不同地理种群棘胸蛙的表征与环境适应
2022-12-28蒋泽元卢俊义张丹丹黄思琪宋静翁凌云郑荣泉田苇
蒋泽元,卢俊义,张丹丹,黄思琪,宋静,翁凌云,郑荣泉,3*,田苇
(1.浙江师范大学 化学与生命科学学院,浙江 金华 321004; 2.浙江省野生动物生物技术与保护利用重点实验室,浙江 金华 321004;3.浙江师范大学 行知学院,浙江 金华 321100; 4.贵州黔东南桥水农业科技开发有限责任公司,贵州 黔东南 556000)
棘胸蛙俗称石蛙、石鸡等,隶属于两栖纲,无尾目,叉舌蛙科,棘胸蛙属,主要分布在我国的云贵、华南、华东等长江以南地区[1]。
由于棘胸蛙是我国叉舌蛙科物种中资源利用最广泛的物种之一,同时兼具食用和药用价值,因此,棘胸蛙遗传特征及生长发育条件的相关研究已经成为学者们关注的热点。利用线粒体及微卫星基因标记对棘胸蛙13个地理种群进行了分析,结果发现,野生棘胸蛙可分为4个不同的进化支系,并认为棘胸蛙可能是一种未发现形态变异的物种复合体[2-4]。在宏观特征上,不同地理种群棘胸蛙的鸣声[5]、热偏好性[6]、形态大小[7-9]都存在着显著差异,且不同地理种群的棘胸蛙在热偏好性以及形态大小上与纬度相关[5-6]。本研究旨在分析不同支系棘胸蛙的环境数据与其形态指标之间的关系,以期寻找棘胸蛙支系分化与温度适应方面的联系,同时也为棘胸蛙规模化养殖和品种选育提供依据。
1 材料与方法
1.1 材料与数据采集
2019年6—9月份在韶关、南昌、鹰潭、池州、黄山、龙岩、德化、遵义共8个样地采集二龄性成熟雄性棘胸蛙各15只。采集样品的环境一般为静水的池塘或溪流的缓流处,且均为健康活泼成体。采集的棘胸蛙在实验室条件下驯养待用。
依据费梁等[10]和Watters等[11]的测量标准,利用电子数显游标卡尺(精度0.01 mm)测定了所有样本的16个形态特征性状。所有形态参数测量所用样本均为活体,形态学指标均参考《中国动物志》[12]。
1.2 环境因子数据收集
结合中国气象数据网以及样地野外温湿度记录仪的数据,对8个样地的12个环境因子进行收集。通过多维变量的降维分析,推断出影响棘胸蛙适应性进化的主导因子。
1.3 不同地理种群棘胸蛙形态分化
为消除样本个体年龄差异对贡献值分析结果的影响,本研究中采用棘胸蛙各形态指标数据与体长的比值为变量进行主成分分析,结果共得到15个特征变量,提取变量的主成分,并计算各主成分的特征值和贡献率,最后基于主成分得分值做散点图。
通过逐步法对8个地理种群棘胸蛙的形态数据进行判别式分析,从而探讨棘胸蛙不同地理种群间形态差异情况。逐步判别删除或保留变量是由变量的F值决定,当F值大于Entry默认值3.84时,该变量就会进入模型;当变量的F值小于默认的Removal值2.71时,该变量就会从模型中剔除。利用SPSS 21.0计算所测量形态数据的特征值、典型相关系数、Wilks’λ和非标准化系数,并根据判别系数计算各种群棘胸蛙的判别函数,最后利用判别函数的得分值绘制散点图。样本的归属判别则是通过马氏距离判断,马氏距离指每个样本在图中的位置到每个分组组心的距离,样本到哪个分组组心的马氏距离最短,该样本则被判为该组。利用交互检验方法检验判别函数的正确率。
2 结果与分析
2.1 形态学指标
通过对8个地理种群的形态学数据进行统计,描述性统计结果以平均数±标准差(Mean±SD)形式标出,结果见表1。
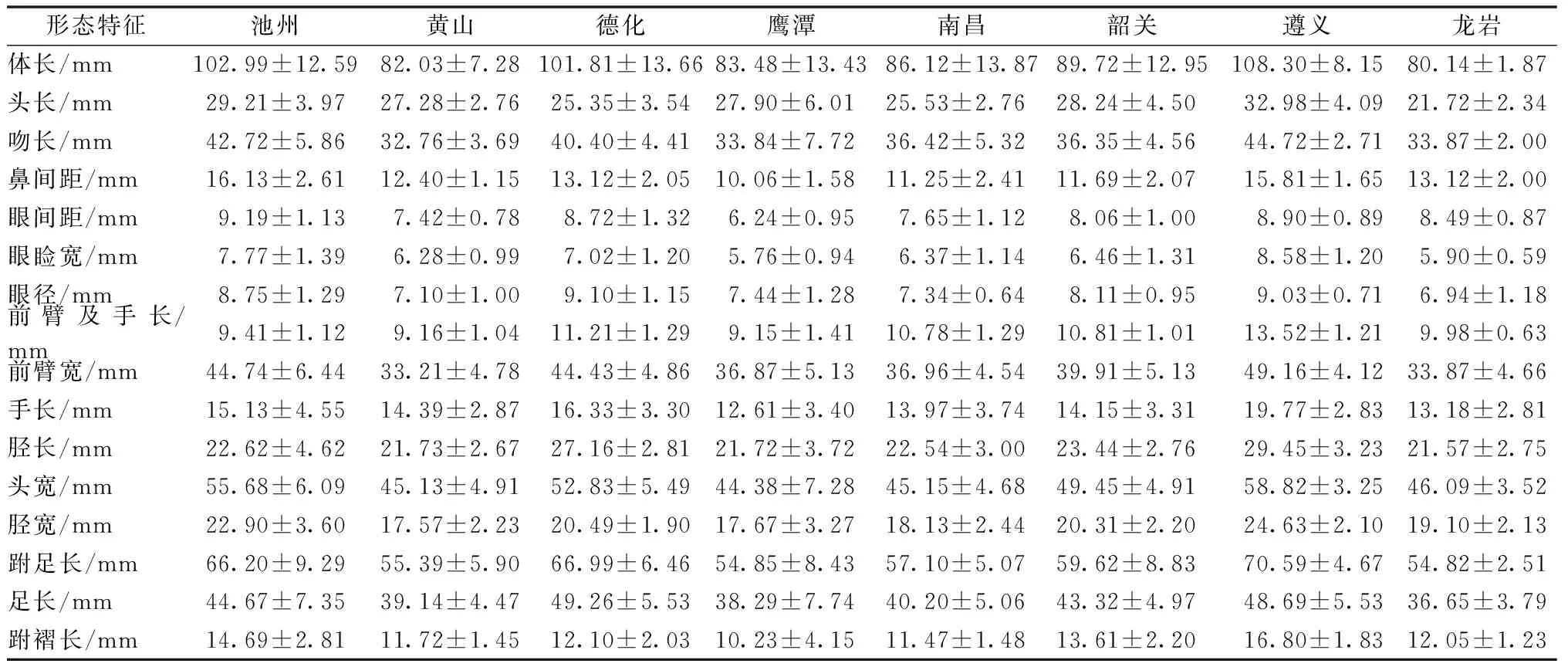
表1 不同地理种群棘胸蛙形态特征的描述性统计值
2.2 主成分分析
对15个形态指标量度与体长的比值进行主成分分析,分析显示,数据的KMO值为0.926,Barttlett’s球形检验拒绝零假设,变量之间存在相关性,可以进行主成分分析。前3个主成分的特征根分别为7.467、1.241和1.02,贡献率分别为49.781%、8.271%和6.802%。3个主成分的累积贡献率为64.854%,涵盖了全部数据的大部分信息,所以,我们只对前3个主成分进行分析。3个主成分因子负荷值见表2。依据所得的第一主成分和第二主成分得分值绘制PCA散点图(图1中A),结果显示,不同地理种群的棘胸蛙在散点图上明显可分为2个组。而黄山、德化、鹰潭等种群样本差异不大。在PC2轴上,广东韶关的棘胸蛙样本特征指标值范围大于其他地理种群,说明在吻长/体长、鼻间距/体长、眼间距/体长和眼睑宽/体长等与头部横向形态量度相关的特征上,广东韶关样本多样性较高。
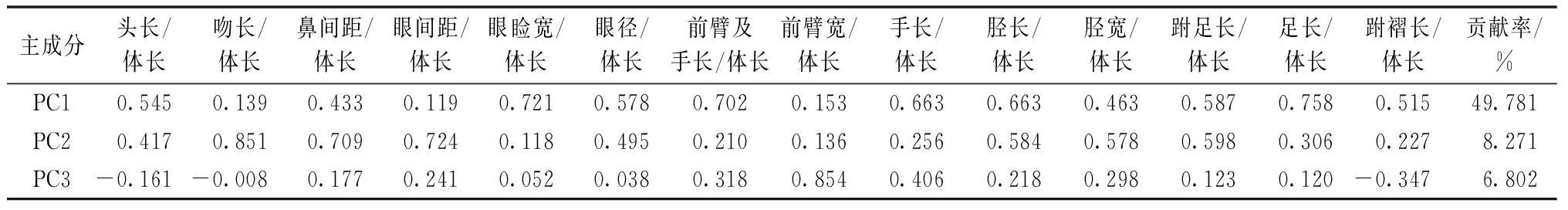
表2 棘胸蛙外部可量度性状主成分分析的因子负荷值

A—不同地区棘胸蛙外形数据聚类分析;DH—德化;NC—南昌;YT—鹰潭;SG—韶关;LY—龙岩;HS—黄山;CZ—池州;ZY—遵义;B—不同地区棘胸蛙外形数据判别分析散点。图1 不同地区棘胸蛙分子形态与身体形态分析
2.3 判别式分析
对亲缘关系较远的物种,传统意义上的测量及计算就能得到有效的鉴别特征描述,然而对于表型特征相近或者相似的物种,传统的分类方法无法鉴定分类。聚类分析可以量化相似个体的细微差异,因此,能够用来作物种初步鉴别归类的依据,因而该方法在相似程度较大的物种辨别分析中应用广泛。本研究中对15个样本形态指标与体长的比值进行聚类分析(图1中A),显示各地区间无明显差异。进一步对15个形态指标量度与体长的比值进行对数变换的逐步判别分析,其中头长/体长,鼻间距/体长,眼径/体长,胫宽/体长,吻长/体长,手长/体长和跗褶长/体长7个指标对判别贡献率较高,被选择进入函数。使用7个性状的特征值建立了不同地理种群棘胸蛙的判别函数。其中,函数式中的X1至X7分别代表上述7个判别贡献率高的性状。8个地理种群的判别公式如下:
安徽池州地理种群:Y=-52.83+48.62X1+63.53X2+117.03X3-60.80X4+56.57X5+195.97X6+
111.42X7;
安徽黄山地理种群:Y=-58.43+82.80X1+18.86X2+79.93X3+66.01X4+122.35X5+103.07X6+
88.19X7;
福建德化地理种群:Y=-51.20+23.91X1-18.78X2+90.12X3+129.19X4+157.17X5+113.07X6+
57.05X7;
江西鹰谭地理种群:Y=-55.61+93.26X1-36.12X2-29.96X3+125.12X4+132.35X5+130.72X6+
52.45X7;
江西南昌地理种群:Y=-57.96+59.20X1-55.82X2+102.00X3+229.08X4+113.72X5+109.29X6+
66.97X7;
福建龙岩地理种群:Y=-63.21+14.27X1+11.32X2+192.80X3+120.05X4+98.33X5+166.24X6+
97.16X7;
广东韶关地理种群:Y=-61.74+66.73X1-53.26X2+101.01X3+158.56X4+99.42X5+153.76X6+
94.27X7;
贵州遵义地理种群:Y=-61.90+49.47X1+12.10X2-59.65X3+174.65X4+126.10X5+152.69X6+
103.64X7。
依据判别函数的得分值分别作为X轴和Y轴,绘制不同地理种群棘胸蛙外形数据判别分析的散点图(图1中B)。依据各个样本中心到各组中心的马氏距离长短来判别样本组别,样本到哪组组心的马氏距离最短,该样本则被判为该组。判别分析散点图显示,8个不同地理种群的棘胸蛙样本被区分为三组,通过主成分分析对8个地理种群棘胸蛙进行了形态上相似性和差异性的分析和比对,在判别分析散点图中(图1中B),遵义地区一组,德化、龙岩、鹰潭、韶关、南昌、黄山地区一组,池州地区一组。结果说明,这三组地理种群的棘胸蛙在形态学度量数据上存在较大差异。
2.4 环境因子与形态特征
6种主要环境因子分别与棘胸蛙各形态特征相关性分析的结果表明,手长与体长的比值与温度因子呈显著正相关,与年均降水因子、气压因子、纬度因子呈显著负相关;眼径与体长的比值与温度因子呈显著正相关,与年均降水因子、纬度因子呈显著负相关;跗褶长与体长的比值与经度因子呈极显著负相关(表3)。
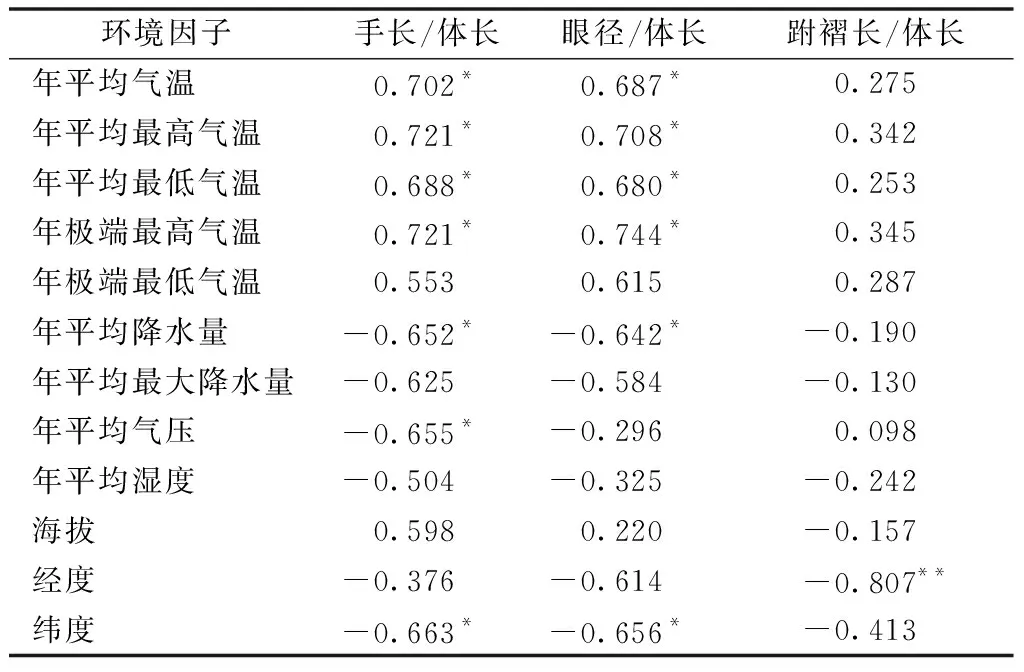
表3 环境因子与形态特征的关系
3 小结与讨论
形态学方法是最早进行物种分类的基本方法之一,在两栖动物的物种分化及新种界定研究中应用广泛,例如早在1991年,魏刚等[13]就通过胫长与体长的比例,头长与头宽等形态特征的比较,将不同地理种群的中国林蛙进行区分及定种。费梁等[14]在中国西北地区中国林蛙各居群的分类学研
究中发现,中国林蛙与高原林蛙后肢与体长之比存在显著性差异,因此,将该特征作为高原林蛙的鉴别特征之一。费梁等[15]对分布于广西龙胜和湖南宜章的龙胜臭蛙作了对比分析,发现分布于湖南宜章的龙胜臭蛙前臂宽与体长之比与龙胜种群产生了显著性差异,因此,将其命名为宜章臭蛙。本研究中,不同地理种群棘胸蛙形态特征存在显著的分化,其中安徽池州种群棘胸蛙的形态特征最为清晰稳定,江西南昌、福建龙岩可能处于种群形态分化的过渡带,安徽池州与其他地理种群的棘胸蛙形态分化最为明显,尤其是与贵州遵义种群形态差异显著,已达到种间可区分的水平。
生态过渡带中栖息地的多样性形成了各种各样的生物形式,然后形成了带有可以遗传的多种生物物种。环境梯度变化产生形态变异、交配方式变异,生殖隔离通常经历生态过渡带的演化并保持[16]。不同栖息地选择比地理隔离更能引起适应性形态分化。依赖栖息地的选择影响一些具有生态意义的特征,形成形态和行为分化,这些分化特征促进了相应的交配,从而建立沿着环境梯度的遗传分化的机制[17]。近年来随着全球气候的变化,两栖类的物种或种群数量受到严重威胁,了解环境因素对群落成员的影响是确定物种在空间中分布方式的重要部分[18]。从环境因素的角度解释物种的地理布局模式,可以对物种在其分布区域的适应机制提供见解,同时通过整合环境和生态变量,可以更好地解释物种分布格局[19]。例如,Amador等[20]研究了空间和环境因素对南美洲两栖动物物种分布格局的影响,结果发现,降水和海拔对保持当地两栖动物物种多样性起着关键的作用。在中国林蛙中发现,体型及四肢随着纬度的增加、温度的降低而逐渐增长[21]。生活于温度较低的高海拔红瘰疣螈,会增长四肢及头部以获取更多的食物[22]。在本研究中探究了不同支系棘胸蛙形态特征与环境因子间的相关关系,结果显示,与个体大小相关的形态特征与当地环境温度呈现明显负相关,即生活于温度更高的较低纬度棘胸蛙显示出前肢更长,眼径更大,从而其整体体形偏大的趋势,该趋势符合“初级生产力假说”。同时棘胸蛙作为叉舌蛙科资源利用最广泛的类群之一,早期对其生物学特征地理变异的研究已较为普遍,比如,体形大小、鸣声、性二态性、热生理学特征等,本文中仅探讨了棘胸蛙形态特征与环境因子间的相关关系,对于阐述其适应性进化的问题研究还不够全面,因此,对于棘胸蛙适应性分化及其影响因子的研究需要进一步增加综合特征指标及环境因子种类并使用更科学的分析方法。
