青藏高原3种特有植物的核型报道
2022-12-18马小磊刘瑞娟陈世龙高庆波
马小磊, 刘瑞娟, 曹 倩, 邢 睿, 陈世龙, 高庆波
(1. 中国科学院西北高原生物研究所青海省作物分子育种重点实验室, 西宁 810008;2. 中国科学院大学, 北京 100049)
青藏高原拥有多样的自然环境和丰富的生物资源,是我国特有物种最丰富的地区[1]。我国高等植物的细胞学研究仍有大量空白,仅有约22.6%的被子植物有细胞学研究[2];青藏高原地区植物的细胞学研究较为薄弱,有细胞学研究的物种仅占该地区物种总数的10%[3]。
黑虎耳草[Micranthesatrata(Engl.)Losinsk.]、黑蕊虎耳草[M.melanocentra(Franch.)Lonsinsk.]在以往的分类系统中隶属于虎耳草科(Saxifragaceae)虎耳草属(SaxifragaTourn. ex L.)小花组(Sect.Micranthes)[4-6]。形态学和分子生物学等多方面的证据支持小花组从虎耳草属独立,成立新的属MicranthesHaw.[6-8],该属暂时没有确定的中文名。Micranthes大约有70个物种,主要分布于北美安第斯山脉,我国分布有13种,黑虎耳草(M.atrata)仅分布于祁连山及其周边区域;黑蕊虎耳草(M.melanocentra)主要分布于青藏高原东缘和南缘[5]。前期研究表明Micranthes的物种核型复杂,染色体基数和倍性存在较大差异,且存在非整倍体和多倍体;国内分布的Micranthes物种有6个已有细胞学方面的研究[9-10]。黑蕊虎耳草(M.melanocentra)有多倍体[10]和非整倍体[11]的报道;黑虎耳草(M.atrata)暂未看到相关研究报道。
祁连獐牙菜(SwertiaprzewalskiiPissjauk.)隶属于龙胆科(Gentianaceae)獐牙菜属(SwartiaL.)。獐牙菜属的系统发育关系一直未能解决,分子证据显示獐牙菜属并非单系类群,且分子系统学和形态分类学出现明显的分歧[12-13]。獐牙菜属大约有150个物种,75个物种分布于我国。祁连獐牙菜(S.przewalskii)分布于祁连山及其周边区域[14]。目前,獐牙菜属29个物种有核型研究[9-10],其中20个物种在中国有分布。獐牙菜属的物种多为二倍体,但属内染色体基数差异较大,部分物种染色体基数在不同报道中存在差异[15-16]。祁连獐牙菜暂未看到相关研究报道。
黑虎耳草(M.atrata)、黑蕊虎耳草(M.melanocentra)和祁连獐牙菜(S.przewalskii)是重要的藏药资源植物,3种植物开发利用和多样性保护相关的研究较为欠缺。近年来部分学者对3种青藏高原特有植物的分子遗传多样性[17]、生物保护[18]等方面做了部分工作,但细胞学等基础研究资料仍是空白。本研究以采集自青海和四川的黑虎耳草(M.atrata)、黑蕊虎耳草(M.melanocentra)和祁连獐牙菜(S.przewalskii)的多个居群的种子为材料,开展核型分析,以期为3个物种的开发利用及保护提供细胞学资料。
1 材料与方法
材料于2018—2019年采集自青海省和四川省,具体采集信息见表1。所有标本由中国科学院西北高原生物研究所高庆波研究员鉴定,凭证标本保存于西北高原生物研究所标本馆(HNWP)。
采用滤纸萌发法和压片法对材料进行处理。种子萌发:种子用50 mg/L的赤霉素处理2 h,置于垫有一张灭菌滤纸的培养皿中,用50 mg/L的赤霉素溶液浇透,放置于培养箱中萌发。培养箱的程序设定为16 h光照+8 h黑暗,25 ℃恒温。萌发期间每隔2~3天补充一次培养液。预处理:待幼苗根尖长到0.3~0.5 cm,将幼苗放置于喷有少量培养液的EP管中,室温下用500 000~600 000 Pa压强的一氧化氮(NO)处理2 h。固定:处理后的幼苗置于4 ℃冰箱中,加入卡诺试剂(冰醋酸∶无水乙醇=1∶3)固定2 h左右。染色:固定后的幼苗置入45%的冰醋酸处理5 min以上,将处理好的幼苗根尖切下(约1 mm),放于载玻片上,滴一滴改良石炭酸品红染液,染色3 min后压片观察。染色液配置方法和压片操作参考《植物染色体研究技术》[19]。镜检:制作的临时装片放置于显微镜下进行观察和计数,并拍摄染色体形态较好、分散均匀的有丝分裂中期细胞。
每个物种选取30个以上细胞进行染色体计数(黑虎耳草样本数少,所统计细胞数目不足30个),以其中80%以上细胞具一致且恒定的染色体数目作为该物种的染色体数。每个物种挑选5张染色体分散较好的中期细胞照片,使用KaryoType 2.0软件[20]测量染色体长度。黑虎耳草和黑蕊虎耳草的染色体极小,大部分染色体无法判断着丝粒位置,依据李懋学和张敩方[19]的标准,两物种属于具小染色体的植物,故仅测量染色体绝对长度,并求取平均值用于计算染色体相对长度、单倍体染色体总长以及最长和最短染色体长度比。祁连獐牙菜染色体大,故测量染色体绝对长度、长臂和短臂绝对长度,求取平均值并参考李懋学等[21]的标准进行染色体核型分析。判断核型类型依据Stebbins等[22]的标准,判断着丝点位置和染色体类型依据Levan等[23]的标准;同时计算着丝粒指数变异系数(CVCI)、染色体长度变异系数(CVCL)[24]、平均着丝粒不对称系数(MCA)[25]、核型不对称指数(As. K%)[26];绘制核型模式图。

表1 物种采集信息
2 结果与分析
黑虎耳草的2个居群采集于青海省祁连山国家公园,染色体数目均为2n=2x=22,染色体相对长度范围为6.02%~12.74%(表2),单倍体染色体总长THL=16.26 μm,最长和最短染色体长度比为1.93。黑虎耳草的花及中期染色体形态见图1 (a)和 (b)。
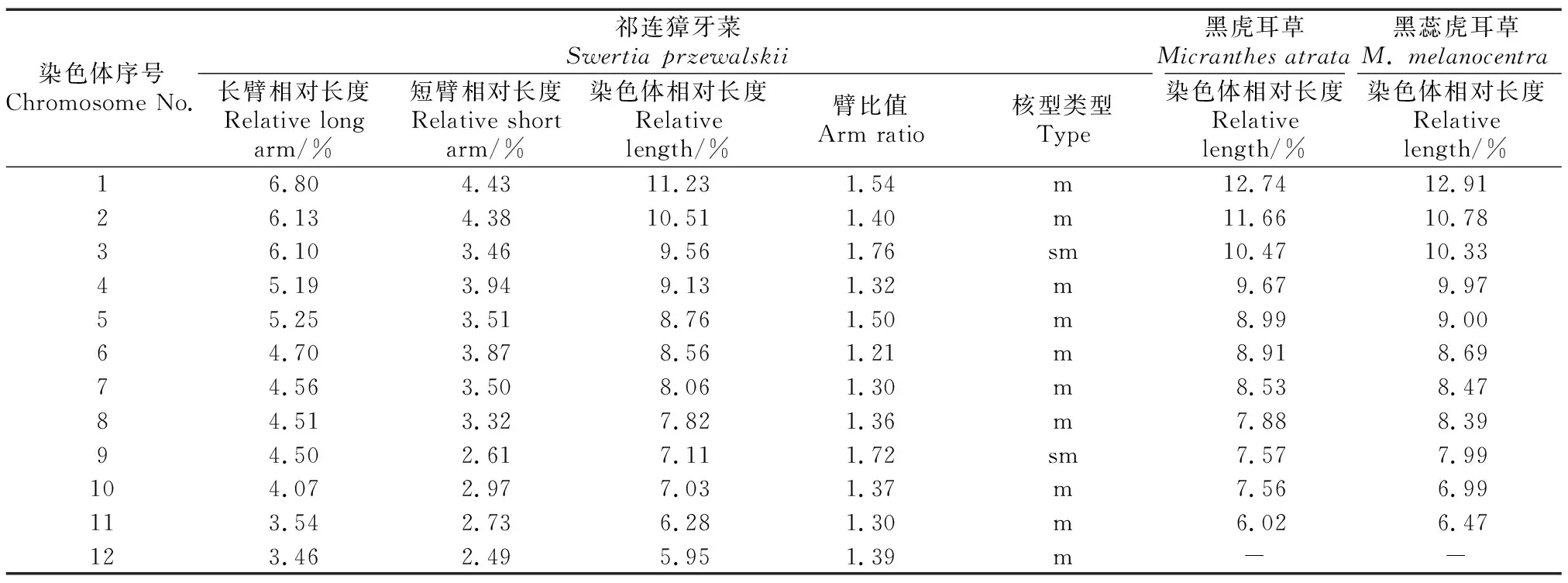
表2 3个物种的核型参数
黑蕊虎耳草的5个居群采集于四川和青海,染色体数目均为2n=2x=22,染色体相对长度范围为6.47%~12.91%(表2),单倍体染色体总长THL=15.08 μm,最长和最短染色体长度比为2.04。黑蕊虎耳草的花及中期染色体形态见图1 (c) 和(d)。
祁连獐牙菜的5个居群均采集于青海省祁连山国家公园。5个居群的根尖细胞的染色体数目相同,核型公式为2n=2x=24=20m+4sm,由中部着丝点和亚中部着丝点染色体组成,其中第3对和第9对染色体为亚中部着丝点染色体(sm),其余为中部着丝点染色体(m)。染色体相对长度范围为5.95%~11.23%(表2),单倍体染色体总长THL=31.98 μm;最长和最短染色体长度比为2.03,臂比大于2的染色体比例为0,核型类型为1B;核型不对称指数As.K%=58.8%、着丝粒指数变异系数CVCI=6.62、染色体长度变异系数CVCL=19.18、平均着丝粒不对称系数MCA=17.61。祁连獐牙菜的花、有丝分裂中期染色体形态及核型模式图,见图1(e) 、(f) 和(g)。

(a)和(b)黑虎耳草;(c)和(d)黑蕊虎耳草;(e)~(g)祁连獐牙菜。图1 3个物种的花、有丝分裂中期细胞及祁连獐牙菜核型模式图Figure 1 Flower,mitotic metaphase chromosomes of three species and ideograms of Swertia przewalskii
3 讨论与结论
全球有细胞学报道的广义虎耳草属物种有226种,多倍化和非整倍体现象在虎耳草属中较常见[27-28]。以往研究中黑蕊虎耳草的倍性变化较大(2n=22,44,66,88,110 )[10],且存在非整倍体(2n=22,12)[11]。但本研究的5个居群中并没有发现多倍体及非整倍体,与前人研究结果不一致,推测是因为采样区域不同。研究结果揭示分布在青海和四川的黑蕊虎耳草居群中出现多倍体的频率较低。胡晶晶[27]使用流式细胞仪测定了65个虎耳草属植物的染色体倍性和基因组大小,其中黑蕊虎耳草为二倍体,与本研究结果一致。黑蕊虎耳草和黑虎耳草的染色体数目均为2n=22。在Tkach等[8]的分类系统中,两个物种隶属于Micranthessect.Melanocentra(Engl. & Irmsch.)Tkach.,该组的物种Micranthesdivericata(Engl. & Irmsch.)Losinsk.[29]和M.pallida(Wall. Ex Ser.)Losinsk.[10]体细胞染色体数目均为2n=22。 2n=22在该属其他组少见,仅M.micranthidifolia(Haw.) Small[30],隶属于MicranthesHaw. sect.Micranthes。
国内外对獐牙菜属的细胞学研究报道相对较少,现有的研究表明,獐牙菜属的物种多为二倍体,染色体基数变化大(x=7~14)。本研究中祁连獐牙菜5个居群均为2n=24,核型公式为2n=2x=24=20m+4sm,核型类型为1B。獐牙菜属有6种植物染色体数为2n=24,但6个物种均存在种内染色体基数变化[9-10],且獐牙菜属的系统发育关系表明祁连獐牙菜和这6个物种的亲缘关系较远[12-14]。祁连獐牙菜及其近缘种染色体基数不一致,黄荣福等[15]报道了二叶獐牙菜(SwertiabifoliaBatal.)的染色体数目大约为26;何廷农等[16]报道了华北獐牙菜(SwertiawolfgangianaGruning)和二叶獐牙菜均为2n=28;长柄獐牙菜(SwertiapetiolataRoyle ex D. Don)2n=26[10]。造成祁连獐牙菜及其近缘种染色体基数不一致的原因有待进一步的研究。
青藏高原地区低基数二倍体和古多倍体在龙胆科和虎耳草科的出现频率很高[3,15]。黑虎耳草、黑蕊虎耳草和祁连獐牙菜均为高基数二倍体,即古多倍体。王家坚等[3]的研究表明青藏高原的地质历史和环境特性虽然有利于多倍体的形成,但植物在二倍体水平上的适应机制在青藏高原上依旧是很成功的,本实验结果为该推断提供了数据支撑。3种植物的核型报道补充了青藏高原特有物种的细胞学资料,同时为该物种的开发、利用和保护研究提供了依据。
