七彩神仙鱼C型凝集素家族成员CD302和MBL的克隆及在病原刺激下的免疫应答
2022-12-20马腾飞高建忠陈再忠
马腾飞,温 彬, ,刘 鑫,缪 琳,高建忠, ,陈再忠,
1. 上海海洋大学/水产种质资源发掘与利用教育部重点实验室/农业农村部淡水水产种质资源重点实验室,上海 201306
2. 上海海洋大学/水产科学国家级实验教学示范中心,上海 201306
七彩神仙鱼 (Symphysodon aequifasciatus) 隶属于鲈形目、慈鲷科、盘丽鱼属,原产于南美洲的亚马逊河流域,是观赏鱼中的名贵品种[1-3]。该物种具有独特的亲代抚育行为,即仔鱼在孵化后的一个月内以亲本体表分泌的黏液为食[4-5]。尽管受到子代长达1个月的咬食刺激,但亲鱼体表并未被感染和病害,暗示亲代体表可能具有较强的免疫防御。Chong等[6]研究发现,为了适应子代免疫需要,C型凝集素 (C-Type lectins, CTLs) 在育幼亲本体表黏液中特异表达,可能在亲子抗菌防御方面发挥重要作用。
CTLs作为一种重要的模式识别受体,对鱼类抵御病原微生物入侵有着重要的作用。CTLs通常含有一个或多个碳水化合物识别结构域 (Carbonhydrate-recognition domain, CRD) 或称C型凝集素结构域 (C-Type lectin domain, CTLD)[7-8]。CTLs研究在脊椎动物中较为广泛深入,目前已有1 000多个CTLs被鉴定出[9]。鱼类CTLs研究主要集中于其对病原相关分子模式的识别与结合、引发致病微生物的凝集、增强巨噬细胞对致病菌的吞噬能力以及激活补体系统等方面[10-11]。CD302 (Cluster of differentiation 302),又称DCL-1 (DEC-205-associated C-type lectin-1),被划分在CTLs家族XV组 (Bimlec),属于I型跨膜蛋白[12-13];CD302作为一种凝集素受体,参与免疫细胞的功能调节。Chen等[14]发现,香鱼 (Plecoglossus altivelis) 肝、头肾和脾中PaCD302在鳗弧菌 (Vibrio anguillarum) 感染下表达显著上调,且可能通过调节单核细胞或巨噬细胞来参与对细菌感染的免疫反应。甘露糖结合凝集素 (Mannan-binding lectins, MBL) 广泛存在于动物中,是第一个被发现具有防御功能的CTLs[15];它可以特异性识别细菌、真菌和病毒等病原体,还可以与微生物表面的糖类结合,起到免疫调节以及抵御外界病原微生物入侵的作用[16-17]。笔者前期对七彩神仙鱼育幼期皮肤转录组的分析发现,CD302在亲代抚育期间持续高表达,而MBL基因在育幼结束后表达显著上调,暗示着二者可能参与了亲子的先天免疫反应,且扮演不同角色。
气单胞菌属 (Aeromonas) 是一种在观赏鱼类中引起疾病的常见细菌[18-19],也是造成七彩神仙鱼发病的主要病原之一[20]。El-Ghany等[21]在患病的七彩神仙鱼中分离出嗜水气单胞菌 (A. hydrophila)。本研究从七彩神仙鱼皮肤中克隆获得2个新的CTLs家族成员,命名为SaCD302和SaMBL,并通过荧光定量PCR (Real-time quantitative PCR, qRTPCR) 分析它们在不同组织中的表达,以及在嗜水气单胞菌刺激下相关组织中的表达变化,以期了解CD302和MBL在七彩神仙鱼免疫系统中可能的作用,丰富鱼类CTLs功能研究基础。
1 材料与方法
1.1 实验材料与试剂
健康的七彩神仙鱼 (体长10~13 cm,体质量80~100 g) 取自上海海洋大学滨海养殖基地,于(28±0.5) ℃,pH 7.8±0.5,持续充氧条件下暂养,使其适应养殖环境。嗜水气单胞菌菌株来自上海海洋大学病原库。
TRIzol (Invitrogen,美国)、PrimeScript™ RT Master Mix (TaKaRa,日本)购自上海生工生物公司。2×T5 Fast qPCR Mix (TsingKe,中国)、金牌Mix green (TsingKe,中国) 购自北京擎科生物公司。所有引物运用Primer Premier 5.0软件设计后,由上海生工生物工程技术服务有限公司合成。
1.2 总RNA的提取和cDNA的合成
选取3条雄性七彩神仙鱼进行解剖,取脑、鳃、心脏、头肾、脾脏、皮肤、肝脏、肠道、肌肉和性腺共10种组织,每个组织3个重复。使用TRIzol试剂提取各组织中的RNA。经质量分数为1% MOPs琼脂糖凝胶电泳与微量核酸蛋白测定仪进行定性和定量检测。根据PrimeScript™ RT Master Mix 说明书,以1 μg总RNA为模板反转录成cDNA 后,−20 ℃ 保存。
1.3 基因克隆
从前期测定七彩神仙鱼皮肤转录组数据库中选取SaCD302和SaMBL的CDS区来分别设计引物(表1)。以七彩神仙鱼皮肤组织的cDNA为模版,利用PCR及上述引物对SaCD302和SaMBL进行扩增。扩增体系25 μL:皮肤cDNA 1 μL,正反向引物各 1 μL,金牌 Mix green (TsingKe) 22 μL。反应条件:98 ℃预变性2 min;98 ℃变性10 s,56 ℃退火10 s,72 ℃延伸10 s,34个循环;72 ℃总延伸1 min;4 ℃保温。反应结束后,将产物置于10 g·L−1琼脂糖凝胶电泳中进行检测,使用凝胶成像系统观察电泳结果,目的片段回收送上海生工生物公司测序。

表1 七彩神仙鱼 SaCD302 和 SaMBL 基因克隆及荧光定量PCR引物Table 1 Cloning and fluorescence quantitative primers of SaCD302 and SaMBL gene in S. aequifasciatus
1.4 生物信息学分析
使用NCBI上的ORF Finder查找SaCD302和SaMBL基因的开放阅读框,并推导出基因的氨基酸序列。使用BLAST (http://www.ncbi.nlm.nih.gov/BLAST/) 进行序列同源性分析;采用ExPASy在线工具 (http://web.expasy.org/compute_pi/) 预测等电点及蛋白分子量,利用SMART (http://smart.emblheidelberg.de/)进行结构域预测;使用SWISSMODEL (https://www.swissmodel.expasy.org/) 进行蛋白三级结构预测;利用Clustal X和MEGA 5.0软件进行多序列比对和聚类分析,采用Neighbor-Joining方法构建系统发育树。
1.5 组织表达分析
根据克隆验证得到的SaCD302和SaMBL序列来分别设计qRT-PCR的特异性引物(表1)。采用前期验证的内参基因β-actin来设计引物β-actinqPCR-F和β-actin-qPCR-R[22]。以上述10个组织的cDNA为模版,利用qRT-PCR分别分析SaCD302和SaMBL在各组织中的表达情况。反应体系 20 μL:2×T5 Fast qPCR Mix (TsingKe) 10 μL,cDNA 模板 1.6 μL,正反向引物各 0.8 μL,ddH2O 6.8 μL。反应条件:95 ℃ 1 min (预变性,1个循环);95° C 10 s,56 ℃ 5 s,72 ℃ 10 s (扩增段,共40 个循环);95 ℃ 5 s,65 ℃ 5 s,95 ℃ 15 s (熔解段,1个循环)。每个样品重复检测3次,包括目的基因 (SaCD302和SaMBL) 和内参基因 (βactin)。采用 2−ΔΔCt计算 SaCD302 和 SaMBL 在各组织的相对表达量。
1.6 嗜水气单胞菌免疫刺激后的表达变化
实验组随机选取36尾健康的七彩神仙鱼,养殖于6个水族箱,使用1×107CFU·mL−1嗜水气单胞菌菌液对其进行腹腔注射 (200 μL·尾−1),对照组于另外36尾鱼中注射等量的PBS缓冲液。两组分别在注射后第0、第6、第12、第24、第48和第72小时取样,每个时间点实验组与对照组均随机选取6尾存活的七彩神仙鱼,提取皮肤、头肾、肝脏和肠道4个组织的总RNA后,反转录为cDNA,通过qRT-PCR检测嗜水气单胞菌刺激SaCD302和SaMBL的应答模式,反应条件和体系同上。使用2−∆∆Ct(∆∆Ct =∆Ct实验−∆Ct对照)法计算所得数据,分析SaCD302和SaMBL的相对表达量,并使用SPSS 20.0软件进行单因素方差分析。
2 结果
2.1 基因克隆及生物信息学分析
SaCD302和SaMBL的cDNA序列ORF区序列长度分别为741和795 bp,分别编码246和264个氨基酸 (图1-a、1-b)。蛋白序列预测蛋白分子量(MW) 分别为27 614.17和27 912.51 D,理论等电点 (PI) 分别为4.89和5.97。SaCD302结构域预测结果显示,在N端29个氨基酸 (Amino acid, aa) 残基为信号肽序列 (aa 1—29),随后是长度为138个氨基酸的CTL结构域 (aa 32—169),C末端有跨膜区共23个氨基酸 (aa 184—206,图2-a)。SaMBL结构域预测显示,其含有2个低复杂度区域 (aa 32—92和aa 104—121) 和1个典型的CTL结构域(aa 137—263,图 2-b)。

图1 SaCD302 (a) 和SaMBL (b)基因的核酸序列及氨基酸序列Fig. 1 Nucleotide and deduced amino acid sequence of SaCD302 (a) and SaMBL (b)
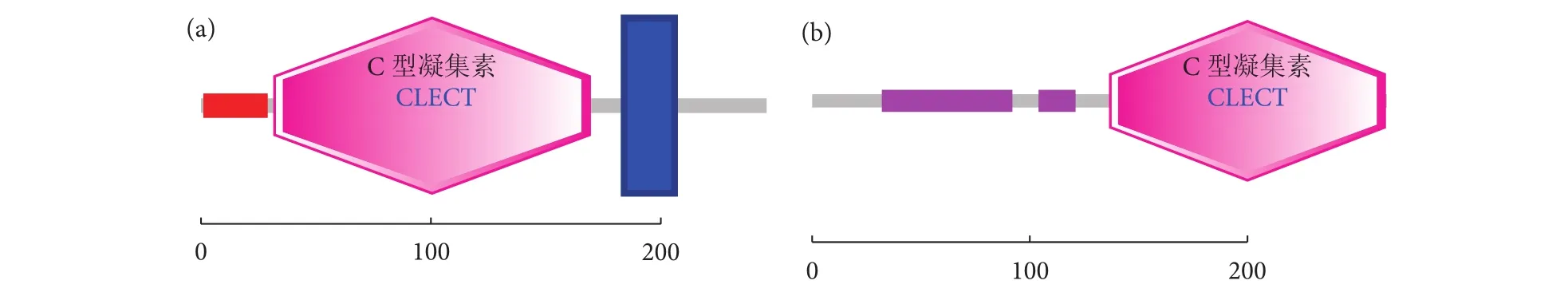
图2 SaCD302 (a) 和SaMBL (b) 结构域示意图Fig. 2 Domain of SaCD302 (a) and SaMBL (b)
2.2 多重序列对比与进化分析
BLAST同源性对比结果显示,SaCD302 与10种鱼的CD302氨基酸序列具有较高的同源性(51.93%~94.72%)。与尼加拉瓜湖始丽鱼 (Archocentrus centrarchus) 同源性最高 (94.72%),其次与斑马拟丽鱼 (Maylandia zebra) CD302同源性较高(82.93%),与斑马鱼 (Danio rerio) 的同源性最低(51.93%,图3-a)。SaMBL与8种鱼的MBL氨基酸序列的同源性介于28.64%~90.73%,与尼加拉瓜湖始丽鱼同源性最高 (90.73%),与斑马鱼的同源性最低 (28.64%,图3-b)。使用MEGA软件对各物种CD302和MBL氨基酸序列进行系统发育树构建,结果显示,所有鱼类的CD302聚为一簇,其他哺乳动物聚为一簇 (图4-a);斑马鱼的MBL与其他哺乳动物聚为一簇,其余鱼类聚为一簇 (图4-b);SaCD302和SaMBL均与同为慈鲷科的尼加拉瓜湖始丽鱼关系最近,符合自然物种的进化规律。

图3 SaCD302 (a) 和SaMBL (b) 氨基酸的多重序列比对Fig. 3 Multiple sequence alignment of amino acid sequences of SaCD302 (a) and SaMBL (b)
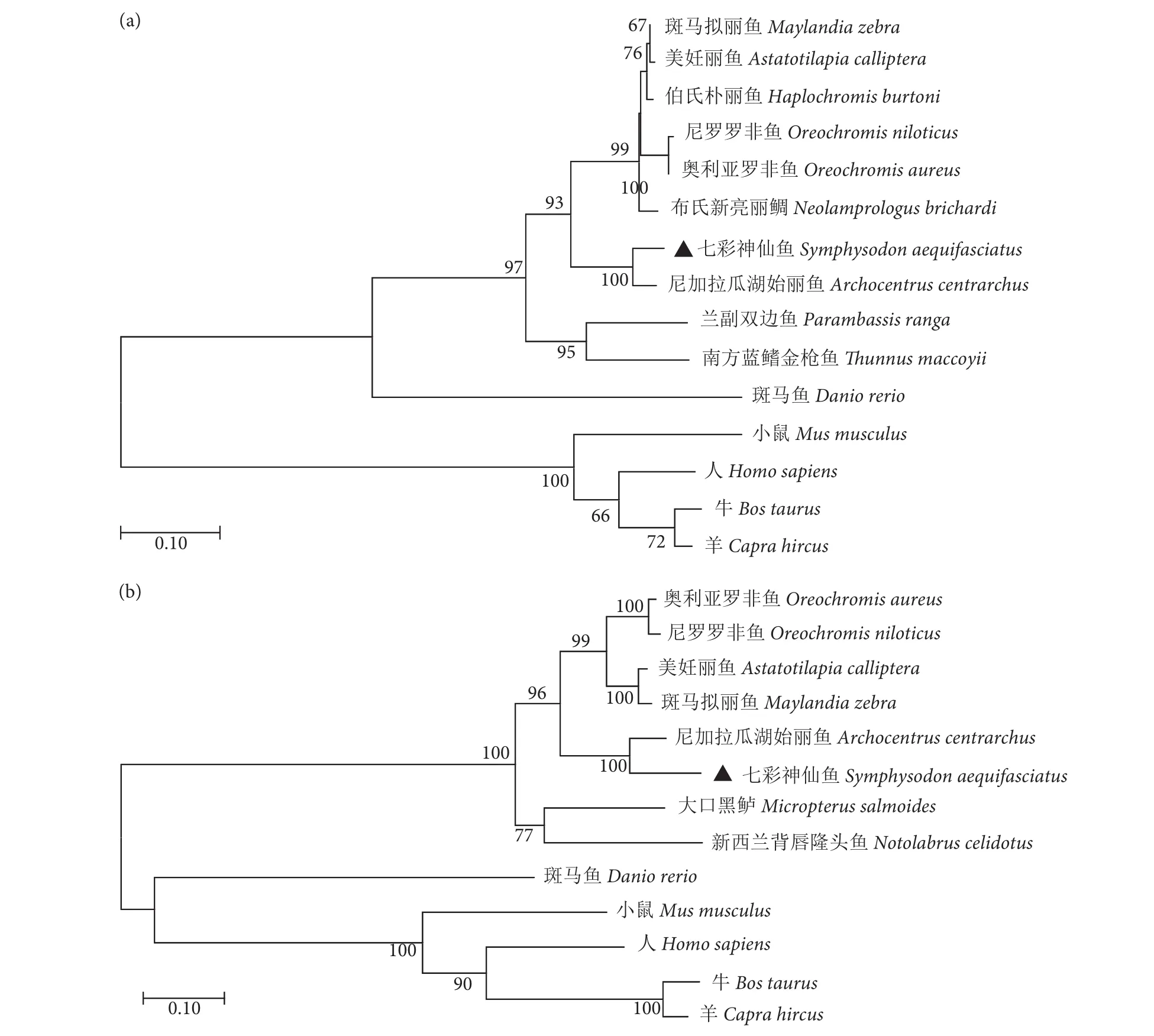
图4 基于NJ法构建的SaCD302 (a) 和SaMBL (b) 氨基酸序列系统进化树Fig. 4 Neighbor-joining phylogenetic analysis of SaCD302 (a) and SaMBL (b)
2.3 组织表达分析
通过qRT-PCR对SaCD302基因在不同组织中表达水平进行分析。结果显示,SaCD302广泛表达于七彩神仙鱼各组织中,但存在一定的组织差异性。其中在头肾中表达量最高,而后依次为肝脏、鳃、脑、脾脏等,在肌肉中表达量最低 (图5-a)。SaMBL在皮肤中显著表达,而在肠道、肝脏、脾脏和性腺中表达量却非常低 (图5-b)。

图5 SaCD302 (a) 和SaMBL (b) 在七彩神仙鱼不同组织中的表达特征注:图中不同字母表示在不同组织中存在差异性显著 (P<0.05);后图同此。Fig. 5 Relative expression of SaCD302 (a) and SaMBL (b) in different tissues of S. aequifasciatusNote: Different letters indicate significant difference in expression of different tissues (P<0.05). The same case in following figures.
2.4 病原刺激的表达变化
利用qPCR方法分析嗜水气单胞菌刺激后不同时间七彩神仙鱼各组织中SaCD302和SaMBL的表达变化特征。结果表明SaCD302 mRNA在脾脏中的表达在第6小时增加并达到顶峰,为对照组的2倍;在第12小时开始降低,第24—第72小时有轻微起伏,最终降低到正常水平 (图6-a)。在头肾中的表达第6小时开始增加,在第12小时达到顶峰,分别为对照组的3.6和4.1倍;在第24小时开始降低,在第48和第72 小时回升,分别为对照组的1.3、3.0和2.2倍 (图6-b)。SaCD302 mRNA在肠道中的表达量第6小时开始增加,为对照组的1.8倍;第12小时达到峰值,显著高于对照组,为对照组的6.4倍;第24小时出现降低,第48小时达到最低值,为对照组的0.03倍,第72小时回调至正常水平 (图6-c)。在皮肤中的表达量第6小时开始增加,第12小时达到顶峰,分别为对照组的3.7和8.0倍;第24小时开始下降,在第48小时降到最低,为对照组的0.15倍;第72小时回调至正常水平 (图6-d)。
SaMBL在感染后脾脏中的表达量均低于对照组(图7-a)。在头肾中的表达量在第6小时增加并达到顶峰,为对照组的1.95倍;第12小时开始下调,第72小时回升到对照组的1.78倍 (图7-b)。SaMBL在肠道中的表达量第6小时开始增加,第12小时达到顶峰,为对照组的1.2倍 (图7-c)。在皮肤中,第6小时表达量开始增加并达到顶峰,为对照组的9.04倍;从第12小时开始下降,第48小时回调到正常水平 (图7-d)。

图7 嗜水气单胞菌感染后SaMBL在七彩神仙鱼不同组织中的表达分析Fig. 7 Expression analysis of SaMBL in different tissues from S. aequifasciatus infected by A. hydrophila
3 讨论
C型凝集素是一种依赖钙离子 (Ca2+) 选择性结合碳水化合物的蛋白质家族,在免疫中发挥重要作用[23]。本研究克隆得到SaCD302和SaMBL。SaCD302推导的氨基酸序列包含一个29个aa的信号肽,可以指引蛋白质在细胞内运输,目前为止,大多数CTLs基因均包含信号肽[24],但是SaMBL却不含信号肽,表明该基因在七彩神仙鱼CTLs家族中可能有一定的特殊性。另外,两个基因均包含一个CTLs结构域,这也是证明其为CTLs基因的重要依据。同源性对比结果表明,SaCD302和SaMBL均与同为慈鲷科的尼加拉瓜湖始丽鱼同源性最高。系统发育树表明,SaCD302和SaMBL均与鱼类聚为一簇,暗示着SaCD302和SaMBL进化的保守性。
SaCD302和SaMBL在组织表达分布上存在显著差异。SaCD302在七彩神仙鱼各组织中均有表达,其中在头肾中表达量最高,其后依次为肝脏、鳃、脑、脾脏等。研究表明CD302在鱼类各组织中广泛分布,且在大多免疫相关组织中表达丰富[25-26],这与本研究结果一致,表明CD302在鱼类先天免疫中发挥着重要作用。松江鲈 (Trachidermus fasciatus) CD302在头肾中表达最高,这与本研究结果相似[25]。SaCD302在七彩神仙鱼头肾中高表达,原因可能是头肾为鱼类的重要免疫器官,含有淋巴细胞和吞噬细胞等多种免疫细胞[27]。在虹鳟 (Oncorhynchus mykiss) 和草鱼 (Ctenopharyngodon idellus) 的研究中,MBL在肝脏和脾脏表达较高[28-29]。本研究中,SaMBL在皮肤中显著表达,而在肠道、肝脏、脾脏和性腺中的表达量却非常低,表明与其他鱼类相比,MBL在七彩神仙鱼的皮肤中可能扮演更加重要的角色。鱼类皮肤上分泌的黏液几乎覆盖在鱼体所有表面,构成其与外界直接接触的第一层免疫防御,保护鱼体免遭外界环境中的病菌、寄生物和病毒侵袭[30-31]。SaMBL在皮肤中的高表达暗示着其可能在鱼类黏膜免疫中发挥重要作用。
松江鲈的TfCD302广泛存在于各组织中,且头肾中的表达量最高,在鳗弧菌的刺激下,松江鲈的鳃、皮肤、头肾、肝脏中的CD302均上调表达[25]。哈维氏弧菌 (V. harveyi) 刺激花鲈 (Lateolabrax japonicus) 后,LjCD302在头肾、肠、鳃、脾和肝中的表达量显著上调[26]。CD302可能在鱼类抵抗病原入侵的先天免疫应答中具有重要作用。本研究在嗜水气单胞菌的刺激下,检测了七彩神仙鱼免疫组织中SaCD302 mRNA的时序表达。SaCD302的表达水平在皮肤和肠道中总体呈现先上升后下降的趋势,而在头肾和脾脏中均出现了在第48小时回升,第72小时下降,推测SaCD302在头肾和脾脏中的免疫反应更加持久。脾脏中第6小时表达量达到峰值,肠道、皮肤和头肾在第12小时达到峰值,推测可能与嗜水气单胞菌刺激后免疫组织发挥的时效和作用原理有关。而各组织表达量在每个阶段均显著高于对照组,表明SaCD302在嗜水气单胞菌感染时发挥了重要的免疫防御作用。
SaMBL在嗜水气单胞菌刺激后,肠道和脾脏的表达变化低于其他组织,可能是因为其在组织中的表达本身较低。草鱼MBL在嗜水气单胞菌感染后,在头肾中的表达在第4小时达到最大值,然后开始逐渐降低至正常水平[29]。这与本研究结果相似,七彩神仙鱼在病原感染后,头肾在第6小时表达量达到峰值,为对照组的1.95倍,之后开始降低。而皮肤中同样在第6小时达到峰值且显著高于对照组,为对照组的9.04倍,表明七彩神仙鱼MBL在皮肤里抗菌免疫中可能发挥更显著的作用。总之,SaCD302和SaMBL在病原感染后的大多组织中均有表达上调,在感染24 h后均下降到对照组水平,表明嗜水气单胞菌引起的感染可能在第24小时结束第一轮感染。但SaCD302和SaMBL不论是组织表达,或是病原刺激下的免疫应答模式,均有不同程度的差异,这暗示在七彩神仙鱼亲代抚育阶段,两个基因可能都对亲鱼先天免疫反应发挥作用,但作用机制不同。
本研究从七彩神仙鱼皮肤中克隆得到SaCD302和SaMBL 基因,对其进行了结构域和进化树分析,并测定了SaCD302和SaMBL在不同组织以及嗜水气单胞菌感染后的免疫应答。结果表明,在病原菌的刺激下,SaCD302和SaMBL均可以迅速作出应答,但应答模式不同,表明两个CTLs基因可能在七彩神仙鱼抗病原入侵时发挥不同作用,具体机制还有待进一步的功能验证。
