不同小生境下典型喀斯特森林植物叶片功能性状特征
2022-12-03周汀崔迎春叶雨艳赵文君侯贻菊吴鹏丁访军
周汀,崔迎春,叶雨艳,赵文君,侯贻菊,吴鹏,丁访军
(1.贵州大学 生命科学学院,贵州 贵阳 550025;2.贵州省林业科学研究院,贵州 贵阳 550005)
喀斯特区地表呈现大量碳酸岩裸露的特殊地貌特征,加上剧烈的岩溶作用以及土壤空间分布的异质性,导致喀斯特区小生境空间分异较大,并且小生境中温度、水分、光照等生态因子复杂多样,致使其土壤及小气候环境高度变异[1-3]。由于特殊的地理构造和水热条件,喀斯特区植物的生长发育通常受到环境因子的制约,为适应复杂多变的生存环境,植物自身从生理结构和生化过程中产生一系列变化,进而形成不同的生存策略[4]。同时,生境高度的异质性也是维持该地区生态系统物种多样性的重要因素[5]。
植物叶片作为植物的主要功能器官,其功能性状是植物在适应生存环境时选择生态策略和生存能力的主要表征之一[6],并在植物碳同化及水热条件适应机制等方面起着重要作用[7]。针对不同区域、不同尺度下植物叶片的特性及其对环境变化的响应机制等方面开展了大量研究[8-11],其结果也均表明植物叶片的功能性状都会随着环境的不同而产生差异。但是对典型的喀斯特森林生态系统,由于小生境的高度异质性,植物在如此严酷的生境下采取怎样的生存策略却不得而知。基于此,本研究从植物叶片功能性状的角度出发,以10 种典型喀斯特森林植物为对象,研究其对不同小生境(土面、石面及石沟)下植物的叶片功能性状及种间变异特征,并以资源获取利用关系与适应策略选择为基础,分析探讨了该地区植物对生境适应的生理响应,阐明了不同小生境下植物的生存策略,旨在为喀斯特区生态恢复与治理提供科学依据。
1 研究方法
1.1 研究区概况
研究区位于贵州茂兰国家级自然保护区、荔波喀斯特森林生态系统定位观测研究站,地理位置107°52′10′′~108°05′40′′E、25°09′20′′~25°20′50′′N。地势呈西北高东南低,最高海拔1 078.6 m,最低为430.0 m,平均为550~850 m。气候属中亚热带季风湿润气候区;年平均气温15.3℃,其中1 月份5.2℃,7 月份23.5℃;年均降水量1 752.5 mm,多分布于4—10 月;年平均湿度83.0%;年日照时数1 272.8 h,无霜期315 d,太阳年辐射总量为63 289.80 kW·m-2,≥10℃积温4 598.6℃[12]。
1.2 试验设计
1.2.1 植物叶片采集与处理
在茂兰喀斯特森林生态系统定位观测研究站原有的固定样地中,按照植物根部着生的不同小生境类型(土面、石面、石沟)选择了10 种典型的喀斯特森林植物(常绿乔木(AL)3 种、落叶乔木(DT)3 种及常绿灌木(ES)4 种),分别记录其物种名、胸径/地径(cm)、树高(m)、海拔(m)、坡度、坡位、根部小生境类型及经纬度等(表1)。
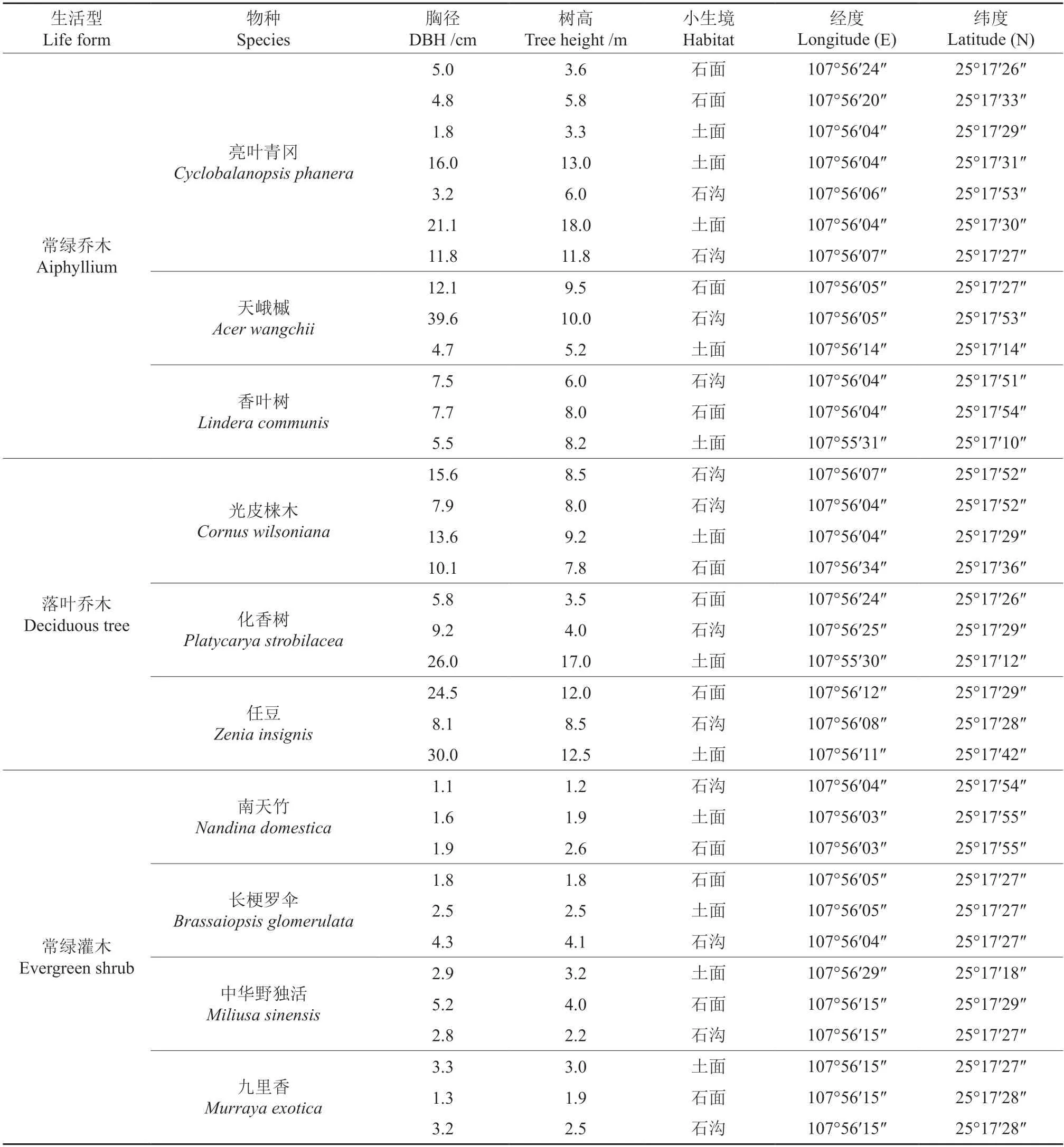
表1 典型喀斯特地区常见植物生境概况Table 1 Characteristics of common plant habitats in typical karst areas
用高枝剪剪取每个样株冠层东南西北和上中下不同部位的枝条,采摘每个枝条上的叶片(5~7 片),必须是完全伸展、无病害、完整的叶片(均不带叶柄),每个样株采集叶片60~80 片,充分混合均匀后,采用四分法,密封在塑封口袋内并置于便携式冷藏箱中备用;每个样株挑选5 片完整叶片使用分析天平(精度0.000 1 g)称其鲜质量(g),使用游标卡尺(精度0.01 mm)沿主叶脉选取4 点(避开叶脉)测量叶片厚度(mm),之后将叶片于蒸馏水中暗光环境下浸泡24 h 后,迅速取出并用吸水纸吸去叶片表面的水分,用电子天平分别称取其饱和鲜质量(g);然后利用便携式叶面积仪(LI-3100,America)分别对其叶面积(LA,cm2)进行测定;最后将叶片放入烘箱中于65℃下烘干至恒质量(72 h),再用电子天平分别称其干质量(g)。
1.2.2 植物叶片光合参数测定
选择晴朗天气,在上午9:00—11:00使用Li-6400(Li-CorInc.,USA)便携式光合作用测定系统,对样株冠层上部选取充分展开、成熟、向阳的叶片进行测定,测定前对植物叶片使用光强为1200μmol·m-2·s-1的红蓝光源诱导15~20min,测定时红蓝光叶室光强设置为1 200 μmol·m-2·s-1[13];空气流速500 cm3·min-1;叶室温度26±2℃;CO2浓度400±10 μmol·mol-1。每株植物间隔1 h 整点测定叶片的瞬时光合指标(净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)),每次测定1 片,每片重复测定3 次。针对个别植株高大的树种,采用离体测定的方法对其光合参数进行测定,即:将离体枝条或复叶离体后迅速插入水中、环剥去除切口处3 cm 左右的韧皮部并除去剩余叶片后立即开展相关光合指标的测定[14]。
1.2.3 相关叶片指标计算方法
各叶片指标计算方法如下[15]:
比叶面积(SLA)=叶面积(LA)/叶片干质量;
叶干物质含量(LDMC)=叶片干质量/叶片饱和鲜质量;
叶片含水率(LWC)=(叶片鲜质量-叶片干质量)/叶片鲜质量;
叶组织密度(LTD)=叶片干质量/(叶面积(LA)×叶厚度(LT));
水分利用效率(WUE)=净光合速率(Pn)/蒸腾速率(Tr);
羧化效率(CE)=净光合速率(Pn)/胞间CO2浓度(Ci)。
1.2.4 小生境类型划分
结合前人研究成果和本试验设立研究区的实际情况,将小生境类型区分为土面、石面、石沟3个类型[16]:
土面:基岩裸露率小于50%,并以土体为主,没有形成岩石凹槽或凹沟深度超过30 cm 的小生境,土面的土层较厚,通风条件较好,水分散失较慢。
石面:基岩裸露率大于50%,连片土壤覆盖面积小于1 m2,形成的岩石凹槽或凹沟深度不超过30 cm,土壤层厚度不超过20 cm,石面的通风条件好,但水分散失快,保水保肥能力弱,短期内极易形成频繁的临时性干旱,其生境条件极为严酷。
石沟:基岩裸露率大于50%,连片土壤覆盖面积小于1 m2,形成的岩石凹槽或凹沟深度不超过30 cm,土壤层厚度超过20 cm;或虽基岩裸露率小于50%,以土体为主,且形成了岩石凹槽或凹沟深度超过30 cm 的小生境,石沟里面一般土层较厚,不易失水,保水保肥能力较好,不易受短期干旱的影响,其生境条件相对较好。
1.3 数据处理
采用Excel 2019 和SPSS 21.0 软件对数据进行统计分析、主成分分析及Pearson 相关性检验;采用Origin 8.1 软件进行绘图。数据进行相关分析、One-way ANOVA 分析以及T 检验时,为满足正态分布要求和ANOVA 假设,需先将各类数据转换为自然对数形式,即:ln (x+1);在进行多重比较时,首先利用Levene’s test 对方差进行齐性检验,基于其检验结果,齐性(或非齐性)采用LSD 法(或Tamhane’s T2 检验法)进行多重比较[12]。
2 结果与分析
2.1 不同小生境下植物叶片的结构型性状
不同生活型植物叶片的结构型性状在各小生境中基本上都具有较一致的变化趋势(图1)。在土面小生境中,叶密度(LTD)和叶干物质含量(LDMC)呈现出常绿乔木(AL)>落叶乔木(DT)>常绿灌木(ES),而比叶面积(SLA)和叶片含水率(LWC)则相反,并且之间差异显著(P<0.05),叶厚度(LT)和叶面积(LA)在不同生活型植物间无显著性差异(P>0.05);在石面与石沟小生境中,常绿灌木的LT 较常绿乔木和落叶乔木偏低且差异显著(P<0.05),常绿乔木的LTD 较常绿灌木和落叶乔木偏高且差异显著(P<0.05),LWC 和SLA 呈常绿灌木>落叶乔木>常绿乔木,LDMC 反之,并且差异显著(P<0.05)。相同生活型植物叶片结构型性状在各小生境间存在一定程度的差异,常绿乔木在土面小生境中的LDMC、LT 显著低于石面与石沟(P<0.05),而LWC 则相反;落叶乔木在土面生境中的SLA 显著高于石面与石沟(P<0.05),而LT则反之;常绿灌木在3 个小生境间均无显著性差异(P>0.05)。
在不同小生境中,各叶片指标都存在不同程度的种间变异(图1),其中变异系数最大的为石沟生境中LA(96.09%),变异系数最小的为土面生境中LWC(14.89%)。在土面生境中,各叶片指标间变异大小为LA >SLA >LDMC >LT >LTD >LWC;在石面生境中,叶片指标变异大小为LA >SLA >LTD >LT >LDMC >LWC;在石沟生境中,叶片指标变异大小为LA >SLA >LT >LTD >LDMC >LWC。在不同生境之间,植物叶片的LDMC、SLA、LTD、LT、LWC 变异大小均为石面>石沟>土面,LA 为石沟>土面>石面。

图1 不同小生境下植物叶片的结构型性状Fig.1 Structural traits of plant leaves in different niches
在小生境水平上(图1),石面小生境植物叶片的LA 显著低于石沟小生境(P<0.05),土面小生境植物叶片的LT 显著低于石面与石沟小生境(P<0.05),其余指标均无显著性差异(P>0.05)。但从性状值大小上看,LA 为石面<土面<石沟;LT、LDMC 为土面<石面<石沟;SLA、LTD 为石沟<石面<土面;LWC 为石面<石沟<土面。对不同小生境下植物叶片的性状特征进行相关性分析(表2),在3 个小生境中,植物叶片的LDMC 与LTD,SLA 与LWC 均为极显著正相关(P<0.01),LDMC 与SLA、LWC,LT 与SLA,SLA 与LTD,LTD 与LWC 均呈现极显著负相关(P<0.01)。在石面和石沟生境中,LDMC 与LT 呈极显著正相关(P<0.01);LT与LWC 呈极显著负相关(P<0.01)。在土面生境中SLA 与LA 呈极显著负相关(P<0.01)。这表明植物因叶片结构型性状特征的不同而使用不同的生存策略来适应所处生境。

表2 不同生境下各叶片结构型性状间的相关性†Table 2 Correlation among leaf structural traits in different habitats
2.2 不同小生境下植物叶片的功能型性状
不同生活型植物在不同小生境中各功能型性状表现存在一定差异(图2),落叶乔木的净光合速率(Pn)在各小生境中均显著高于其他生活型植物(P<0.05),并且其羧化效率(CE)均显著高于常绿灌木(P<0.05);在土面小生境中,落叶乔木的气孔导度(Gs)显著高于常绿乔木(P<0.05),常绿灌木的胞间CO2浓度(Ci)显著高于常绿乔木与落叶乔木(P<0.05);在石面小生境中,常绿灌木的Ci显著高于常绿乔木(P<0.05),落叶乔木的蒸腾速率(Tr)显著高于常绿乔木和常绿灌木(P<0.05),常绿乔木的水分利用效率(WUE)显著高于落叶乔木(P<0.05);石沟小生境中,落叶乔木的气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)显著高于常绿乔木和常绿灌木(P<0.05),WUE 则与之相反。相同生活型植物在小生境之间表现也各有不同,常绿乔木在石沟小生境中Ci、WUE 显著高于土面与石面(P<0.05);落叶乔木在土面小生境中WUE显著高于石面与石沟(P<0.05),而Ci则表现为石沟小生境要显著高于土面与石面(P<0.05);常绿灌木在各小生境之间均无显著性差异(P>0.05)。
不同小生境下,各植物叶片功能型指标均存在不同程度的种间变异(图2),其中最大为石沟生境中的Gs(167.35%),最小为石沟生境中的Ci(13.68%)。在土面生境中,各功能型性状间变异大小表现为Gs>CE >Pn>Tr>WUE >Ci;石面生境中,表现为Gs>Tr>Pn>CE >WUE >Ci;石沟生境中表现为Gs>Tr>Pn>WUE >CE >Ci。在不同小生境间,Gs、Tr、WUE 变异大小均表现为石沟>石面>土面,Pn、Ci、CE 变异大小均表现为土面>石面>石沟。
对不同小生境间植物功能型性状指标进行比较发现(图2),其中Ci在土面生境与石沟生境之间存在显著差异(P<0.05);Gs、Pn、Tr、WUE和CE 在小生境之间差异不显著(P>0.05)。从性状值大小来看,Gs、Pn及Ci均呈石沟>石面>土面;Tr为石面>石沟>土面;WUE 为石沟>土面>石面;CE 为土面>石面>石沟。虽然小生境之间性状值整体差异不显著,但其大小表现存在不同,说明植物对不同小生境的响应有所不同。对植物叶片功能型性状指标进行相关性分析(表3),发现各指标之间存在一定关联性。在3 个小生境中,Pn与Gs、Tr、CE 都呈极显著正相关(P<0.01);Gs与Ci、Tr均呈极显著正相关(P<0.01);CE与Tr均呈极显著正相关(P<0.01)。在土面和石沟生境中,Gs与CE 呈极显著正相关(P<0.01);WUE与Tr在石面和石沟生境中呈极显著负相关(P<0.01)。

表3 不同生境下各叶片功能型性状间的相关性Table 3 Correlation of leaf functional traits of plants in different habitats

图2 不同小生境下植物叶片的功能型性状Fig.2 Functional traits of plant leaves in different niches
2.3 不同小生境植物叶片功能性状的相关关系
Pearson 相关性检验结果(表4)表明,在土面生境中,Gs与LWC 极显著正相关(P<0.01),与LDMC 极显著负相关(P<0.01);Ci与SLA、LWC 极显著正相关(P<0.01),与LTD、LDMC极显著负相关(P<0.01);在石面生境中,Ci与SLA 呈极显著正相关(P<0.01),与LDMC呈极显著负相关(P<0.01),其余性状指标均无极显著相关性(P>0.01)。在植物叶片功能性状的主成分分析中(图3),不同小生境下结构型性状指标与功能型性状指标存在不同的表现,土面生境中,PC1 轴与PC2 轴的总解释度为73.3%,PC1 与LA、LWC、Tr、Gs、CE 呈正相关关系,与LTD、Ci为呈相关关系,PC2 与LT、LDMC呈正相关关系,与WUE、SLA、Pn呈负相关关系;石面生境中,PC1 轴与PC2 轴的总解释度为64%,PC1与Ci、SLA、LWC、Gs呈正相关关系,与LTD、LDMC 呈负相关关系,PC2 与CE、Pn呈正相关关系,与LA 呈负相关关系;石沟生境中,PC1 轴与PC2 轴的总解释度为66%,PC1与Ci、Gs呈正相关关系,与WUE、LTD 呈负相关关系,PC2 与LT 呈正相关关系,与LA、SLA 呈负相关关系。

表4 不同生境下植物叶片功能性状的相关性Table 4 Correlation of leaf functional traits of plants in different habitats
从植物生活型水平来看(图3),常绿乔木在3 个生境中都位于PC1 轴负向区域,具有较高LDMC、LTD,并在石面和土面生境下具有较高的WUE、LT;落叶乔木在土面生境中位于PC2 轴负向区域,具有较高的WUE、Ci。在石面和石沟生境中,落叶乔木分布于PC2 轴正向区域,都具有较高的WUE、LT,其中任豆与光皮梾木、化香树的欧氏距离较远,说明落叶乔木在石面和石沟小生境中可能存在性状分异较大的情况。常绿灌木在3 个生境中都基本分布于PC2 轴负向区域,都具有较高的SLA、LWC。
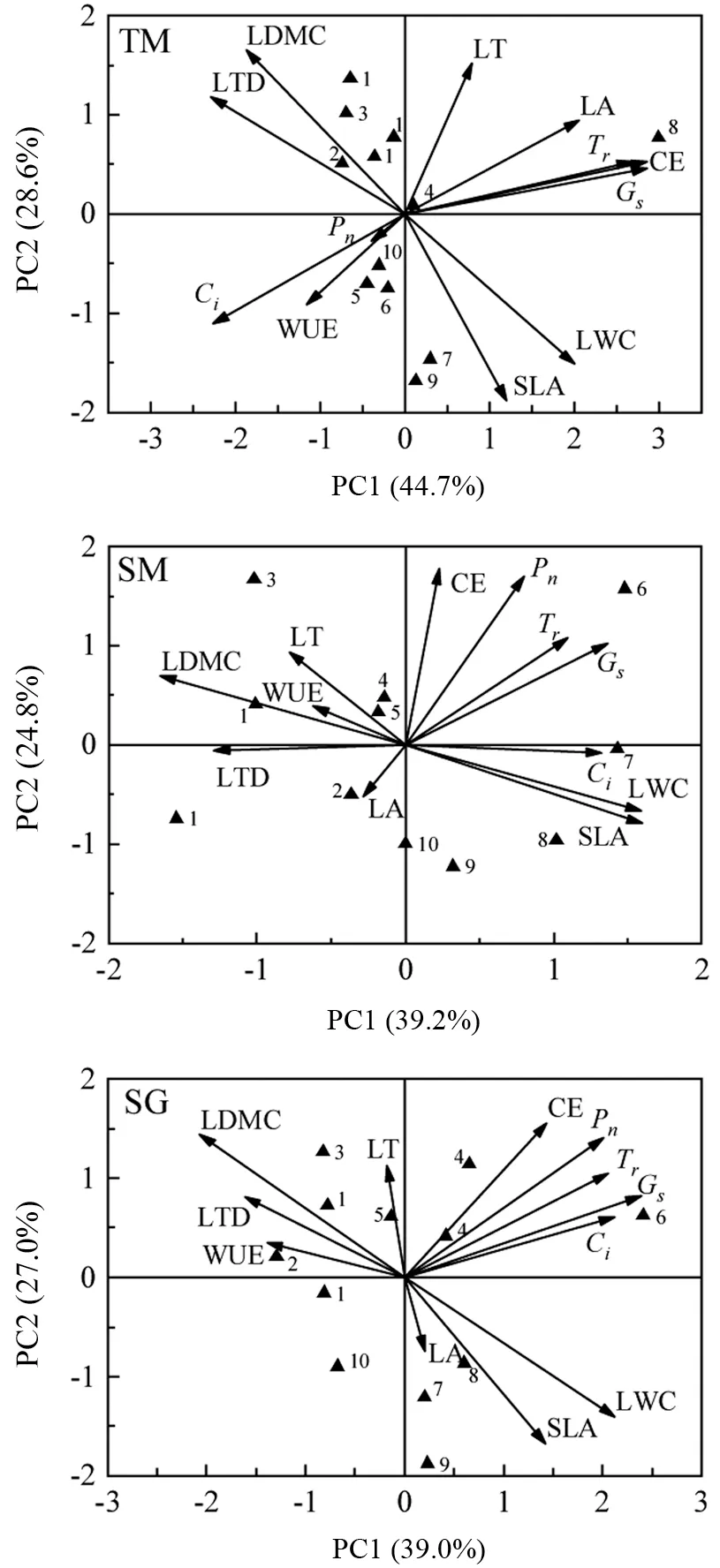
图3 不同小生境主成分分析Fig.3 Principal component analysis of different niches
3 讨论
3.1 植物叶片功能性状对小生境的响应
叶片是植物对环境适应表现的主要器官[17]。不同分布区内,植物受到不同环境因子的影响,进而产生较大差异[18-19]。在本研究中,叶干物质含量、比叶面积、叶密度、叶厚度和叶片含水率的种间变异均在石面小生境中最高,土面小生境最低,表明当地植物在光、温、水等分布高度异质性的环境中,通过程度较高的叶片形态表征变异来对生境做出响应。同时叶面积表征植物捕光及固碳能力[20],在3 个生境中叶面积种间变异大小均为最高,该结果与之前研究结果[21]基本一致,说明植物生长在受到外界环境干扰时,叶面积与生境异质性的匹配度较高,能准确反应生境间的差异。
诸多研究显示在喀斯特区小生境严苛程度为石面最大,石沟与土面相当[22-24],植物叶干物质含量越高对环境抗逆性越强,能够在资源匮乏的环境中更好地生存。而比叶面积与植物的环境资源利用率有关[25],从研究数据来看(图1),石沟小生境植物叶干物质含量大于其他小生境,而比叶面积和叶密度则相反,这可能是由于石沟小生境中土壤水分含量和温度相对较高[26],受热后不易散失,水热条件波动较稳定[24],使得石沟小生境适宜植物生长,能够蓄积更多的生物量。而在土面小生境中,叶干物质含量、比叶面积与叶密度的性状表现与石沟小生境相反,这可能是由于土面小生境上层土壤吸收热量容易向下层土壤扩散[24],使其表层土壤温度在除冬季外要低于其他小生境[26],同时土面生境相对开放,土层较厚,表层土壤水分留存相对要低于其他小生境[27]。石面小生境水热条件波动剧烈,土壤温差较大,持水能力较差[26],导致植物叶片含水率较低,叶面积较小,通过减少水分散失降低自身结构失水或高温损伤风险。在本研究中发现,石面小生境中植物叶干物质含量、叶厚度大于土面小生境,比叶面积、叶密度小于土面小生境,这可能与喀斯特森林群落中有丰富的林下持水,且石面小生境土壤对水分响应较快[26],能够快速聚集水分有关。
不同生活型植物对不同小生境中的环境响应基本一致,常绿乔木的叶干物质含量和叶密度较高,但叶片含水率和比叶面积较低,而常绿灌木与之相反,落叶乔木则居中。落叶乔木的光合能力在各小生境中均高于其他生活型植物,而常绿乔木与常绿灌木在光合能力上差异不明显。不同生活型植物在小生境间的差异不显著,这可能是由于植物自身遗传因素的影响。
光合作用是植物生长及生物量蓄积的重要基础,其受到植物自身生理和环境因素的共同影响[28]。干旱胁迫能够导致植物叶片气孔关闭限制光合作用,严重的情况下能够破坏植物光合结构,导致光合作用严重下降[29]。胞间CO2浓度作为光合作用的物质来源,受到大气CO2浓度、植物叶片气孔开闭大小及CO2在叶片内部扩散阻力的共同影响[30]。研究结果显示(图2),在小生境水平,石沟小生境植物叶片的胞间CO2浓度显著大于土面生境(P<0.05),其余性状值均差异不显著(P>0.05),但大小存在差别。植物叶片气孔导度、胞间CO2浓度、净光合速率均为石沟>石面>土面,这可能是由于不同小生境中土壤水分含量差异导致植物受到不同程度的干旱胁迫,影响气孔开闭大小并进一步影响植物净光合速率。由于石面小生境中水热条件不稳定[26],植物为避免高温损伤,同时维持光合作用,所以具有较高的蒸腾速率和较低的水分利用效率。
3.2 植物叶片对小生境适应的策略选择
在植物长期对环境适应过程中,植物性状受到生理、遗传、环境等多因素的影响,各性状之间存在一定的相互作用关系进而形成适应复杂环境的性状组合[31-32]。从研究结果来看,3 种小生境中,比叶面积、叶片含水率与叶干物质含量、叶密度均呈现极显著负相关(P<0.01),印证了植物叶干物质含量增大,叶片含水率降低,使得叶密度增大,进一步导致比叶面积降低,这样的性状组合普遍存在于植物群落中[33]。
有研究表明,当土面小生境的表层土壤失水时,深层土壤水或表层岩溶带地下水能够稳定补足[27],同时土壤有机质含量高,热容量和导热率大,吸收的热量容易向下层传递,失去热量时容易补充[24]。所以植物在土面小生境中资源供给相对稳定,因此表现为较低的叶干物质含量和叶厚度及较高的叶密度、比叶面积和叶片含水率,同时将自身光合能力维持在较低水平;羧化效率与植物光合对CO2的利用率成正比[32],由于其气孔导度、胞间CO2浓度较低,CO2供给不足,土面植物通过自身较高的CO2利用率以维持光合作用。在石面小生境中,水热环境波动剧烈,水分和热量快速富集与散失,温度过高时植物为防止高温损伤便增加蒸腾速率降低温度,同时植物内部蒸腾耗水及碳同化导致出现干旱胁迫[35],因此植物在石面小生境中以较低的叶片含水率和叶面积增加自身水分剧烈变化的适应能力,并以较高的蒸腾速率避免自身光合结构不受损伤。石沟小生境相对较为封闭,水分和热量不易散失[24],水热条件较好[22],所以植物在相对适生的环境中,叶片呈现出较高的叶干物质含量、叶厚度、叶面积及较低的比叶面积、叶密度,并增加叶片气孔导度、胞间CO2浓度,提高净光合速率,以便能够快速生长和累积更多能量。研究还发现,在石面与石沟生境中,叶干物质含量与叶厚度极显著正相关(P<0.01),但与叶面积不显著相关,表明在石面与石沟小生境中,植物主要通过增加叶厚度的方式增加叶干物质含量,进而维持自身水分储备[36]。
叶经济谱能够从经济学角度反映植物在所处生境中自身对资源分配、利用、平衡等的适应策略[15]。主成分分析结果显示,在不同小生境中,各生活型植物分布基本一致,常绿乔木具有较高的叶干物质含量、叶密度以及较低的比叶面积和叶片含水率。常绿乔木在生长策略上更趋向于叶寿命长、叶干物质含量和叶密度高、生长缓慢,在叶经济谱中位于 “慢速投资收益性”一端,而常绿灌木与其相反位于“快速投资收益型”一端,落叶乔木则位于两者之间,属于中间型。
植物的生存策略是经过长期不断进化和适应而形成的。本研究从植物叶片功能性状的角度在一定程度上解释了典型喀斯特森林植物对不同小生境的适应性及其生存策略,但由于喀斯特区小生境分布的高度异质性、小生境内部环境的复杂性以及植物对养分吸收的选择性,还需要对其开展长期、连续的定位观测,进行更加全面和系统的研究与探索。
4 结论
在土面生境中,水分和养分资源供给稳定,植物对外界资源竞争的压力较小,同时叶干物质含量、叶厚度及光合水平相对较低,并且比叶面积与叶片含水率较高,因此其自身抗逆性、资源获取及保持能力相对较弱。植物为降低生态位重叠,趋向于生长缓慢、资源竞争力弱的生存策略。在石面生境中,植物生长所需的养分供给不足,水热条件严苛,资源竞争压力较大,叶干物质含量和光合水平相对高于土面植物,比叶面积、叶片含水率及叶面积则低于土面植物,因此其抗逆性以及对外界资源的获取及保持能力要强于土面植物,但因生境养分限制导致生长水平较低,植物趋向于生长缓慢但资源竞争力较强的生存策略。而在石沟生境中,水分和养分条件较好,资源竞争压力适中,植物叶干物质含量、叶厚度、叶面积及光合水平要高于其他生境植物,且比叶面积低,因此其抗逆性及对外界资源获取及保持能力较强,植物趋向于生长快速且资源竞争力较强的生存策略。
综合来看,石沟生境中,植物生长水平、抗逆性以及资源的获取与保持能力均要优于土面和石面。石面上植物生长水平虽受制约,但其具有较强的资源竞争能力,而土面生境中,植物由于资源供给稳定,资源竞争压力相对较小,资源获取与保持以及生长水平均低于石沟与石面生境。本研究中,叶经济谱存在于不同生活型植物间,而植物不同的生活型对叶片功能性状指标的影响要明显高于不同小生境对其的影响,小生境可能不是影响该地区植物叶片功能性状的主要因子。
