莲子抗性淀粉与双歧杆菌的黏附作用
2022-12-20刘霞薛旭亮缪松柳国霞邓凯波
刘霞,薛旭亮,缪松,柳国霞,邓凯波*
(1.福建农林大学 食品科学学院,福建 福州 350002;2.中国-爱尔兰国际合作食品物质学与结构设计研究中心,福建 福州 350002;3.福建省特种淀粉品质科学与加工技术重点实验室,福建 福州 350002;4.爱尔兰农业部Teagasc食品研究中心,爱尔兰 科克 P61C996;5.中国科学院 微生物生理与代谢工程重点实验室,北京 100101)
抗性淀粉(resistant starch,RS)是一种益生元,广泛存在于淀粉及含淀粉的食物中,具有可被结肠细菌发酵并生成短链脂肪酸的特性,可在肠道内作为碳源促进双歧杆菌的生长[1-2]。双歧杆菌是人和动物肠道中具有重要生理功能的一类益生菌,对宿主发挥生物屏障、内毒素血症、抗肿瘤、抗衰老、促进机体免疫机能等生理功能[3-4]。肠道菌群紊乱会引发肠道疾病,严重影响人体健康。作为肠道内的优势菌群,双歧杆菌对肠道菌群的调节起着至关重要的作用[5]。但人体内的双歧杆菌数量会随着个体年龄增长或肠道内环境的影响下降,导致肠道菌群失调[6]。
有研究表明,高直链玉米抗性淀粉可提高双歧杆菌在低pH值、胆盐环境及小鼠肠道中的存活能力,提出细菌对淀粉的黏附作用可能是提高菌体存活能力的机制之一[7]。因此,可以开发一种淀粉基益生元来促进肠道中双歧杆菌增殖,调节人体肠道菌群平衡。但目前对抗性淀粉与双歧杆菌的黏附机制尚不清楚,推测具有黏附性的双歧杆菌与淀粉的结合机制涉及一种特异性的细胞表面蛋白[8]。莲子中直链淀粉含量高,有利于形成Ⅲ型抗性淀粉(RS3),Ⅲ型莲子抗性淀粉(lotus seed resistant starch,LRS3)稳定的双螺旋结构、粗糙的沟壑状表面和空腔凹陷可以与双歧杆菌更好的黏附[9]。
目前关于LRS3的研究多为制备方法优化及其益生功能,而抗性淀粉与双歧杆菌黏附机制的研究相对较少。本研究以莲子抗性淀粉和双歧杆菌为研究对象,通过分析抗性淀粉与双歧杆菌黏附作用的影响因素来探究两者之间的黏附机制,以期为制备二者的高黏附体系提供理论依据。
1 材料与方法
1.1 材料与试剂
鲜莲:绿田(福建)食品有限公司;短双歧杆菌(Bifidobacterium breve)ATCC 15700、长双歧杆菌(Bifidobacterium longum)ATCC 15697、两歧双歧杆菌(Bifidobacterium bifidum)ATCC 2952、青春双歧杆菌(Bifidobacterium adolescentis)ATCC 15703:广东省微生物菌种保藏中心;胃蛋白酶(3 000 U/g~3 500 U/g)、α-淀粉酶(100 000 U/g)、糖化酶(100 000 U/g)、胆盐(胆酸含量65%):北京索莱宝科技有限公司。
1.2 仪器与设备
SW-CJ-2FD型双人单面垂直净化工作台:苏州净化设备有限公司;Allegra X-30R型台式高速离心机:美国Beckman Coulter公司;SpectraMax i3x型酶标仪、Utech PH700型pH计:赛默飞世尔科技有限公司;NanoZS90型马尔文粒度分析仪:英国Malvern公司;HH-6型数显恒温水浴锅:常州国华电器有限公司。
1.3 方法
1.3.1 微波法制备莲子抗性淀粉
参照Zhang等[10]的方法,将一定质量的莲子淀粉配制成一定浓度的淀粉乳。将淀粉乳置于微波炉中,640 W加热2 min~4 min,使淀粉糊化。65℃烘干,粉碎,过80目筛,得到粗提LRS3。加入α-淀粉酶、葡萄糖淀粉酶,用蒸馏水洗涤沉淀,得到酶解纯化的LRS3。
1.3.2 菌种活化
挑取冷冻保藏的4种供试双歧杆菌粉末在TPY琼脂培养基上划线培养,经48 h厌氧培养后,挑取单菌落于5 mL TPY液体培养基中,37℃静置厌氧培养36 h,再以2.0%接种量转接到5 mL TPY液体培养基中,连续传代2次~3次,使菌株完全活化。
1.3.3 LRS3与双歧杆菌黏附率的测定
将对数期双歧杆菌菌体用0.1 mol/L磷酸盐缓冲液(phosphate buffer saline,PBS)(pH7.0) 洗涤 2 次后,重悬于相同缓冲溶液中至细胞终浓度约为107CFU/mL。取其中2 mL细胞重悬液与LRS3重悬液(10 g/L,0.1 mol/L PBS,pH7.0)在直径1 cm试管中同体积彻底混匀。室温下静置1 h至淀粉沉淀后,在液面下0.5 cm处吸取两份150 μL样液,酶标仪测定540 nm下吸光值a,以PBS为空白对照。黏附于抗性淀粉上细菌比例(黏附率)按公式(1)计算。
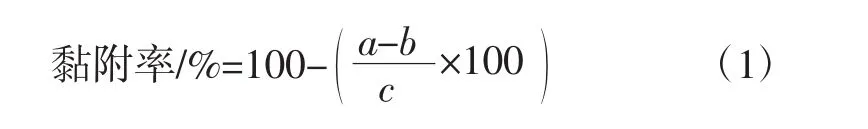
式中:b为不添加细菌的淀粉重悬液OD540值;c为不添加淀粉的细菌重悬液OD540值[11]。
1.3.4 黏附作用影响因素研究
以1.3.3中所述黏附率的测定方法为基础,进行多条件变量试验设计[11-12],系统探讨生长介质、疏水相互作用、静电相互作用、细菌表面蛋白及温度等条件对LRS3与4株供试双歧杆菌黏附作用的影响。黏附试验的基本操作同1.3.3黏附率测定方法,其中各变量条件试验中使用的处理方法见表1,每个试验3次重复。
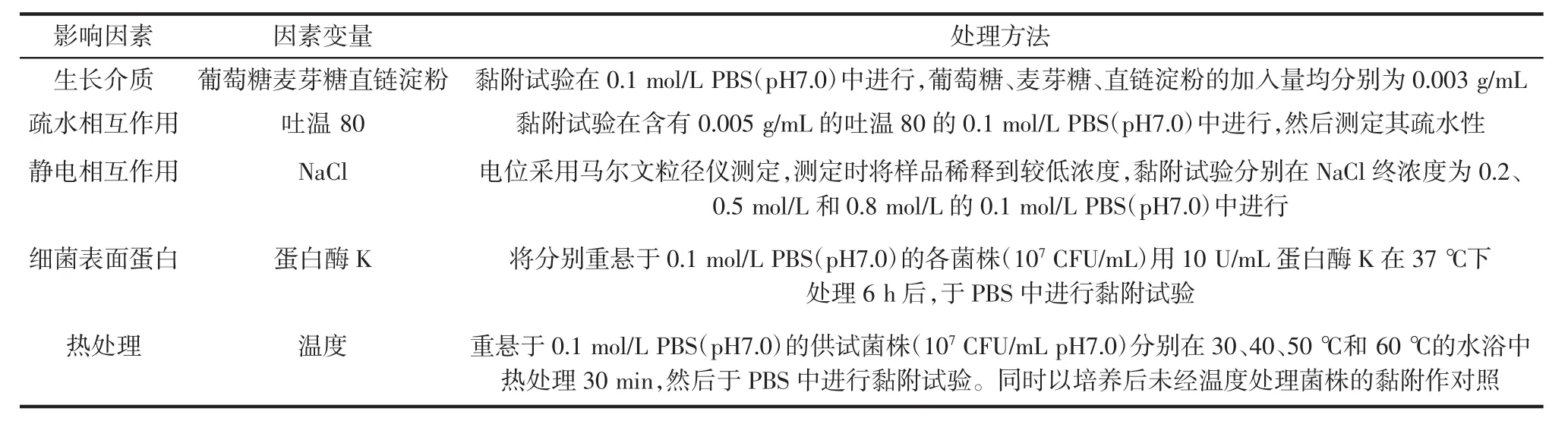
表1 黏附影响因素变量试验的处理方法Table 1 Treatments used in the experiments of variable adhesion influenceable factors
1.3.5 菌体表面疏水性的测定
疏水性是通过细菌对碳氢化合物的黏附性测试来测定的[13-14],具体来说,将菌悬液在室温下离心(6 000 r/min,10 min),收集的菌体经 0.1 mol/L PBS(pH7.0)洗涤2次后,重悬于无菌0.1 mol/L KNO3溶液中。在540 nm处调节吸光度为0.50±0.02,记作A0。取3 mL菌悬液与1 mL二甲苯混匀后在室温静置10 min(此时形成两相体系)。将两相体系涡旋振荡2 min后再静置20 min,重新形成两相体系(水相和有机相)。小心吸取水相,在540 nm下测定吸光度(A)。按公式(2)计算细菌表面疏水性,重复3次试验取平均值。
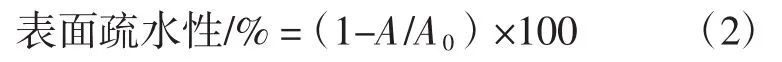
1.3.6 模拟上消化道条件对黏附的影响
为了确定双歧杆菌通过胃和小肠对LRS3黏附的影响,在模拟上消化道生理条件的体外条件下进行了黏附试验[15]。pH值、胃蛋白酶、胰蛋白酶和胆汁对黏附影响的处理方法如表2所示。所有试验均重复3次。
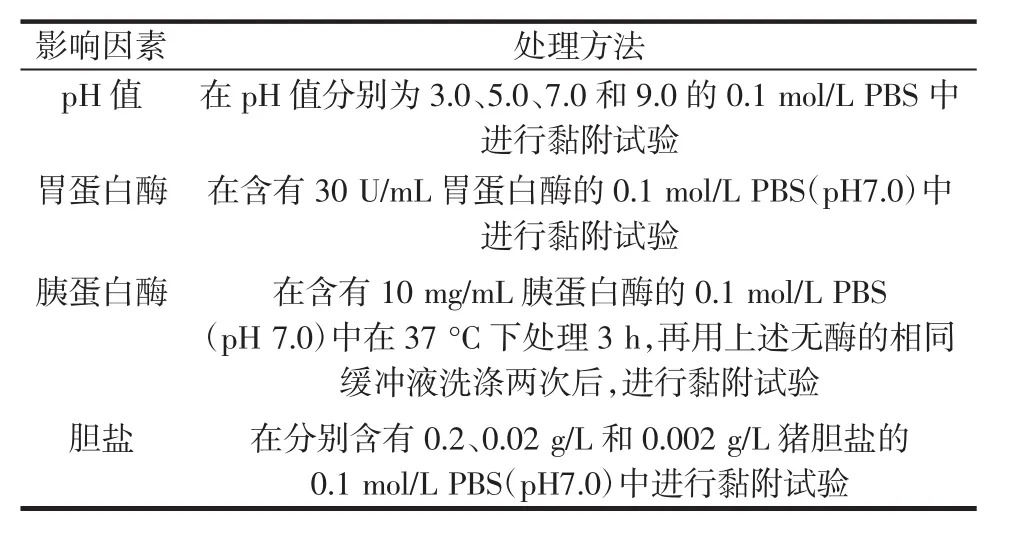
表2 模拟上消化道条件下黏附的处理方法Table 2 Treatments for adhesion determination under simulated upper digestive tract
1.4 数据处理
采用Origin 8.0软件对试验结果进行统计和制图,SPSS 20.0进行单因素方差分析。
2 结果与分析
2.1 生长介质对黏附作用的影响
处于对数期的4株双歧杆菌均可与LRS3发生黏附反应,其黏附率分别为82.83%(B.longum)、85.27%(B.adolescentis)、71.13%(B.breve) 和 75.13%(B.bifidum)。其中,前两种为成年人体内的优势菌株,后两种为婴儿体内的优势菌株。黏附率的差异说明菌株与LRS3的黏附之间存在菌株差异性,也可能存在不同的黏附机制。通过筛选不同双歧杆菌的表面黏附素已发现,不同菌株表面可能存在不同的与黏附有关的蛋白质[16],而这有可能是影响菌株与淀粉黏附的因素之一。不同生长介质对LRS3黏附双歧杆菌的影响如图1所示。

图1 不同生长介质对LRS3与双歧杆菌黏附作用的影响Fig.1 Influence of growing medium on the adhesion between LRS3 and Bifidobacteria
从图1中可以看出,加入的生长介质均对LRS3与双歧杆菌的黏附作用产生了抑制作用,且其抑制作用的强度顺序为直链淀粉>麦芽糖>葡萄糖。其中,葡萄糖对LRS3与B.longum黏附的抑制作用最大,黏附率降低了7.87%,而对LRS3与B.breve黏附的抑制作用最小,只降低了4.83%。麦芽糖对LRS3与B.adolescentis黏附的抑制作用最明显,黏附率降低了25.2%,而LRS3与B.breve、B.longum和B.bifidum 黏附率则分别降低了15.3%、19.8%和15.6%。故总体上麦芽糖对黏附体系的抑制程度比葡萄糖高。与对照组相比,直链淀粉对4株菌的抑制作用最明显,其中直链淀粉显著抑制了LRS3与双歧杆菌的黏附(P<0.05),其对LRS3与B.longum黏附的抑制作用最大,黏附率降低达38.97%。葡萄糖、麦芽糖、直链淀粉均可作为双歧杆菌生长的碳源,但将这些碳源添加到LRS3与双歧杆菌的黏附体系中时,由于不同双歧杆菌的碳源运输系统不同[16-17],导致LRS3与双歧杆菌的黏附率在含有不同碳源的黏附体系中表现出不同程度的差异。尽管葡萄糖在小程度上抑制了黏附,但随着淀粉聚合物长度的增加,抑制程度随之增加。这表明黏附体系中的关键因子可能对较大分子具有更高的亲和力;也可能是较大淀粉聚合物的更大空间位阻,更有效的阻断了淀粉颗粒与细菌表面的黏附相互作用。这与Ghalia等[18]研究结果一致。
2.2 细胞表面疏水性对黏附作用的影响
细菌表面疏水性取决于细菌表面非极性基团的数量,多与黏附现象有关[8]。菌体表面的理化性质很大程度上依赖于表面疏水程度,较高的表面疏水性意味着在水相体系中与非极性物质具有较高的结合能力[19]。图2展示了供试菌株的细胞表面疏水性对LRS3与菌株黏附作用的影响。

图2 疏水性对LRS3与菌株黏附作用的影响Fig.2 Influence of hydrophobic interaction on the adhesion between LRS3 and Bifidobacteria
菌株表面的疏水性与LRS3之间的黏附率存在弱相关性。由图2可知,在成人肠道中处于高丰度水平的B.longum和B.adolescentis[20],后者与LRS3黏附率更高,而疏水性却低于前者。由此可见双歧杆菌细胞表面疏水性在黏附LRS3的过程中会产生一定的作用,但仍存在其他影响因素。具有疏水性的细菌更倾向于黏附具有疏水性表面的碳源[21]。Niderman-Meyer等[22]使用两种不同的有机溶剂作为疏水相,发现霍乱弧菌(Vibrio cholerae)对不同碳氢化合物的黏附程度不同。细菌表面疏水性对LRS3黏附作用的影响也可能是由于黏附条件的不同而不同,细胞在中性pH值或稀释的缓冲液中可能不会黏附在碳氢化合物上,但在低pH值或高离子强度介质中则会黏附。
2.3 静电相互作用对黏附作用的影响
静电相互作用介导黏附需要带有相反电荷的离子,表3为不同离子强度下LRS3与供试菌株黏附体系的电位值变化情况。

表3 不同离子强度下LRS3与供试菌株黏附体系的电位值Table 3 Electric potential values of adhesion systems of LRS3 and Bifidobacteria under different ion strengths mV
经测定,LRS3与双歧杆菌均带负电荷,因此其黏附体系也均为负电位。在添加了NaCl后,引入的阳离子与体系原有粒子电荷相反,虽中和了一部分体系负电,但黏附体系仍带负电荷,且电位值均随NaCl浓度的增高而显著减小(P<0.05)。
长距离静电力可能影响细菌黏附到LRS3表面的初始阶段,这种相互作用会以双向方式(即吸引力或排斥力)影响黏附过程[23]。图3为添加不同浓度NaCl对二者黏附作用的影响。

图3 离子强度对LRS3与菌株黏附作用的影响Fig.3 Influence of ion strength on the adhesion between LRS3 and Bifidobacteria
由图3可知,当NaCl浓度在0~0.5 mmol/L时,供试菌的黏附率大体随离子强度增大而降低,而当NaCl浓度为0.8 mmol/L时,黏附率却均呈显著升高趋势。阳离子的加入可以把双歧杆菌细胞壁上的负电荷和LRS3的负电荷通过静电作用结合起来,起到中间桥梁的作用。研究结果表明,当阳离子的加入量达到一定的浓度时,会使体系的黏附率相对提高,这表明静电相互作用对黏附体系具有一定的影响。
2.4 蛋白酶K处理对黏附作用的影响
蛋白酶K处理的双歧杆菌与LRS3的黏附作用见图4。
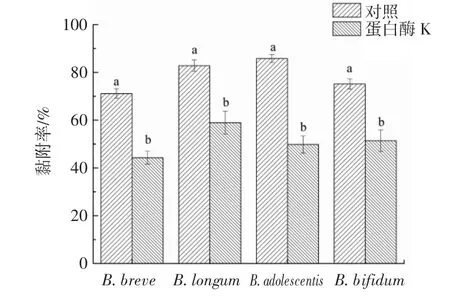
图4 蛋白酶K处理的双歧杆菌与LRS3的黏附作用Fig.4 Adhesion of LRS3 to proteinase K treated Bifidobactiria
由图4可知,蛋白酶K对LRS3与4株双歧杆菌的黏附能力均产生了显著的抑制作用(P<0.05),其中对B.adolescentis的抑制作用最明显,黏附率降幅达35.99%;对LRS3与B.longum和B.bifidum的抑制作用相对较小,降幅约23%。有研究发现,LRS3与双歧杆菌的黏附可能是由于细菌细胞表面蛋白或糖蛋白没有黏附任何与淀粉颗粒有关的蛋白质或肽链[24]。蛋白酶K对菌株的处理可对其表面蛋白产生降解作用,本研究中黏附水平的显著降低表明,在双歧杆菌与LRS3的黏附中,细菌表面蛋白可能参与了该过程,并起到较为关键的作用。
2.5 菌株热处理对黏附作用的影响
不同温度处理后的双歧杆菌与LRS3的黏附作用见图5。
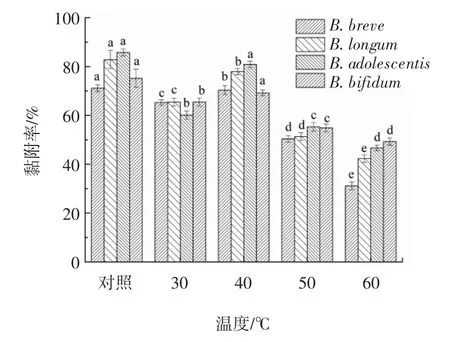
图5 不同温度处理后的双歧杆菌与LRS3的黏附作用Fig.5 Adhesion of Bifidobacteria to LRS3 after different temperature treatment
由图5可知,菌株热处理对黏附率存在较大影响。总体上来说,40℃~60℃组双歧杆菌与LRS3的黏附率随温度升高呈显著下降趋势(P<0.05),其中,各菌株40℃组的黏附率均最接近室温对照组,B.adolescentis组黏附率最高(80.87%),B.bifidum组最低(69.16%);60℃时的黏附率受抑制程度最大,LRS3与B.longum的黏附率降幅达40.53%,推测其可能原因是高温使菌体表面蛋白(包括可能的黏附蛋白)变性[25],低温则会使蛋白质的活性降低,导致黏附率下降。而30℃组黏附率的普遍低值,其原因可能为双歧杆菌的最适宜生长温度为37℃~41℃,过低(30℃)或过高(50℃及以上)温度均可能影响菌株各成分生理功能的发挥,但30℃处理相对高温对黏附率的抑制程度较小,说明LRS3与双歧杆菌结合的关键黏附因子对高温更敏感,这也从一个侧面说明了LRS3与双歧杆菌的黏附可能与细胞的表面蛋白有关。
2.6 模拟上消化道条件对黏附作用的影响
2.6.1 pH值对黏附作用的影响
宿主菌对淀粉的物理性黏附可能是其利用碳源的前提条件,与碳源的利用性有关[18]。LRS3和双歧杆菌作为较好的人体益生元和益生菌物质,其黏附体系在胃和小肠逆环境中的适应性也关系到二者益生功能的作用程度。pH值对LRS3与双歧杆菌黏附作用的影响见图6。

图6 pH值对LRS3与双歧杆菌黏附作用的影响Fig.6 Effects of pH value on adhesion between LRS3 and Bifidobacteria
由图6可知,与对照组相比,pH值为3时LRS3与4株双歧杆菌黏附率的抑制程度均最为显著(P<0.05),其中 B.bifidum组黏附率由 75.17%降至16.82%,降幅达68.34%。由于双歧杆菌的最适宜生存环境为pH6.5~7.0,高酸性环境可导致菌株酶活被抑制、蛋白质变性及代谢过程改变等,因此高酸性环境可能使菌株表面蛋白等黏附关键物质的功能受到影响,而致黏附率大幅下降。然而,pH5、pH7和pH9环境则对LRS3与供试菌的黏附率影响较小,尤其是在双歧杆菌最适宜pH值时黏附率相对最高,即4株供试菌与LRS3均在pH7环境下表现出最佳黏附效果。因此可以推断,体系的黏附率与环境pH值有关,且在菌株可表现出最佳活力的环境中效果最好。
2.6.2 消化道蛋白酶对黏附作用的影响
蛋白酶处理对LRS3与双歧杆菌黏附作用的影响见图7。
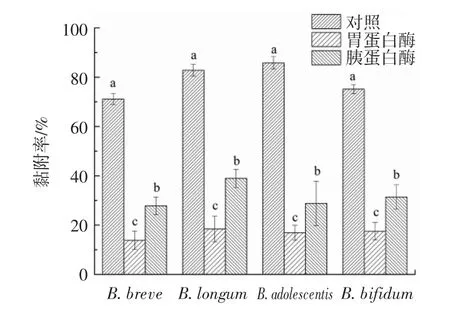
图7 蛋白酶处理对LRS3与双歧杆菌黏附作用的影响Fig.7 Effects of pretease treatment on adhesion between LRS3 and Bifidobacteria
由图7可知,LRS3与双歧杆菌的黏附对消化道胃蛋白酶和胰蛋白酶均敏感,酶处理会使蛋白质的活性降低,从而降低二者的黏附率。用胃蛋白酶处理时发现,LRS3与4株双歧杆菌的黏附率都受到明显的抑制(P<0.05),与LRS3黏附率较好的B.longum在胃蛋白酶的处理下黏附率为18.93%,其余3株的黏附率也都仅为15%左右,推测二者黏附体系在通过胃的过程中损失较大。胰蛋白酶对二者黏附体系也有明显的抑制作用(P<0.05),与胃蛋白酶相同,胰蛋白酶对LRS3与B.adolescentis的黏附抑制率也为最高,黏附率从85.8%降至28.25%;LRS3与B.breve的黏附率也降低了43.3%。这与蛋白酶K、热处理及pH值影响黏附作用的结果互相印证,说明LRS3与双歧杆菌之间的黏附关键因子可能包括黏附相关蛋白。胰腺分泌物含有淀粉分解酶,影响淀粉颗粒表面,在胰腺消化后观察到黏附水平显著降低,这表明黏附可能涉及淀粉颗粒表面一些糖苷的特异性[22]。
2.6.3 消化道胆盐对黏附作用的影响
胆盐浓度对LRS3与双歧杆菌黏附作用的影响见图8。

图8 胆盐浓度对LRS3与双歧杆菌黏附作用的影响Fig.8 Effects of bile salt concentration on adhesion between LRS3 and Bifidobacteria
由图8可知,不同浓度的消化道胆盐也均对LRS3与双歧杆菌的黏附产生了抑制作用。其中,胆汁浓度0.2 g/L的胆盐浓度对体系的黏附抑制程度最高(P<0.05),LRS3与B.longum的黏附降低至56.63%。而随着胆盐浓度的不断降低,其对LRS3与双歧杆菌的黏附抑制作用越来越弱,当胆盐浓度为0.002 g/L时,B.breve和B.longum两组的黏附率已与其对照组无显著差异(P>0.05)。对此,研究发现胆盐在到达一定浓度后会抑制二者的黏附,但是随着浓度的降低,抑制作用几乎不存在。肠道中的总胆汁浓度为1 g/L~10 g/L,但游离胆汁浓度因在肠道的不同区域和人体摄入脂质的量而异。当胆汁分泌量增加时,胆汁会抑制双歧杆菌对不溶性膳食纤维的黏附。相反,胆汁被重吸收后大肠中游离胆汁浓度较低,使双歧杆菌容易黏附在不溶性膳食纤维上。而且在胆汁和低pH值存在的情况下,抗性淀粉的加入可促进双歧杆菌在大鼠肠道中的存活[26]。
3 结论
细菌对颗粒淀粉的黏附作用是其降解的首要步骤,也是有效降解的先决条件。LRS3与双歧杆菌间的良好黏附有利于改善二者的营养互作关系。对不同环境条件下LRS3与双歧杆菌黏附作用的研究结果表明,LRS3与双歧杆菌的黏附作用不受静电相互作用的影响,而疏水相互作用对二者黏附体系的影响存在菌株差异性。同时,在低pH值、蛋白酶处理及高胆盐浓度等条件下,体系的黏附作用会受到显著抑制,这与在V.cholerae等菌中的现有结果一致,推测LRS3与双歧杆菌的黏附作用机理可能也与一类特异性的细胞表面蛋白相关,这为进一步揭示黏附机制指明了方向,也为高黏附体系的制备及其在合生元开发方面的应用提供了理论依据。