木质素的生物学功能及其在畜禽生产中应用的研究进展
2022-12-08余可佳蒋桂韬范志勇
余 建 余可佳 蒋桂韬 范志勇*
(1.湖南农业大学动物科学技术学院,长沙 410128;2.湖南省畜牧兽医研究所,长沙 410131)
木质素几乎存在于所有植物之中,为植物细胞壁三大组成成分之一,全球年产量仅次于纤维素,是地球上第二大生物可再生酚类聚合物[1]。然而,仅就从木材和造纸工业中获得的木质素却不到2%被充分利用,其余大部分都用作低级燃料被焚烧或直接被丢弃[2]。虽然多酚物质在畜牧领域被广泛研究,但木质素却一直被认为是一种抗营养因子,阻碍营养物质消化,也难被肠道微生物发酵。然而,有研究显示木质素富含多种低分子质量酚类单体,发挥益生元的作用,并且可以被肠道菌群代谢,形成对动物体健康至关重要的哺乳动物木脂素,从而在畜禽生产上有着替代抗生素的可能性[3-4]。因此,本文就木质素的理化性质、生物学功能及其在畜禽生产中的应用前景进行综述,旨在为其开发成为畜禽新型饲料添加剂提供理论参考。
1 木质素的结构及性质
1.1 木质素的结构特征
木质素主要由C、H、O 3种元素组成,但其结构极为复杂,无固定分子式[5]。目前研究显示,木质素结构模型中存在着3种不同苯基丙烷单体,分别为香豆醇(p-coumaryl alcohol,H)、松柏醇(coniferyl alcohol,G)和芥子醇(sinapyl alcohol,S)[6](图1)。木质素各单体之间通过醚键(β-o-4、α-o-4、4-o-5)和C—C键(β-1、β-2、β-5、β-β、β-6、5-5)相连,而β-o-4连接方式占1/2左右[7-8]。同时,木质素含有多种活性官能团,其中以酚羟基活性最强,其次为羰基、羧基、甲氧基与甲基[9]。图2为木质素部分结构。

图1 木质素主要单体结构
1.2 木质素的来源
除苔藓及菌类外,木质素几乎存在于所有植物细胞壁中,与纤维素和半纤维素以醚键、糖苷键和酯键等共价键的形式彼此连接[11]。不同植物木质素结构因各单体所占比例不同而存在差异。以松柏醇为主聚合而成的G型木质素,大多存在于裸子、蕨类植物;以松柏醇和芥子醇为主聚合而成的G-S型木质素,主要存在于双子叶植物;单子叶植物则主要是香豆醇、松柏醇和芥子醇聚合成的G-S-H型木质素[12]。表1列举了木质素来源及含量。
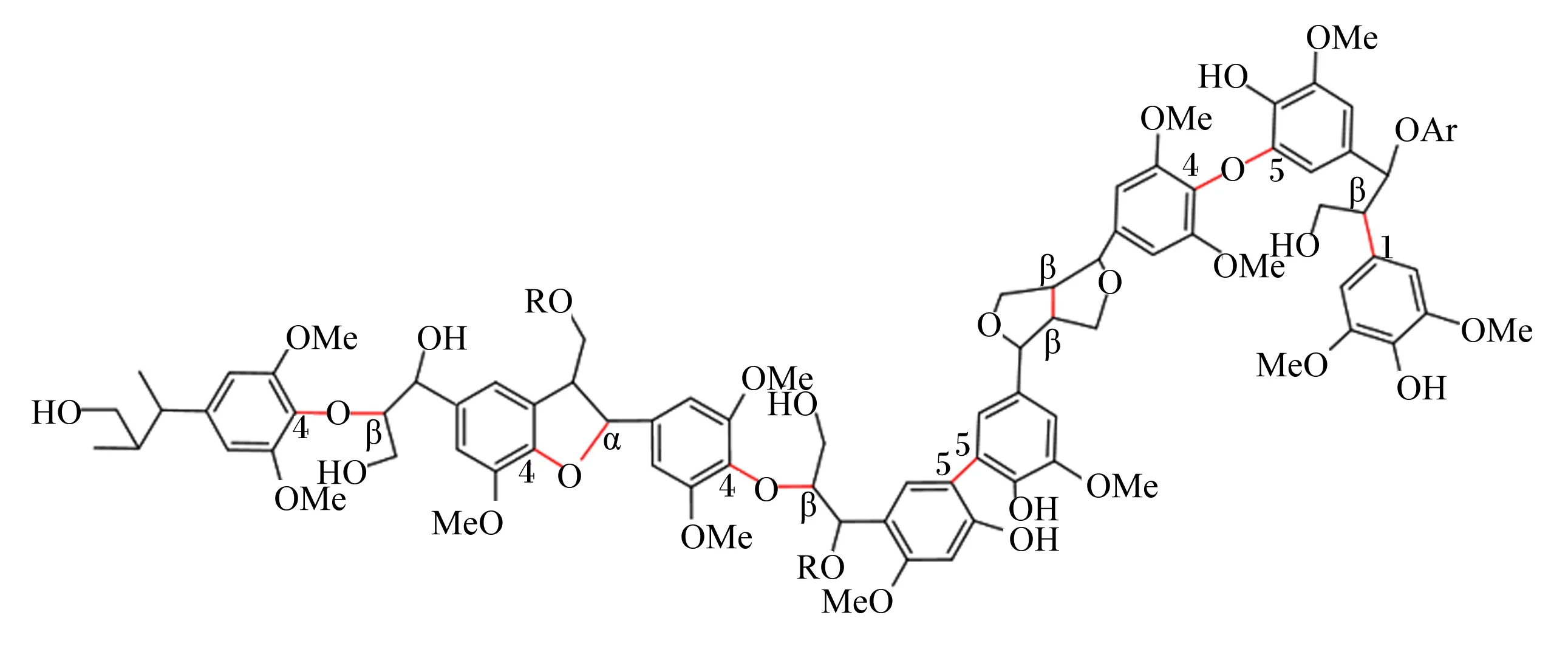
图2 木质素部分结构
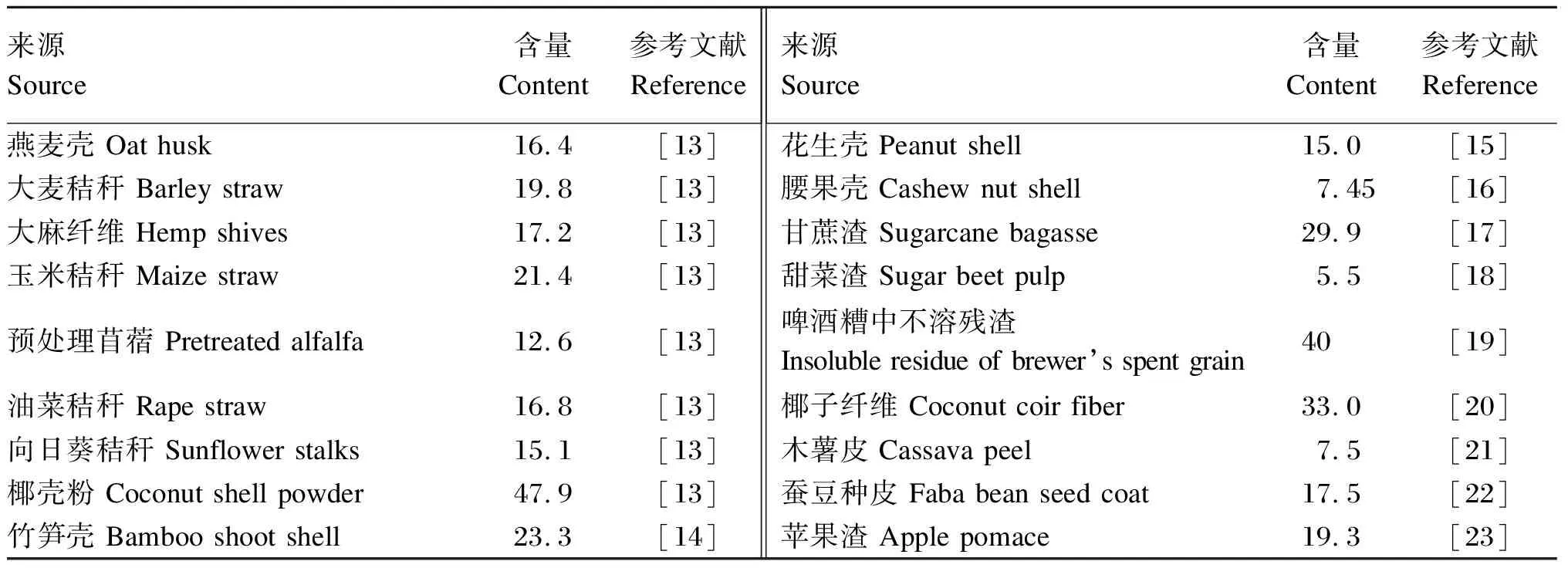
表1 木质素来源及含量
1.3 木质素的理化性质
木质素本身是一种白色或近无色的球形颗粒,但从植物中分离得到的木质素颜色一般呈灰黄至灰褐色。木质素的相对密度为1.35~1.50 g/cm3,热值约26.0 J/g[24],因其存在着大量羟基而有着很强的氢键作用,导致木质素很难与任何溶剂相溶合[25]。同时研究显示,木质素无特定熔点,其玻璃化转变温度差异也较大(89.9~174 ℃)[26]。
木质素的化学性质与其活性官能团紧密相关。芳香基、酚羟基、甲氧基、醇羟基和羰基等重要活性基团,可使木质素发生氧化、还原、环氧化、水解、酯化、磺化和缩聚等化学反应,且其中的酚羟基还能引发自由基反应[27]。
2 木质素的生物学功能
2.1 抑菌作用
酚类化合物一直被用作食品防腐剂,其抑菌作用已被充分证实[28],而木质素存在11种不同酚类单体结构[29],含有大量酚羟基和甲氧基等含氧抑菌官能团,同时其侧链α碳与β碳之间的双键结构以及γ碳上的甲基也能显著增强其抑菌效果[30]。虽然对于木质素抑菌机制尚未得出明确论断,但Jung等[31]提出,木质素中的多酚结构可以破坏磷脂双分子结构使细菌裂解,并释放细胞内容物。除此之外,Domínguez-Robles等[32]进行试验发现,在所测试材料中,含有软木硫酸盐木质素的复合材料与常见医用抗菌材料聚合物聚丁二酸丁二醇酯相比,抗金黄色葡萄球菌黏附能力达到90%。由此推测,木质素的另一种抑菌方式可能是阻止细菌黏附。
目前对于木质素的体外研究表明,从甘蔗渣、棉花秸秆与油棕空果串等中提取出的木质素可不同程度地抑制表皮葡萄球菌[33]、枯草芽孢杆菌、大肠杆菌、鼠伤寒沙氏杆菌和金黄色葡萄球菌的生长繁殖[34]。针对木质素在体内的研究还鲜有报道,但从以往研究显示,松木木质素具有抑制小鼠体内大肠杆菌和假单胞菌的特性[35];纯木质素增加了肉鸡盲肠乳酸杆菌和双歧杆菌数量,而降低了盲肠大肠杆菌总菌数[36]。与此同时,木质素对真菌如酵母菌、白色念珠菌和皮肤毛孢子菌以及霉菌如黑曲霉菌同样具有一定抑制作用[37-38]。表2总结了木质素的抗菌活性。

表2 木质素的抗菌活性
2.2 抗氧化作用
研究表明,木质素具有惊人的高抗氧化活性,其抗氧化活性甚至能与商用抗氧化剂(愈创木酚、丁羟甲苯和阿魏酸)媲美[49]。Latif等[50]与Dong等[45]通过试验分别证实,从油棕榈叶和玉米秸秆中提取的木质素对Fe3+还原能力比香草醛更强,并且氧自由基吸收能力显著高于维生素C。木质素所显现出的抗氧化功能主要与其结构中的酚羟基、邻甲氧基、侧链羰基和羟基等特殊官能团有关,而这些官能团的抗氧化活性由强至弱依次为酚羟基>甲氧基>侧链羟基>侧链羰基[51]。这些含氧官能团与烷氧自由基发生脱氢反应以此终止氧化链式反应。如木质素中的酚羟基(ArO-H)以氢转移和单电子转移反应清除自由基(R·),可归结为ArO-H+R·→ArO·+R-H[52]。这与研究显示的木质素抗氧化能力与总酚羟基含量呈正相关而与其分子质量呈负相关结果[53]相一致。
Lu等[54]研究报道,碱木质素能分别以混合型非竞争方式和剂量依赖性方式抑制参与超氧阴离子自由基产生的黄嘌呤氧化酶活性和葡萄糖-6-磷酸脱氢酶活性;并且该试验还提出木质素的抗氧化功能可用于解释膳食纤维能降低结肠癌发生的观点。这一观点在Košíkov等[55]试验中得到证实,其体外研究显示,改性后的云杉木质素(15.2%羟基和6.5%酚羟基)和山毛榉木质素(13.7%甲氧基和4.57%酚羟基)可以显著减少人结肠癌CaCo-2细胞内由过氧化氢(H2O2)诱导的DNA链断裂;随后的体内研究还表明,在雄性大鼠对照组饲粮中添加8%用二恶烷/水提取的山毛榉木质素饲喂21 d后,可显著增强大鼠肝细胞抗氧化应激能力。表3总结了木质素的抗氧化活性。

表3 木质素的抗氧化活性
2.3 调节血糖作用
有研究发现,木质素通过延缓肠道葡萄糖吸收[62]、促进胰岛素释放以及增加胰岛B细胞数[63]等方式治疗高血糖症呈现出巨大潜力。
葡萄糖苷酶参与动物机体糖代谢,其中β-葡萄糖苷酶具有水解淀粉和纤维素以及糖苷的脱糖基化功能,而α-葡萄糖苷酶则主要促进肠道对淀粉糊精、多糖及蔗糖的分解和吸收。在医学领域,常对α-淀粉酶和α-葡萄糖苷酶进行可逆性抑制,从而延缓肠道碳水化合物吸收,降低2型糖尿病患者餐后血糖水平,以此维持患者正常血糖[64]。Berlin等[65]证实,乙醇有机溶剂溶解木质素和酶残木质素均对β-葡萄糖苷酶表现出显著抑制作用。与此同时,Chen等[66]试验研究报道,木质素以竞争性方式抑制α-葡萄糖苷酶,其抑制机理为木质素通过疏水作用和氢键与α-葡萄糖苷酶结合,形成1∶1复合物,由此改变该酶内色氨酸残基结构,导致α-螺旋含量减少。其次,Barapatre等[58]体外试验,首次证明了经化学修饰后的木质素与葡萄糖之间的亲水基团能以氢键形式结合成木质素-葡萄糖复合物,以此降低葡萄糖运动;该试验还指出木质素结构中的-OH与α-淀粉酶中的-NH、-OH以及-COOH等氢受体基在疏水作用力和氢键驱动下能形成木质素-α-淀粉酶络合物而抑制该酶活性。除此之外,Hasegawa等[67]对木质素磺酸的试验研究揭示,木质素磺酸以可逆的非竞争形式与α-葡萄糖苷酶形成复合物,降低α-葡萄糖苷酶的活性,并且在相同饮食情况下,添加5%木质素磺酸能显著增加大鼠粪便含糖量,提示木质素对肠道葡萄糖吸收的抑制作用。Xie等[68]用分子动力学初步澄清了芭蕉芋木质素对α-D-葡萄糖苷酶的抑制机制;该试验指出α-D-葡萄糖苷酶上存在着1个与木质素结合的单一位点,此结合位点与常用于治疗2型糖尿病的阿卡波糖截然不同,并且驱动木质素与α-D-葡萄糖苷酶结合的作用力主要是氢键、疏水力以及范德华力。木质素对α-D-葡萄糖苷酶的抑制效果要强于阿卡波糖,其米氏常数(Km值)和抑制常数(Ki值)分别约为0.53和0.98 μmol/L。
2.4 抗炎作用
炎症是机体应对伤害性刺激的一种防御反应,但当促炎反应与抗炎反应失衡却会引起病理性炎症反应,对机体造成二次伤害。木质素因富含多种酚类单体而具有抗炎作用,有被用于治疗和预防多种炎症相关疾病的可能性。有研究报道,水溶性木质素(water-soluble lignin,WSL)能激活红系衍生的核因子2相关因子2(Nrf2)信号通路,正向调控血红素氧合酶1的转录,从而降低胃肠道炎症和氧化应激的风险;不仅如此,WSL还能抑制核因子-κB(NF-κB)信号通路下游炎症因子的表达,降低小鼠单核巨噬细胞白血病细胞和结肠炎小鼠体内白细胞介素-6(IL-6)及肿瘤坏死因子-α(TNF-α)等促炎因子的表达水平,从而实现抗炎作用[69]。Saluja等[70]发现,用10~50 μmol/L的新型低分子质量木质素能显著抑制人肺癌细胞(Calu-3)中NF-κB萤光素酶活性,并能减少TNF-α诱导的Calu-3细胞白细胞介素-8(IL-8)释放量。Matsuhisa等[71]证明木质素磺酸能抑制由TNF-α和IFN-γ在Caco-2细胞中诱导的NF-κB p65从胞浆向细胞核移位,从而减弱NF-κB活化;同时,木质素磺酸还可降低由TNF-α和IFN-γ诱导的Caco-2细胞中促炎因子IL-1β和IL-6的表达水平。以上结果与De Melo等[72]及Do Nascimento Santos等[73]所评估的木质素抗炎作用结果相一致。
除此之外,木质素与哺乳动物木脂素结构存在相似性,富含松脂醇和落叶松树脂醇结构的木质素可经肠道菌群代谢为肠内酯(enterolactone,ENL)[4],而ENL能增加超氧化物歧化酶和过氧化氢酶的表达并降低促炎因子的释放,从而发挥抗氧化和抗炎作用[74]。
3 木质素在畜禽生产中的应用前景
现阶段木质素在动物体内的功效仍饱受争议,从营养学角度分析,木质素犹如一把“双刃剑”。一方面,木质素可作为抗营养因子,对动物生长性能无积极效应[75];另一方面,富含多种低分子质量酚类单体的木质素,却表现出益生元效应[3],对动物生产性能和肠道健康有一定积极作用[76-78](表4)。不仅如此,研究表明,从甘蔗中提取出的Ligmed-A(含90%木质素)还能有效治疗动物腹泻[79]。
从动物试验不难发现,木质素作用效果与其来源、形式、添加浓度以及动物种类紧密相关,但木质素表现出的抗菌、抗炎及抗氧化等生物学功能,在如今集约化养殖与畜禽替抗研究热潮中,却值得再度关注。除此之外,其维持血糖稳定功能在母猪生产上也值得深入研究。母猪在妊娠过程中,机体激素分泌、免疫应答及生理代谢发生改变,围产期易发生胰岛素抵抗,而过度的胰岛素抵抗能诱发妊娠期糖尿病[80]。木质素具有延缓肠道葡萄糖吸收,促进胰岛素分泌,因此可能对维持妊娠母猪餐后血糖的稳定,降低母猪胰岛素抵抗有着潜在作用价值,加之木质素能清除R·,抑制氧化酶活性,由此提高动物机体抗氧化能力,因而对缓解氧化应激对畜禽机体造成的损伤也有巨大帮助。
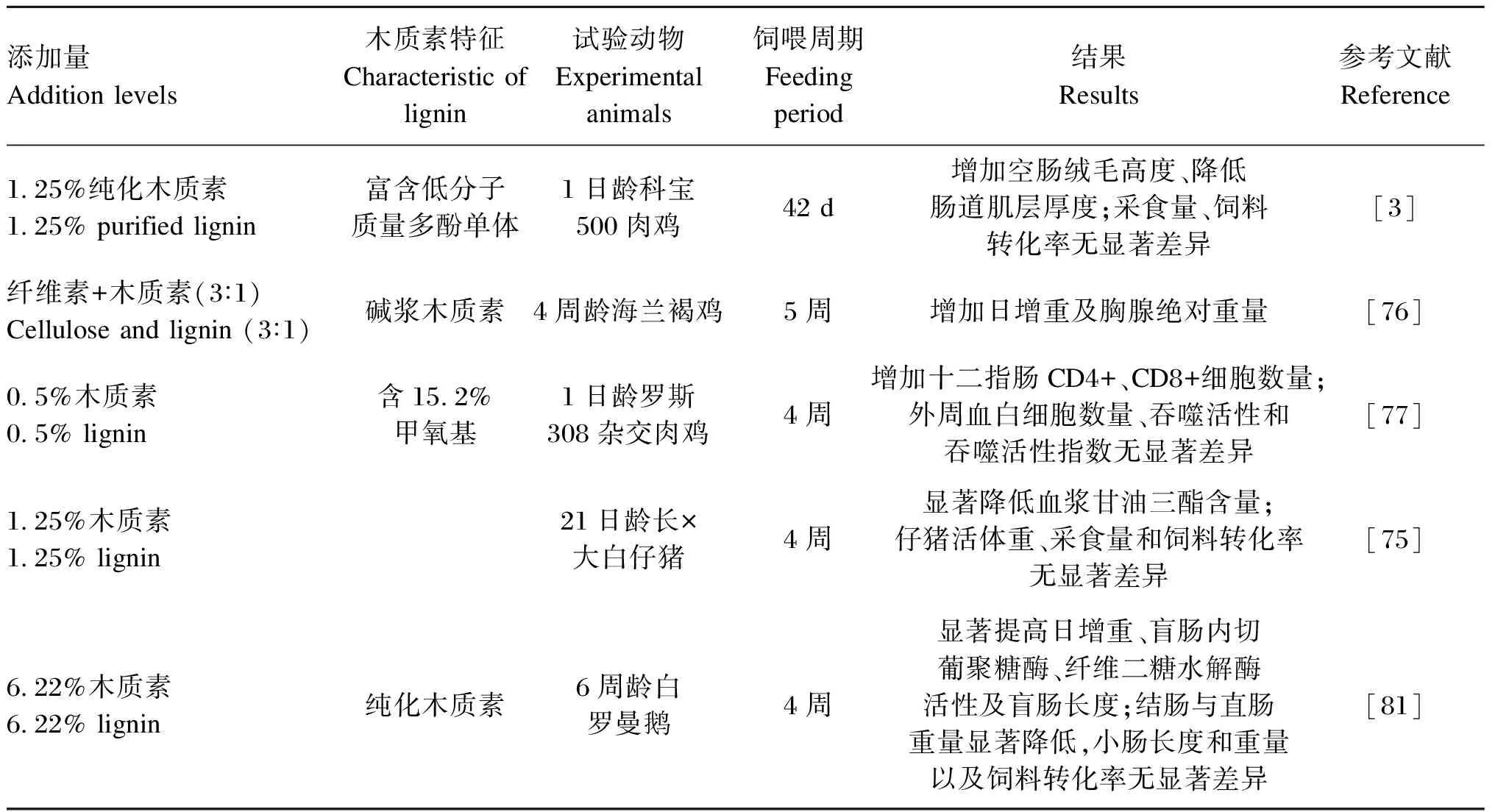
表4 木质素在畜禽生产中的部分应用研究
4 小 结
综上所述,木质素作为植物饲用功能成分,其独特的理化特性,对畜禽生长与健康表现出的良好潜力,是一种值得研究人员和生产者强烈关注的新型绿色添加剂。今后,除了就木质素在畜禽饲粮中的添加水平、组合方式及影响因素开展系统研究外,针对木质素中的结构单体与其功能之间的联系机制深入探讨,进而为揭示木质素改善肠道健康与机体代谢稳态的作用机理,指导其在畜禽饲粮中的合理应用提供科学依据,也具有十分突出的学术价值和实际意义。
