Nutrient Agar,NA)培养基,内生真菌分离一般采用马铃薯葡萄糖琼脂(Potato Dextrose Agar,PDA)培养基,内生放线菌分离一般采用高氏I号培养基。优点:得到新菌种资源;对分离得到的内生菌进行表征研究。缺点:分离结果易受培养基种类的限制,分离较缓慢。
分离培养方法由于所用培养基不能完全满足微生物的生长需求,且大多数微生物的培养条件人们还不了解或了解很少,导致分离到的微生物种类很少,不能全面客观的反映植物微生态系统中微生物的真实状况。以高通量测序技术为基础的非培养法则能对微生物进行较客观的分析,较精确地揭示样品中微生物种类和遗传多样性。通过高通量测序技术在人参基因组方面也取得了一定的研究成果,人参全基因组序列的获得对人参的选育和栽培具有重大意义[23-24]。二代测序技术具有速度快、准确性高、易操作等优点,三代测序技术(如:PacBio SMRT)则具有可以在物种级别进行分类的优势[25]。优点:效率高、测序得到的内生菌覆盖面更广。缺点:不能对具体菌株进行表征研究,无法保留菌种资源。由表1可知,传统培养分离方法和高通量测序技术相结合,为微生物的功能研究以及有益微生物的筛选提供了数据。
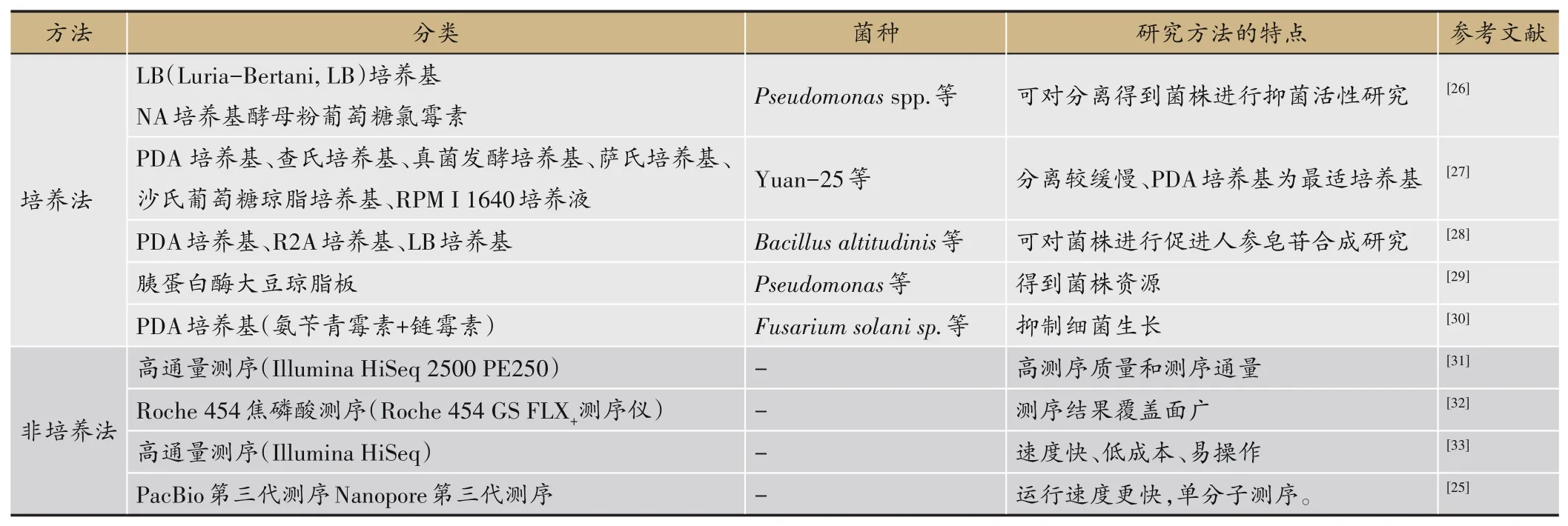
表1 人参内生菌分离与鉴定研究方法
2 人参内生菌的多样性
2.1 不同器官内生菌的多样性
2.1.1根部内生菌的多样性
目前已从人参植株不同部位分离出数量众多、不同分类地位的内生菌[34]。李勇等[26]通过平板培养法从人参根中分离得到内生细菌10株,包括Pseudomonasspp.、Stenotrophomonassp.、Bacillusspp.等。Yan等[35]通过组织分离法共分离出了81株内生细菌,包括菌株PDA-2、LB-5和R2A-7等。邱服斌[36]从3年生人参根中 分 离 得 到Bacillussp.、Pseudomonassp.、农 杆 菌(Agrobacterium)等多种内生细菌,另外还有一种新型菌株,定名为Leifsonia ginsengi。人参根部内生真菌有较丰富的多样性。Wu等[37]以林下山参作为研究对象,从其根部分离得到38株内生真菌,经鉴定将其归类为9个 属,分 别 是 丛 赤 壳 属(Nectria)、曲 霉 属(Aspergillus)、镰 刀 菌 属(Fusarium)、轮 枝 孢 属(Verticillium)、Engyodontium、Plectosphaerella、青霉属(Penicillium)、Cladosporium、子囊菌属(Ascomycete)。
2.1.2茎和叶内生菌的多样性
Vendan等[38]通过16S rDNA测序对韩国人参内生细菌的多样性进行分析研究,从人参茎内共得到51个菌落,属于7个不同属:Bacillussp.、Staphylococcussp.、Pseudomonassp.、Agrobacteriumsp.、Stenotrophomonas、Microbacterium、Paenibacillus。Hong等[39]将人参茎经组织裂解后涂布在LB琼脂平板上,通过16S rDNA测序鉴定出阿氏芽孢杆菌(Bacillus aryabhattai)、Acinetobacter guillouiae、巨 大 芽 孢 杆 菌(Bacillus megaterium)、Agrobacterium tumefaciens、Bacillus marisflavi、Bacillus safensis等。Khan等[29]进行山参的根、茎和叶片内生菌培养分离实验时,发现内生菌Bacteroidetes只存在叶片组织中,在根和茎中没有发现,表明一些菌株具有组织特异性。Gao等[40]从人参叶中分离得到一种内生细菌Paenibacillus polymyxa。
赵智灵等[41]对吉林省不同器官(根、茎、叶)的人参样品进行内生菌的分离纯化研究,发现了对锈腐病菌具有较强抑制效果的内生真菌齿孢青霉(Penicillium daleae)。孙畅[42]从人参叶片中分离获得18株人参内生真菌,归属于Penicillium、枝孢属(Gladaxporism)、Aspergillus、毛 壳 属(Chaetomium)和 枝 顶 孢 属(Acremonium)等8个属。以上研究说明了人参茎和叶片中内生菌的多样性和复杂性,部分内生菌可成为人参病害生防菌的新来源。
2.1.3种子和花内生菌的多样性
Hong等[39]从花梗中分离得到多种内生细菌,主要包 括Acinetobacter pittii、Bacillus amyloliquefaciens、Pseudomonas frederiksbergensis、腐生葡萄球菌(Staphylococcus saprophyticus)。孙月[43]从人参花中总共分离得到3株内生真菌,经形态学鉴定确定一株内生真菌茎点霉(Phomasp.)可能会产生人参皂苷Re。马伟等[32]发现人参种子中内生真菌占比最大的为子囊菌门(Ascomycota),优势菌属为Fusarium;人参种子中内生菌的分布以及丰度会由于个体不同而产生差异。综上,人参不同器官内生菌多样性存在差异。不同器官内生菌研究见表2。
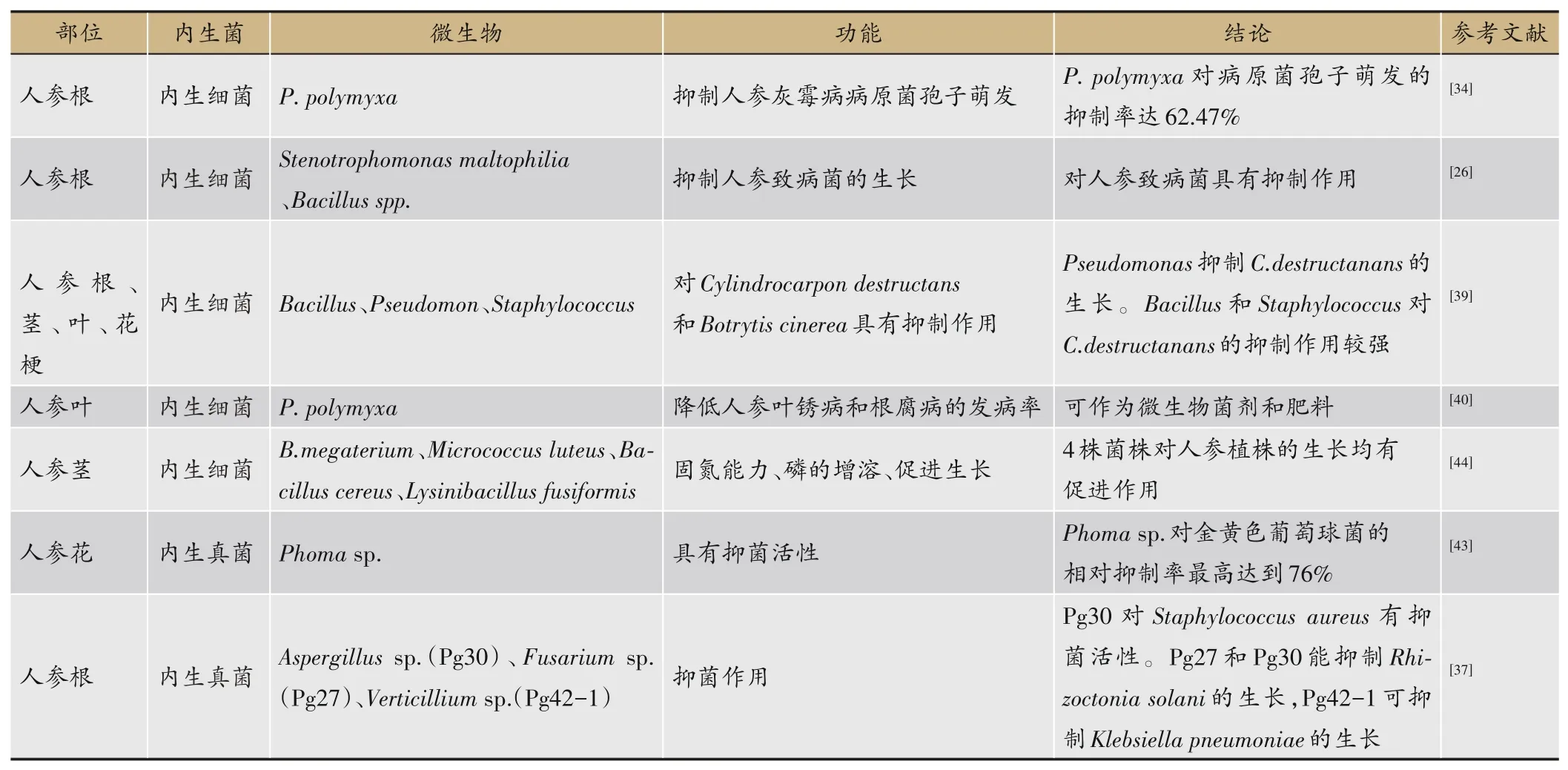
表2 人参不同器官内生菌研究
2.2 不同生长年限内生菌的多样性
人参在不同生长年限下,其内生细菌的多样性存在差异。Vendan等[38]发现1年生人参茎段内生细菌以Bacillus为 主,2年 生 的 菌 株 属 于Bacillus、Pseudomonas、Agrobacterium和Stenotrophomonas,3年生菌 株 属 于Bacillus、微 杆 菌 属(Microbacterium)、Agrobacterium和类芽孢杆菌属(Paenibacillus),成熟的4年生人参茎中主要含有葡萄球菌属(Staphylococcus),这与邱服斌[36]的研究结果相似,内生细菌多样性也会随着参龄增加呈下降趋势[45]。
人参内生真菌种类繁多,分布与人参植株年龄有关。Park等[30]研究发现Fusarium solani和Phoma radicina分别是1和2-4年生人参根中的优势内生真菌;Fusarium oxysporum和P.radicina在2-4年生人参中均有分布,在3年生人参根中未发现F.solani;桔绿木霉(Trichoderma citrinoviride)只出现在1年生人参中,Colletotrichum panacicola只出现在2年生人参中,锐顶镰刀菌(Fusarium acuminatum)只出现在3年生人参中。
2.3 不同栽培方式内生菌的多样性
根据栽培模式不同可以将人参划分为园参、林下山参和野生人参[45]。雷锋杰等[46]发现野生抚育人参根内生细菌的群落丰富度和多样性均高于农田栽参,栽培模式显著影响人参根内生细菌的多样性。这与张亚光等[45]的结论一致:栽培环境不同会引起人参内生细菌组成差异。人参的栽培方式不同,其内生真菌多样性也存在差异。徐丽莉[47]研究发现钩状木霉(Trichoderma hamatum)为园参内生真菌的优势种,鞭毛藻丛赤壳菌(Nectria haematococca)为移山参内生真菌的优势种。雷锋杰等[46]发现Ascomycota是3种栽培模式人参根中内生真菌相对丰度最高的类群,占各人参样品的80%以上。人参不同栽培方式内生菌多样性见表3。综上,栽培模式会影响人参内生菌群落丰富度、多样性以及优势物种。

表3 不同栽培方式人参内生菌的多样性
2.4 不同地理区域内生菌的多样性
不同地理区域的人参内生细菌多样性存在差异。Khan等[29]以韩国24个地区的山参为研究对象,发现每个门的相对丰度在不同采样点之间有很大的差异。Cho等[48]对韩国3个不同地区的人参进行研究,发现江华郡是唯一含有节杆菌属(Arthrobacter)、Rahnella和Microbacterium的采样点。锦山郡是唯一含有Paenibacillus和欧文氏菌属(Erwinia)的采样点。镇安郡是唯一含有泛菌属(Pantoea)和Pectobacterium的采样点;Pseudomonas和Bacillus是比较常见且容易通过培养得到的内生菌,这与李勇等[26]以及Vendan等[38]的研究一致。不同地理区域人参内生菌的多样性见表4。
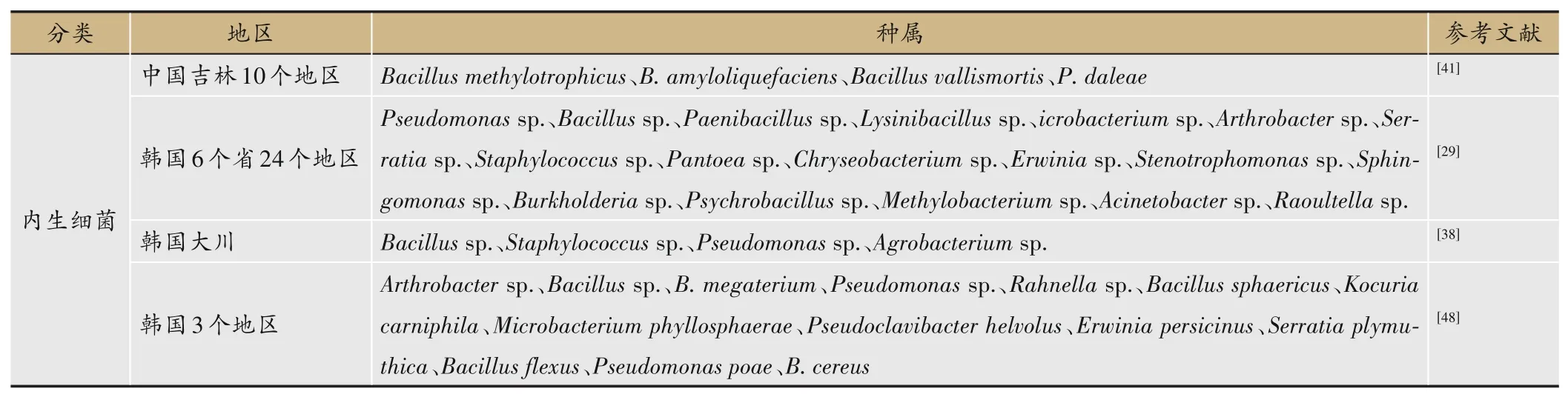
表4 不同地理区域内生菌的多样性
生长年限和栽培方式对人参内生菌多样性都具有显著影响[45,46];对可培养内生细菌进行研究发现,不同地理区域的细菌多样性水平差异很大;研究可培养内生细菌在不同组织中的分布和多样性,发现总物种丰富度在叶组织中最高,在茎中最低[29]。综上,不同地理区域、不同栽培模式、不同生长年限以及不同器官对内生菌的多样性均有影响。
3 人参内生菌及其次级代谢产物的活性
目前,已有文献表明植物内生菌对于植物中的有效活性成分或者植物的生长发育等方面都具有一定的促进或者抑制作用[49-50]。Carrion等[51]证明植物内生菌与NRPSs和PKSs编码的次生代谢物生物合成有关,能够较好地使植物免受外来病菌侵入,增强植物自身抵抗力。从内生菌中分离的萜类、黄酮类以及生物碱类等活性成分也具有抗癌、抑菌的作用[52],这将成为新的抗生素和抗癌剂的重要来源。
3.1 内生菌转化人参皂苷能力
内生菌具有将人参皂苷Rb1转化为人参皂苷Rd的能力。张薇[53]从人参根中分离得到2株内生真菌灰绿犁头霉和莫勒接霉,可将人参皂苷Rb1转化为人参皂苷Rd。陈雪英[54]研究发现内生细菌肠杆菌属(Enterobactersp.)可将人参皂苷Rb1转化为人参皂苷Rd。李学[55]发现人参内生细菌P.polymyxa可以促进其它皂苷转化为人参皂苷Rd。内生菌可将常见人参皂苷转化为稀有人参皂苷。人参内生细菌Burkholderiasp.将人参皂苷Rb1转化为人参皂苷Rg3,通过薄层色谱和高效液相色谱确定转化途径是Rb1→Rd→Rg3[44,56-57]。Fu[58]分离得到的人参内生细菌黄杆菌属(Flavobacteriumsp.)将人参皂苷Rb1转化为稀有人参皂苷Rg3,转化途径为Rb1→Gyp-XVII和Rb1→Rd→Rg3,为今后人参皂苷的制备提供了新思路。
Yan等[35]利用高效液相色谱法发现1株内生细菌Agrobacteriumsp.具有较高产稀有人参皂苷Rg3和Rh2的能力。Wu等[37]利用高效液相色谱法发现3株内生真菌具有产人参皂苷能力:在菌株Pg27(Fusariumsp.)与Pg30(Aspergillussp.)的次级代谢产物中检测出人参皂苷Rb2;在菌株Pg42-1(Verticilliumsp.)的次级代谢产物中检测出人参皂苷Rc。这为制药工业制备人参皂苷提供了新方法。人参内生菌及其次级代谢产物统计见表5。

表5 人参内生菌及其次级代谢产物统计
3.2 抗肿瘤活性
鞠鑫等[59]通过16S rDNA分子鉴定、蛋白质印迹法等手段从人参中分离到假单胞菌菌株G13476,并通过研究确定了此菌株对三阴性乳腺癌的调控机制,此结果为今后人参内生菌在抗肿瘤药物的开发研究提供了新的理论依据。徐丽莉等[27]将从人参根中分离得到的yuan-27、yuan-4、yuan-8、yuan-22等内生真菌进行稻瘟霉模型筛选,选择完全抑制稻瘟霉孢子萌发的菌株进行抗肿瘤活性筛选,经MTT法检测确定,发现有5株内生真菌抗肿瘤活性较好,其中yuan-27的抗肿瘤活性最优,为进一步开发抗肿瘤药物提供了新的研究对象。
3.3 抑菌活性
Kim等[60]发现人参根内生细菌Burkholderia stabilisEB159对人参病原菌具有明显的抑制作用,从该菌株的代谢产物中也分离出了抗菌活性成分,证实了人参内生细菌的抗菌活性;另外其在进行人参内生真菌抗人参根腐病化合物的全基因组序列分析的研究中也证实了EB159产生的代谢产物吡咯氮可作为潜在的抗菌剂。人参内生菌Trichoderma citrinoviride可以显著减轻B.cinerea和C.destructans引起的病害症状[61],人参内生菌Trichoderma polysporum对人参病原菌(Alternaria panax、R.solani等)所引起的疾病症状具有明显的抑制作用[62]。这些结果为今后植物内生菌在生物防治方面提供了理论依据。
3.4 其它活性
Kim等[63]从人参内生真菌Penicilliumsp.中分离提取纯化得到一种可下调酪氨酸酶和酪氨酸酶相关蛋白等黑色素生成基因表达的代谢产物,但对其酶活性并无影响;证实了该代谢产物具有抑制黑色素生成积累的活性,从而揭示了植物内生菌具有作为护肤化妆品成分的潜在利用价值。这些研究将为今后开发新的抑菌、抗癌药物甚至是化妆品的研发提供新的参考,为植物内生菌在农药、医药、化妆品行业的发展起到了推动作用。
4 人参内生菌对宿主的影响
植物内生菌与植物之间相互作用影响植物自身及其周围环境,从而影响植物自身的生长发育[64-65]。Pinski等[66]提出植物内生菌与植物之间的相互作用可增强植物对外界环境变化的耐受性及对病原菌入侵的抵抗能力,从而促进植物自身的生长,达到增产的目的。Giauque等[67]发现植物中的内生菌特性与植物的生活环境以及其胁迫耐受性和资源利用有关。这些研究都为植物内生菌与植物自身之间的相互作用研究提供了新的参考依据。
4.1 增强人参对外界环境抵抗力并促进人参生长
研究发现P.polymyxa可有效降解人参上5种残留农药[68],还具有降低人参叶锈病和根腐病发病率的能力[40],表明人参内生菌可以提高人参对外界环境的抵抗力,促进人参的健康生长[69]。Hong等[25]对2-6年生的韩国人参进行研究:发现相较其他年限的人参,在3年生的人参根中铁载体和生长素相关性状最为突出,并且在3年生人参中检测到6个促进植物生长基因的表达,表明内生细菌与3年生人参的相互作用有利于根系生长。以上研究为今后植物生物菌肥的研究奠定了良好的基础。
4.2 促进新的化合物合成
已有研究表明:从桔梗中分离得到的内生菌具有促进新的化合物合成的能力,CGMCC6882菌株能够在人参皂苷及其内生共生微生物的共同作用下生成多糖,但会受到植物内生共生真菌及植物本身产生的皂苷或者其他活性成分的影响[70]。人参也具有类似的功能,Bao等[71]以人参内生真菌F.oxysporum为原料采用X射线、透射电镜等方法及技术手段进行了纳米银粒子的合成,合成的新材料在室温下非常稳定,此研究为今后以植物内生菌为原料合成新的合成材料提供了依据,使今后采用新方法大规模合成生产纳米银材料成为可能。
4.3 提升人参产量与品质
药用植物内生菌具有生物防治、促进生长的特性,影响药用植物的次生代谢、生物合成、产量和品质,合理地开发和利用内生菌资源将有利于药用植物种植业向安全、高品质方向发展[7,72]。在栽培条件下,人参会受到锈腐病、根腐病等病害的影响,而内生菌对于人参病害病原菌具有抑制作用[41]。人参内生菌P.polymyxa处理的1、2、3和4年生人参植株的发病率均低于同龄对照,处理组的平均重量分别比对照组提高31.64%、58.87%、46.70%和18.61%[40],表明内生菌参与人参病害的生物防治,提高人参的产量。Gao等[40]通过田间试验研究发现,人参内生细菌P.polymyxa处理的1、2、3和4年生人参植株的平均高度均高于同龄对照组,且总皂苷含量分别比对照组提升36.83%、44.52%、67.96%和79.44%,4年生人参植株中人参皂苷Rd的含量比对照组高308.01%;表明从人参中分离到的P.polymyxa是一种有益内生细菌,可能提升人参的品质。1-氨基环丙烷-1-羧酸脱氨酶(ACC脱氨酶)在重金属污染及盐、干旱胁迫等逆境条件下,能显著提高农作物的抗逆性和产量,目前已有研究从人参中分离得到产ACC脱氨酶的内生细菌[69],其对非生物胁迫下人参的促生作用、产量与品质的影响还需进一步研究。
然而,在某些情况下内生菌也会对宿主植物产生一些不利的影响。Faeth等[73]发现在营养缺乏的土地上,未感染内生真菌的亚利桑那羊茅长势好于感染内生真菌的羊茅。陈佳昕等[74]将从茅苍术中筛选到的15株内生真菌回接至茅苍术组培苗,结果发现有3株内生真菌致使组培苗死亡,推测在茅苍术内生菌中存在部分有害内生菌类型。目前未见人参内生菌负面影响的文献报道。
5 小结与展望
目前,人参内生菌的研究仍处于起步阶段,研究过的人参内生菌数量仅为其巨大资源库的很少一部分,且多限于内生菌对人参生长发育、有效成分、抑菌性等表观研究,对其具体作用机制仍不明确。人参内生菌采用的研究方法包括传统的分离鉴定以及高通量测序技术,涉及多样性、生物转化、抑菌活性等多个方面,暂未发现其对宿主具有负面影响。利用现代技术手段,对人参内生菌进行进一步的系统研究亟待开展。目前关于人参内生菌的研究还存在下述问题:①虽然同时采用多种培养基/培养条件进行内生菌的分离,但工作量巨大;②人参内生菌与宿主人参的相互作用及作用机制尚不明确,关于分子机制方面的研究较少;③目前关于人参内生菌的研究依然停留在分离、发酵、抑菌实验,主要集中在实验部分,关于人参内生菌的实际应用效果尚未深入研究;④人参内生菌宏基因组测序进展缓慢,如何分离人参内生菌基因组和排除植物基因组数据是研究内生菌-宿主植物相互作用关系的一大阻碍;人参内生菌在人参中增殖和定殖、促生、缓解胁迫方面的研究还未全面开展;在生物和非生物胁迫下,人参内生菌在宿主基因表达、代谢过程以及表型性状调控中的作用有待进一步研究。
对于人参内生菌未来的研究热点或将集中在以下几点:①关于人参内生菌与人参之间的系统进化关系目前尚未有定论出现,这是值得深入研究的方向;②目前人参内生菌与人参之间的相互作用尚未完全阐明,人参内生菌对人参的基因表达、代谢以及其他生理方面的影响等都是值得深入探索的;③人参内生菌中的次级代谢产物及其合成机制等都有待深入研究探索,而对产人参皂苷的人参内生菌菌株,应当加强对其工作机理的研究,进而培育高产内生菌菌株;④宏基因组测序功能基因很难和物种建立一一对应的关系,人参内生菌参考基因集的建立,以及宏基因组学结合培养组学在人参内生菌研究中的应用也将是未来的热点研究方向。
