葡萄糖基水解酶基因表达与白粉病抗性的相关性研究
2022-12-08陈冠宇闫朝辉王跃进
陈冠宇,闫朝辉,王跃进
(西北农林科技大学园艺学院/旱区作物逆境生物学国家重点实验室/农业农村部西北地区园艺作物生物与种质创制重点实验室,陕西杨凌 712100)
欧亚葡萄品种(Vitis viniferaL.)是当今世界栽培范围最广、面积最大的果树之一,但易感白粉病、霜霉病、灰霉病等真菌性病害。生产中大量使用化学农药防治病害的流行,不仅增加了生产成本[1],还造成环境污染。因此,利用野生抗性材料选育抗病品种是葡萄育种的目标之一。
白藜芦醇最早从毛叶藜芦(Veratrum grandiflorumLoes.)中被分离鉴定[2],之后证明具有植保素的功能[3-4],且广泛存在于植物中[5],在葡萄中含量较高[6]。同时,白藜芦醇还具有抗癌、抗氧化、抗衰老和抗病毒作用,有益于人体健康,因而在葡萄上研究报道很多[6]。在植物体中,白藜芦醇是通过苯丙烷类代谢途径中的芪合酶合成[7],有多种存在形式,不仅包括顺反异构体,经过白藜芦醇糖基转移酶、甲氧基转移酶、漆酶、细胞色素P450酶CYP1B1催化,生成糖基化的云杉新苷、甲基化的紫檀芪、寡聚化的葡萄素和羟基化的白皮杉醇[8]。反式白藜芦醇相较于顺式白藜芦醇拥有更强的生物活性,但仍不及紫檀芪和ε-葡萄素[9-10]。高浓度的紫檀芪和葡萄素积累会对植物有害,而作为主要储存形式的云杉新苷,几乎没有抗菌作用[11]。β-糖基水解酶可以通过水解糖苷键而发挥抗病作用[12-13],欧亚葡萄品种中β-糖基水解酶Ⅰ家族共包含38个基因。
Yan等[14]在接种葡萄白粉病菌 [Uncinula necator(Schw.) Burr.]条件下,对中国野生毛葡萄丹凤-2中的38个基因进行筛选,发现VqBGH40a基因可以持续响应白粉菌的侵染,在接种后第5天达到表达峰值,进一步利用同源克隆技术分离获得了VqBGH40a,离体条件下VqBGH40a可以将植物体内不具有抗菌作用的反式云杉新苷水解为具有抗菌活性的反式白藜芦醇,从而参与到葡萄的抗病反应中。
本研究通过分析9份葡萄材料的糖基水解酶基因VqBGH40a在葡萄白粉菌诱导下的表达以及白藜芦醇含量变化,研究VqBGH40a表达是否可以增加白藜芦醇含量,以探索供试材料在接种的白粉菌诱导下,葡萄体内反式云杉新苷降解为反式白藜芦醇的次生代谢变化在抗病过程中的作用及其之间的关系。
1 材料和方法
1.1 材料情况
供试材料共9份,包括中国野生华东葡萄(V.pseudoreticulata)白河-35-1,欧亚种‘佳利酿’,二者的子代6-12-1、6-12-3、6-12-4、6-12-7;中国野生毛葡萄(V. quinquangularis)株系83-4-96,欧亚种‘粉红玫瑰’以及二者的子代‘1-9-6’。所有材料定植于陕西省咸阳市杨陵区西北农林科技大学葡萄种质资源圃内(108°4'27.95" E,34°16'56.24" N),篱壁架整形,中短梢修剪。
1.2 田间白粉病抗性评价
调查于2021年与2022年的6—8月进行。在白粉病发病感染后,根据整个叶面积上病变面积占总面积的百分比分为8个等级,再将分级结果统计,换算成感病指数[15],换算公式如下:
感病指数(%)=∑(等级值×该等级的叶片数)/(最高等级值×调查总叶片数)×100
根据感病程度将感病指数分为5个等级:1级,未感病或免疫;2级,高抗;3级,抗病;4级,感病;5级,高感。对应的感病指数分别为:0、0.1%~5.0%、5.1%~25.0%、25.1%~50.0%、50.1%~100.0%。
1.3 田间葡萄人工接种白粉菌
采用湿压片法对叶片接种白粉菌[16]。采摘田间布满新鲜白粉菌孢子的葡萄叶片,压在预先使用无菌水喷雾且无明显露点的健康幼叶上3~5 s,然后分开,侧面观察叶片可见明显孢子后套袋,对照组则使用健康叶片压片。在接种后0、1、4、7 d采集人工接种白粉菌处理(PM)与对照组(MOCK)的叶片,立即用锡箔纸包裹放入液氮中速冻,放置于﹣80 ℃超低温冰箱中保存待用。
1.4 总RNA提取与实时荧光定量PCR
总RNA提取参考Plant RNA Kit说明书(Omega R6827)。反转录合成cDNA方法参照Evo M-MLVRT Mix Kit with gDNA Clean for qPCR说明书(Accurate Biotechnology AG11728)。按照SYBR® Green PremixPro TaqHS qPCR Kit(Accurate Biotechnology AG11701)说明书配置qPCR反应体系。使用实时荧光定量PCR(Applied Biosystems Step One Plus)与2﹣∆∆CT法[17]分析BGH40a基因相对表达量[14],以葡萄GAPDH(XM_002278316.4)作为内参基因(表1),引物由杨凌天润奥科生物科技有限公司合成。设置3个生物学重复并进行3次技术重复。

表1 试验所用引物Table 1 Primer sequences used in this experiment
1.5 芪类物质的提取与测定
葡萄叶片经真空冷冻干燥机(Christ Alpha 2-4 LSC basic)干燥50 h后,加入色谱级甲醇(1∶10,W∶V)4 ℃避光条件过夜静置抽提,取上清液使用0.22 μm滤膜过滤待测[18]。
反式云杉新苷使用SunFire® C18色谱柱(4.6 mm×250 mm,5 μm)分离,Waters Acquity Arc检测。流动相A为0.1%甲酸水溶液,流动相B为色谱级乙腈。洗脱程序为0 min,80%A;0.7 min,80%A;20 min,25%A;22 min,0%A;24 min,80%A;30min,80%A。流速为1.5 mL·min-1,检测波长306 nm,柱温40 ℃,进样量为10 μL[19]。
反式白藜芦醇使用ZORBAX RRHD Eclipse Plus C18色谱柱(3 mm×50 mm,1.8 μm)分离,Agilent 1290 Infinity Ⅱ-6470检测,采用电喷雾离子源(ESI),多反应监测离子扫描模式(MRM)。流动相与反式云杉新苷检测相同。洗脱程序为:0 min,80%A;0.8 min,80%A;1.5 min,20%A;2.5 min,80%A,3.5 min,80%A。柱温40 ℃,流速0.3 mL·min-1,上样量为2 μL。母离子227.24,子离子226.7,碎裂电压130 V,碰撞能量10 eV[20]。使用反式云杉新苷和反式白藜芦醇标准品(上海源叶)确认保留时间,根据保留时间与峰面积进行定性和定量。
1.6 数据分析
使用Excel对数据进行统计分析和图表绘制。使用IBM SPSS 26对数据进行多重比较,采用单因素ANOVA检验LSD方法(*p<0.05; **p<0.01)。使用IBM SPSS 26对数据进行皮尔逊相关性分析(*p<0.05)。
2 结果与分析
2.1 自然条件下田间抗病性鉴定
自然条件下,所有材料的感病指数及抗病性鉴定结果见表2。其中,4个材料表现抗病,包括白河-35-1、6-12-1、83-4-96、‘1-9-6’,感病指数介于19.57%~24.60%,被划分为抗病3级,其余均表现为感病,感病指数为31.67%~48.29%。华东葡萄白河-35-1感病指数最低,为19.57%,表现为抗病;而‘佳利酿’的感病指数最高,为48.29%,表现为感病,二者的4个子代感病指数介于双亲之间,呈现为亲中值,其中6-12-1表现出抗病性状,而6-12-3、6-12-7和6-12-4表现型为感病。83-4-96和‘粉红玫瑰’杂交组合中,子代‘1-9-6’的感病指数是23.14%,属于抗病类型,与其亲本83-4-96相似。
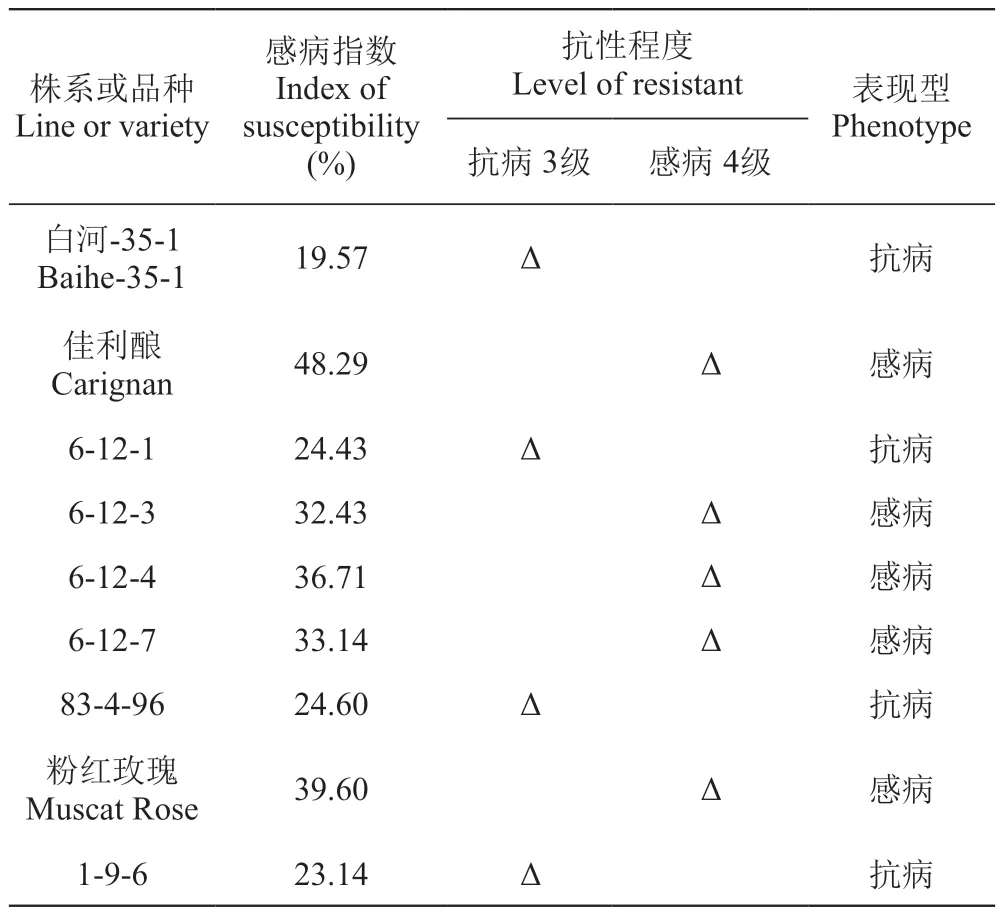
表2 供试葡萄材料叶片田间条件下白粉病抗性鉴定Table 2 Identification of the resistance to Uncinula necator of grapevine leaves under field conditions
2.2 葡萄材料中BGH40a基因差异性表达
利用qRT-PCR分析供试9个葡萄材料中VqBGH40a在白粉菌诱导下的差异表达。结果表明,VqBGH40a在不同材料中表达不同,总体表现出抗病材料中的表达上调趋势(图1、图2)。其中,6-12-3、6-12-4、83-4-96中存在上调表达,特别是83-4-96在接种白粉菌后4 d上调17.7倍,其次是6-12-4接种白粉菌后7 d上调11.9倍。在‘粉红玫瑰’‘1-9-6’、白河-35-1、‘佳利酿’、6-12-1、6-12-7接种白粉菌后0、1、4、7 d均为下调表达。

图1 杂交组合‘白河-35-1×佳利酿’及其子代BGH40a在白粉菌诱导下差异表达Figure 1 Differential expression analysis of the BGH40a from cross combination 'Baihe-35-1×Carigane' and their progeny using artificial inoculation by Uncinula necator

图2 杂交组合‘83-4-96×粉红玫瑰’及其子代BGH40a在白粉菌诱导下差异表达Figure 2 Differential expression analysis of the BGH40a from cross combination '83-4-96×Muscat Rose' and their progeny using artificial inoculation by Uncinula necator
2.3 BGH40a基因差异性表达的芪类物质含量变化
在接种白粉菌条件下,伴随着VqBGH40a在白粉菌诱导下的差异表达,供试材料叶片中反式云杉新苷的降解与反式白藜芦醇含量增加的变化趋势见表3、表4。从总体上,在接种白粉菌后随着时间推延,出现糖基水解酶对反式云杉新苷降解的趋势,而对应的反式白藜芦醇含量表现出不同程度的增加,即一减一增相反的变化趋势。接种白粉病菌处理下,毛葡萄83-4-96的后代‘1-9-6’中反式白藜芦醇含量也是在减少,出现这种现象可能是‘1-9-6’的反式云杉新苷降解后的产物不仅是白藜芦醇一种物质,而且有葡萄素增加(研究结果未展现),这是下一步需要探索的问题。感病葡萄中,反式云杉新苷含量下降,特别是‘佳利酿’中出现大幅下降,但反式白藜芦醇含量没有出现增加趋势,这个结果可能是‘佳利酿’的糖基水解酶基因功能差异导致。抗病葡萄中除白河-35-1外,其他材料的糖基水解酶基因较好的表达,使得反式云杉新苷含量下降的同时,反式白藜芦醇含量上升到了较高的水平。
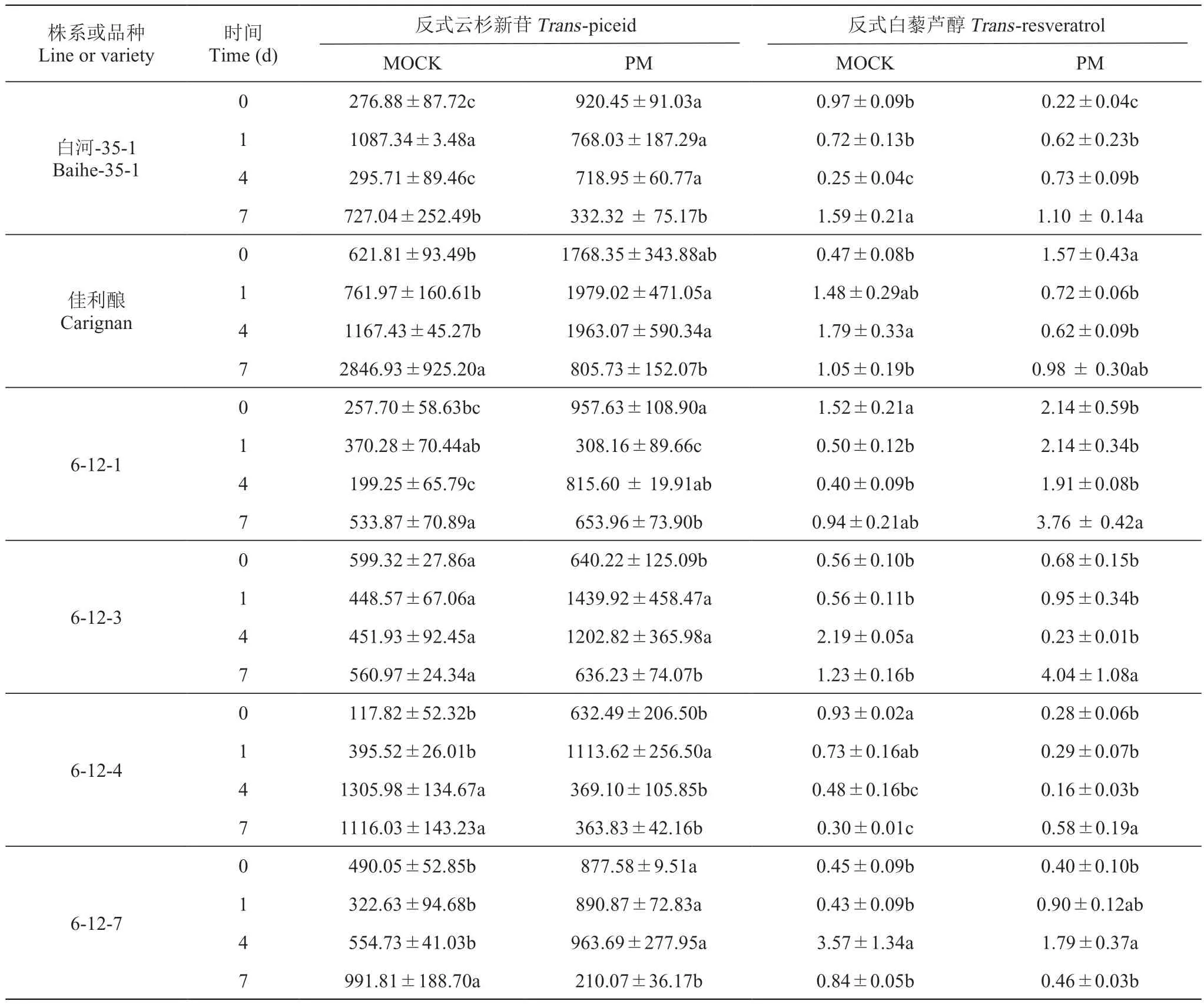
表3 杂交组合‘白河-35-1×佳利酿’及其子代在接种白粉菌条件下BGH40a表达过程中反式云杉新苷和反式白藜芦醇含量变化Table 3 Changes of the content of trans-piceid and trans-resveratrol in the process of BGH40a expression from cross combination'Baihe-35-1×Carignan' and their progeny using artificial inoculation by Uncinula necator μg·g-1 DW

表4 ‘83-4-96×粉红玫瑰’及其子代接种白粉菌BGH40a表达过程中反式云杉新苷与反式白藜芦醇含量变化Table 4 Changes in the content of trans-piceid and trans-resveratrol in the process of BGH40a expression from cross combination'83-4-96 ×Muscat Rose' and their progeny using artificial inoculation by Uncinula necator μg·g-1 DW
2.4 葡萄材料的感病指数与云杉新苷变化量的相关性
对两个葡萄杂交组合及后代的感病指数与芪类物质含量的相关性进行分析。其中‘白河-35-1×佳利酿’杂交组合中感病指数与反式云杉新苷含量呈现出正相关(0.561),与反式白藜芦醇含量呈现出负相关(﹣0.197);反式云杉新苷和反式白藜芦醇之间的相关性为﹣0.114。而在‘83-4-96×粉红玫瑰’杂交组合中,反式云杉新苷与抗病指数相关性为﹣0.006,反式白藜芦醇与感病指数相关性为﹣0.856,但是反式云杉新苷与反式白藜芦醇相关性为0.324(表5、表6)。

表5 ‘白河-35-1×佳利酿’及其子代感病指数和芪类物质含量相关性Table 5 Correlation analysis of index of susceptibility and contents of stilbene in cross combination'Baihe-35-1×Carignan' and their progeny

表6 ‘83-4-96×粉红玫瑰’及其子代感病指数和芪类物质含量相关性Table 6 Correlation analysis of index of susceptibility and contents of stilbene in cross combination'83-4-96×Muscat Rose' and their progeny
3 讨论与结论
糖基水解酶广泛分布于所有植物体中,通过水解葡萄糖苷在生物代谢过程中发挥作用[21]。目前,已在多种植物中报道了糖基水解酶的功能,包括拟南芥[22]、水稻[13]、玉米[12]、棉花[23]、葡萄[14]等。例如,水稻中的37个β-糖基水解酶基因大多数在RS105菌株侵染后2 h表达升高,24 h或48 h达到峰值[13];棉花中GrGLU42和GrGLU43在接种黄萎病菌96 h表达量达到峰值[24];而葡萄中38个VqBGH基因大多数在接种白粉菌后48 h或120 h表达出现峰值[14]。
本研究的两个葡萄杂交组合的9个材料,母本均为中国野生抗病材料,父本均为欧亚种感病材料,在白粉菌侵染0、1、4、7 d的糖基水解酶BGH40a基因表达中,只有3个材料表现上调,其中感病材料6-12-3、6-12-4也表现上调,说明糖基水解酶BGH40a基因的表达与材料自身特性有一定的关系,由于样本量不足与采样时间间隔长而未能表现出与抗病性之间的具体关系。在芪类物质含量变化的测定中,中国野生毛葡萄83-4-96在接种白粉菌的第4天VqBGH40a表达上调,且反式云杉新苷含量减少,与Yan等[14]在中国野生毛葡萄丹凤-2中的试验结果相一致;‘1-9-6’中反式云杉新苷降解后,反式白藜芦醇表现出减少的趋势,这可能是反式云杉新苷降解后的产物转化为其他抗菌性更强的衍生物,如葡萄素、紫檀芪等,因而表现出抗病性状[23]。抗病葡萄除白河-35-1外,虽然反式云杉新苷含量减少的量不明显,但是反式白藜芦醇含量保持较高的水平。感病葡萄中,反式云杉新苷含量下降,特别是‘佳利酿’中含量大幅下降,但反式白藜芦醇含量增加较少,这个现象与不同葡萄材料中基因表达与功能差异有关,将在下一步进行研究。不同葡萄材料在白粉病菌诱导下,反式云杉新苷含量下降与反式白藜芦醇增加的趋势说明,葡萄中糖基水解酶BGH40a在白粉病菌侵染过程中,具有水解反式云杉新苷形成反式白藜芦醇的作用,也就是说,这种糖基水解酶在葡萄植物体内的次生代谢对葡萄抗病性有着一定的作用。
初步分析发现,中国野生毛葡萄83-4-96与欧亚葡萄品种‘粉红玫瑰’的杂交组合中,叶片中糖基水解酶BGH40a在白粉病菌侵染过程中,具有水解反式云杉新苷的作用;而且杂交组合‘83-4-96×粉红玫瑰’及其子代感病指数和反式白藜芦醇表现出较高的负相关性,相关系数是﹣0.856,表现出增强抗病性的趋势。同时,在广西栽培多年的‘1-9-6’表现出抗病性,也支持本研究结果[25-26]。而‘白河-35-1×佳利酿’及其子代的感病指数和反式白藜芦醇的相关性较低,因此有必要对更多野生种质及其子代做进一步研究以证实上述观点。
综上所述,在供试的9份葡萄材料中,有4份表现抗病。抗病葡萄中BGH40a基因在白粉菌诱导下表达,可以将反式云杉新苷水解降解为反式白藜芦醇,增加了反式白藜芦醇含量,使葡萄表现出抗病性,而且这种糖基水解酶的表达特性可以通过杂交育种遗传给后代,这为今后选择糖基水解酶基因表达特性强的葡萄种质用作抗病育种的亲本,提供了科学依据。
