Tumor necrosis factor-α inhibition restores matrix formation by human adipose-derived stem cells in the late stage of chondrogenic differentiation
2022-12-02JiangTaoWanXianShuaiQiuZhuoHangFuYongCanHuangShaoXiongMin
Jiang-Tao Wan, Xian-Shuai Qiu, Zhuo-Hang Fu, Yong-Can Huang, Shao-Xiong Min
Abstract
BACKGROUND
Cartilage tissue engineering is a promising strategy for treating cartilage damage.Matrix formation by adipose-derived stem cells (ADSCs), which are one type of seed cell used for cartilage tissue engineering, decreases in the late stage of induced chondrogenic differentiation in vitro, which seriously limits research on ADSCs and their application.
AIM
To improve the chondrogenic differentiation efficiency of ADSCs in vitro, and optimize the existing chondrogenic induction protocol.
METHODS
Tumor necrosis factor-alpha (TNF-α) inhibitor was added to chondrogenic culture medium, and then Western blotting, enzyme linked immunosorbent assay,immunofluorescence and toluidine blue staining were used to detect the cartilage matrix secretion and the expression of key proteins of nuclear factor kappa-B (NF-κB) signaling pathway.
RESULTS
In this study, we found that the levels of TNF-α and matrix metalloproteinase 3 were increased during the chondrogenic differentiation of ADSCs. TNF-α then bound to its receptor and activated the NF-κB pathway, leading to a decrease in cartilage matrix synthesis and secretion. Blocking TNF-α with its inhibitors etanercept (1 μg/mL) or infliximab (10 μg/mL) significantly restored matrix formation.
CONCLUSION
Therefore, this study developed a combination of ADSC therapy and targeted anti-inflammatory drugs to optimize the chondrogenesis of ADSCs, and this approach could be very beneficial for translating ADSC-based approaches to treat cartilage damage.
Key Words: Adipose-derived mesenchymal stem cells; Human adipose-derived mesenchymal stem cells;Chondrogenic differentiation; Tumor necrosis factor-alpha; Etanercept; Infliximab; Nuclear factor kappa-B
INTRODUCTION
Adipose-derived stem cells (ADSCs) are mesenchymal stem cells with multidirectional differentiation potential that are isolated from autologous adipose tissue[1]. After years of research and application since the first successful isolation of ADSCs in 2001, they have become one of the most widely used types of adult stem cells in the field of tissue regeneration. In terms of their multidifferentiation ability, a large number of reported studies have shown that these cells can differentiate into a variety of cell types, such as bone cells, cartilage cells, and muscle cells of the motor system; myocardium cells, and vascular endothelial cells of the circulatory system; and nerve cells of the nervous system[2,3].Compared with bone marrow mesenchymal stem cells, adipose stem cells are abundant in tissue sources, are easy to extract, and exhibit higher proliferation[4]. However, similar to other tissue-derived adult stem cells, the capacity of ADSCs to synthesize and secrete cartilage matrix decreases during chondrogenic differentiation in vitro. There are several possible explanations for this phenomenon,including cell senescence, apoptosis[5], autophagy[6], oxidative stress[7], epigenetic inheritance[8], and abnormal cell adhesion[9]. Researchers have also proposed some signaling pathways that may participate in the process mentioned above, such as PI3K/Akt, and TGFβ/Smad3 signaling pathways.However, the specific mechanism remains to be further studied.
Tumor necrosis factor (TNF) is a serum glycoprotein that is produced by activated macrophages and other monocytes in mammals. Its functional unit is a homologous trimer that is composed of three subunits and 157 amino acids[10]. TNF, which is also known as TNF-alpha (TNF-α), exerts necrotizing effects on tumor cell lines and increases tumor transplant rejection. TNF-α is only 30% homologous to TNF-beta (lymphotoxin). Nevertheless, they share the same TNF receptors, namely, TNF-R1 and TNFR2.11 Many studies have shown that TNF-R1 mediates most of the biological activity of TNF. The combination of these receptors triggers a series of intracellular events that ultimately lead to the activation of two major transcription factors, namely, nuclear factor kappa-B (NF-κB) and C-Jun[11].Through these transcription factors, TNF-α induces the expression of genes that are essential for various biological processes, including cell growth and death, development, carcinogenesis, immunity, inflammation, and stress responses.
TNF-α inhibitors that are commonly used in clinical practice include etanercept, infliximab, and adalimumab. Etanercept is increasingly used because it is effective and affordable. Etanercept is a dimeric fusion protein that binds to TNF. Etanercept has been widely used in treating cartilage-related diseases, such as osteoarthritis, ankylosing spondylitis, and rheumatoid arthritis. Infliximab is a chimeric monoclonal IgG1 antibody that specifically binds to TNF-α, and it is mainly used in the study of autoimmune diseases, such as Crohn's disease, chronic inflammation and diabetic neuropathy. Both of these drugs inhibit the binding of TNF-α to its receptors on the cell surface, resulting in the biological inactivation of TNF-α, and their application is associated with few adverse reactions. In recent years,with the development of molecular biology, drugs with structures that are similar to etanercept, such as GP2015, LBEC0101, and Chs-0214, have been developed and have passed the assessments of a series of noninferiority studies[12]. In addition, some plant extracts, including saffron extract[13], resveratrol[14], nobiletin[15], etc., have also been reported to exert broad anti-inflammatory effects, and their effects are partly dependent on their ability to inhibit TNF-α.
In this study, we found that the expression of TNF-α increased during the early stage of the induced chondrogenic differentiation of hADSCs and resulted in a decrease in cartilage matrix secretion by hADSCs during late stages of chondrogenic differentiation; these effects could be delayed or eliminated by treatment with etanercept and Infliximab, which are TNF-α inhibitors (Figure 1A).


Figure 1 Human adipose-derived stem cells flow cytometry analysis results and differentiation ability identification. A: Schematic diagram of TNF-α interferes with Human adipose-derived stem cells (hADSCs) chondrogenic differentiation; B: Flow cytometer identification of surface molecules of hADSCs:CD44, CD29, CD45, CD34, and CD105; C: Oil red O staining and Western blot detection of marker proteins performed after lipogenic differentiation induction of hADSCs; D: Alizarin red, alkaline phosphatase staining, and Western blot detection of osteogenic differentiation marker proteins performed after osteogenic differentiation of hADSCs. MMP-3: Matrix metalloproteinase 3; NF-κB: Nuclear factor kappa-B; ECM: Extracellular matrix; GM: Human adipose-derived stem cells grown in growth medium alone.
MATERIALS AND METHODS
Cell culture
Human ADSCs (h ADSCs) were isolated from human fat tissue using the type I collagenase digestion method. The cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco; CA, United States) supplemented with 10% fetal bovine serum (FBS; Gibco) and 1% penicillin/streptomycin (Gibco)at 37 °C and a humidified 5% CO2atmosphere. The growth medium was changed every two days; thirdgeneration cells were used for the experiments. The cells were passaged when they reached 90% confluence. Third-generation cells were used for the experiments below.
Chondrogenic differentiation and treatment
Four groups were established as follows: (1) hADSCs grown in growth medium alone (GM group); (2)hADSCs treated with chondrogenic differentiation medium (CH group); (3) hADSCs treated with chondrogenic differentiation medium and 1 μg/mL etanercept (CHE group); and (4) hADSCs treated with chondrogenic differentiation medium and 10 μg/mL infliximab (CH+Inf group).
The chondrogenic differentiation medium was composed of basic high-glucose DMEM (1X), 5% FBS,1% penicillin/streptomycin, 1% insulin-transferrin-selenium-sodium-pyruvate solution (ITS-A; Gibco),100 nmol/L dexamethasone (Theremofisher; Massachusetts, USA) 50 μg/mL, 40 mg/mL L-proline(Macklin; Shanghai, China), and 10 ng/mL TGF-β3 (Peprotech; NJ, United States). For the CHE and CH+Inf groups, 1 μg/mL etanercept (MCE, NJ, United States) and 10 μg/mL infliximab (MCE) were added to the chondrogenic differentiation medium to inhibit the bioactivity of TNF-α. For every group,the GM was changed every other day, and all the treatments were applied when the cells reached confluence in the culture dishes.
The cells were harvested and assessed on days 7, 14, 21, and 28. The results presented are mainly those of Western blotting analysis, enzyme linked immunosorbent assay (ELISA), fluorescence imaging,and toluidine blue staining.
Flow cytometry
To identify the types of cells used in the experiment, passage 3 hADSCs were removed from the culture dish by digestion with 0.1% trypsin and centrifuged. The cells were suspended (106/mL), and the solution was aliquoted into EP tubes after washing twice with phosphate-buffered saline (PBS). Diluted antibodies, namely, FITC-labeled CD29, CD34, CD44, CD45, and CD105 antibodies, were added according to the antibody instructions. The mixtures were incubated at 4 °C for 30 min, and then, the supernatants were discarded after centrifugation. The remaining antibodies that had bound to cell surface molecules were removed by washing with PBS. The cells in each tube were suspended in 400 μL of PBS and placed into the flow tube. The processed cells were stored at 4°C in the dark or analyzed by flow cytometry.
Cell proliferation measurement
Passage 3 hADSCs were harvested and seeded in a 96-well culture plate. For the experimental group, on the second day, the GM was removed and replaced with chondrogenic differentiation medium containing etanercept. For the control group, the hADSCs were cultured in GM. Every 12 h, the CCK8 kit was used to measure the proliferation of the cells in these groups.
A premixed CCK-8 detection solution was added to the samples and incubated for 2 h. Finally, the absorbance of the samples was measured at 450 nm by a spectrophotometer.
Scratch test
Cell migration ability was test through scratch test. The tip of a 200 μL pipette was used to create several scratches of the same width in the Petri dishes. The cells in the dishes were cultured with GM and chondrogenic differentiation medium without FBS. The width of the scratches was observed and recorded under a microscope every 12 h.
Western blotting
Collagen type II (Col-2) and Aggrecan (ACAN) expression was measured with the Col-2 antibody(NB600-844, Novus, US.) and ACAN antibody (DF7561, Affinity, China) to prove chondrogenic differentiation. An anti-TNF-α antibody (AF7014, Affinity, China) was used to measure TNF-α expression.TNF-α functions mainly by activating the NF-κB pathway and upregulating the expression level of matrix metalloproteinase 3 (MMP-3). Therefore, we measured the expression of MMP-3, NF-κB p65, and pNF-κB p65 (phosphorylated NF-κB p65) with the corresponding antibodies (AF0217, AF5006, AF2006,respectively, Affinity, China).
The cells were lysed and the proteins were extracted with a protein extraction kit (KeyGEN BioTECH;Nanjing, China), and the lysis buffer was prepared according to the instructions. A spatula was used to scrape the cells off the bottom of the dish, and the cell fragments were mixed with lysis buffer. The mixtures were placed on ice, and the lysates were incubated for 30 min. The samples were centrifuged at 14,000 rpm at 4 °C for 15 min and boiled at 99.8 °C for 5 min.
The protein samples were loaded on sodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE) gels, and the separated proteins were transferred to polyvinylidene fluoride (PVDF)(Millipore, United States) membranes. Nonspecific binding sites were blocked with 5% bovine serum albumin (BSA) dissolved in Tris-buffered saline with Tween 20 (TBST). The membranes were then incubated with the antibodies mentioned above overnight at 4°C. The antibodies were then diluted strictly according to the instructions. The secondary antibody [Goat Anti-rabbit IgG (H+L) HRP, S0001,Affinity, China; Goat Anti-Mouse IgG (H+L) HRP, S0002, Affinity, China] was selected according to the species and origin of the primary antibody; the secondary antibody was added and incubated with the membranes at room temperature for 1 h. Signal detection was performed using the FDbio-Dura ECL kit(fdbio science, Hangzhou, China). ImageJ (software) was used to perform a semiquantitative analysis of the Western blotting bands.
Immunofluorescence microscopy analysis
The cells were seeded on cell slides and fixed with 4% paraformaldehyde for 10 min. The membranes of the cells were disrupted by incubation with 0.1% Triton-X 100 for 5 min. The samples were then blocked with 2% BSA dissolved in PBS for 1 h at room temperature. The samples were stained with the following primary antibodies: anti-MMP-3 antibody (1:200, AF0217) and anti-TNFα antibody (1:200,AF7014). Actin was stained with Phalloidin-iFluor 594 Reagent (ab176757). A goat anti-rabbit IgG/Alexa Fluor 555 antibody (Bioss, Beijing, China) was used as the secondary antibody. After these steps were complete, DAPI (4',6-diamidino-2-phenylindole) was used to label the nuclear DNA.
ELISA
The cell supernatants were collected when the culture medium was changed on days 7, 14, 21, and 28. A proteinase inhibitor was added (1 μg/mL) to the samples, and the samples were stored at -80 °C. The cells were starved for 24 h in medium without FBS before the cell supernatants were collected. The concentrations of TNF-α and MMP-3 in the supernatants were measured using specific ELISA Kits(SEA133 Hu, SEA101Hu, Clone Cloud, Wuhan, China) according to the instructions.
Toluidine blue staining
Toluidine blue (0.1 g) was dissolved in 100 mL of 0.2 mol/L acetate buffer and stirred with magnetic stirrers for 1 h. The solution was then filtered; the pondus hydrogenii (pH) of the final solution was approximately 3.72 to 4.25. The hADSCs were fixed with 4% paraformaldehyde for 15 min. Then,toluidine blue and stain were added to the shaker for 30 min. Finally, cytoplasmic staining was observed under a microscope.
Statistical analysis
All the experiments were repeated more than three times. A one-way analysis of variance (ANOVA) or t test (GraphPad Prism 9.0 software, La Jolla, CA, United States) was used to identify significant differences. P < 0.05 was considered statistically significant.
RESULTS
The cells that were isolated from adipose tissue exhibited properties of mesenchymal stem cells
Flow cytometry showed that CD44, CD29, and CD105 were highly expressed, while CD34 and CD45 were hardly expressed, on the cell surfaces (Figure 1B), which was consistent with the reported molecular expression pattern of mesenchymal stem cells. The cells were treated with adipogenic induction medium and osteogenic induction medium. The expression of marker proteins of differentiation was measured, and characteristic staining was performed. The results showed that the expression of CCAAT-enhancer-binding protein-beta (CEBPB), which is the transcription factor that is characteristic of adipogenic differentiation, increased after 7 d of treatment with adipogenic induction medium. The cells were also positive for Oil red O staining (Figure 1C). In addition, the expression of Runt-related transcription factor 2 (RUNX2) and osteopontin (OPN), which are markers of osteogenic differentiation, increased and peaked on day 14 after treatment with osteogenic induction medium;alkaline phosphatase staining and alizarin red staining were also positive (Figure 1D).
A low concentration of etanercept did not reduce the proliferation of hADSCs
In the CCK-8 experiments, hADSCs that were treated with chondrogenic differentiation medium containing 1 μg/mL etanercept showed similar growth curves to those that were treated with GM(Figure 2A). In addition, the migration ability of the hADSCs that were grown in these two culture media was assessed with a scratch experiment. The curve of scratch width with time showed that there was no significant difference in the migration ability of hADSCs grown in GM and CHE medium(Figure 2B). After being cultured for 48 h, the scratches disappeared in both groups.
The expression of cartilage matrix decreased during the late stage of hADSC chondrogenic differentiation
One week after the induction of chondrogenic differentiation, toluidine blue staining showed darker blue staining in the cytoplasm and the surrounding region of the cells (Figure 2D), suggesting the accumulation of cartilage matrix and the differentiation of hADSCs into chondrocytes. Western blotting analysis showed that compared with GM, chondrogenic differentiation medium increased the expression of Col-2 and Aggrecan by hADSCs from weeks 1 to 4, and the expression peaked from days 14 to 21 (Figure 2D). The expression of the chondrogenic marker proteins showed a declining trend on day 28 (Figure 2D).
The expression of TNF-α was increased in the early chondrogenic differentiation of hADSCs, and MMP-3 expression was upregulated in late differentiation
The WB results showed that TNF-α expression was increased in the early stage of chondrogenic differentiation (specifically, it increased by 3.7 times on day 7, P < 0.05), and its expression level was not significantly affected by the presence of interleukin (IL)-1β. Compared with the control group, the expression level of MMP-3 was not significantly increased at week 1 of hADSC chondrogenic differentiation, but it was significantly increased at week 4 (it increased by nearly 15 times on day 28, P < 0.1)(Figure 3A). The ELISA results were consistent with the Western blotting results. The TNF-α concentration in the cell supernatants increased by 2.4 times on day 7 (P < 0.1), and the concentration of MMP-3 in the cell supernatants increased by 10.8 times on day 28 (P < 0.1) (Figure 3B).
At 1 and 4 wk of chondrogenic differentiation, the cellular localization of TNF-α and MMP-3 in hADSCs was analyzed, and their concentrations were semiquantitatively measured, using immunofluorescence staining. The results were consistent with the Western blotting results. TNF-α expression was increased in the early stage of chondrogenic induction, while MMP-3 expression was significantly increased in the late stage. The fluorescence of these two proteins was primarily concentrated in the cytoplasmic region, which was identified by the red fluorescence of the microfilaments (Figure 3C).
IL-1β (10 ng/mL) was added to the culture medium, and chondrogenic differentiation of hADSCs was observed. Western blotting analysis showed that treatment with IL-1β alone did not affect the expression of TNF-α or MMP-3 in hADSCs. However, it upregulated the expression of MMP-3 in the presence of chondrogenic differentiation medium. Surprisingly, the expression level of TNF-α was slightly downregulated by chondrogenic differentiation medium containing IL-1β. Toluidine blue staining showed that the staining was lighter in the group treated with IL-1β and chondrogenic differentiation medium than in the group treated with chondrogenic differentiation medium alone on day 7.The staining of hADSCs with IL-1β alone without chondrogenic differentiation medium showed no significant difference from the control group (Figure 3D). As the viability of ADSCs was affected by IL-1 β, a large number of ADSCs died after 7 d of treatment, so the long-term effects of IL-1β could not be observed. These research results need to be further explored.
Etanercept and infliximab alleviated the reduced secretion of cartilage matrix by hADSCs during chondrogenic induction
hADSCs treated with chondrogenic differentiation medium containing 1 μg/mL etanercept maintained their ability to secrete cartilage matrix for 28 d (Figure 4A, Supplementary Figure 1A). Toluidine blue staining showed that after 28 d of treatment with etanercept combined with chondrogenic induction medium, the staining intensity of the group treated with etanercept combined with chondrogenic induction medium was higher than that of the group treated with chondrogenic differentiation medium alone, and the intensities of both groups were higher than that of the control group (Figure 4B). ELISA showed that the concentrations of TNF-α and MMP-3 in the cell supernatants decreased after etanercept treatment (Figure 4C).
Toluidine blue staining showed that the cartilage matrix accumulation of CH+Inf group was more than that of CH group and GM group but not as much as CHE group (Figure 5A). hADSCs treated with chondrogenic differentiation medium containing 10 μg/mL infliximab maintained their ability to secret cartilage matrix capacity 28 d (Figure 5B and Supplementary Figure 1B). Western blot showed that the expression of TNF-α and MMP-3 in CH+Inf group was lower than CH group on both Day 14 and Day 28 (Figure 5C).
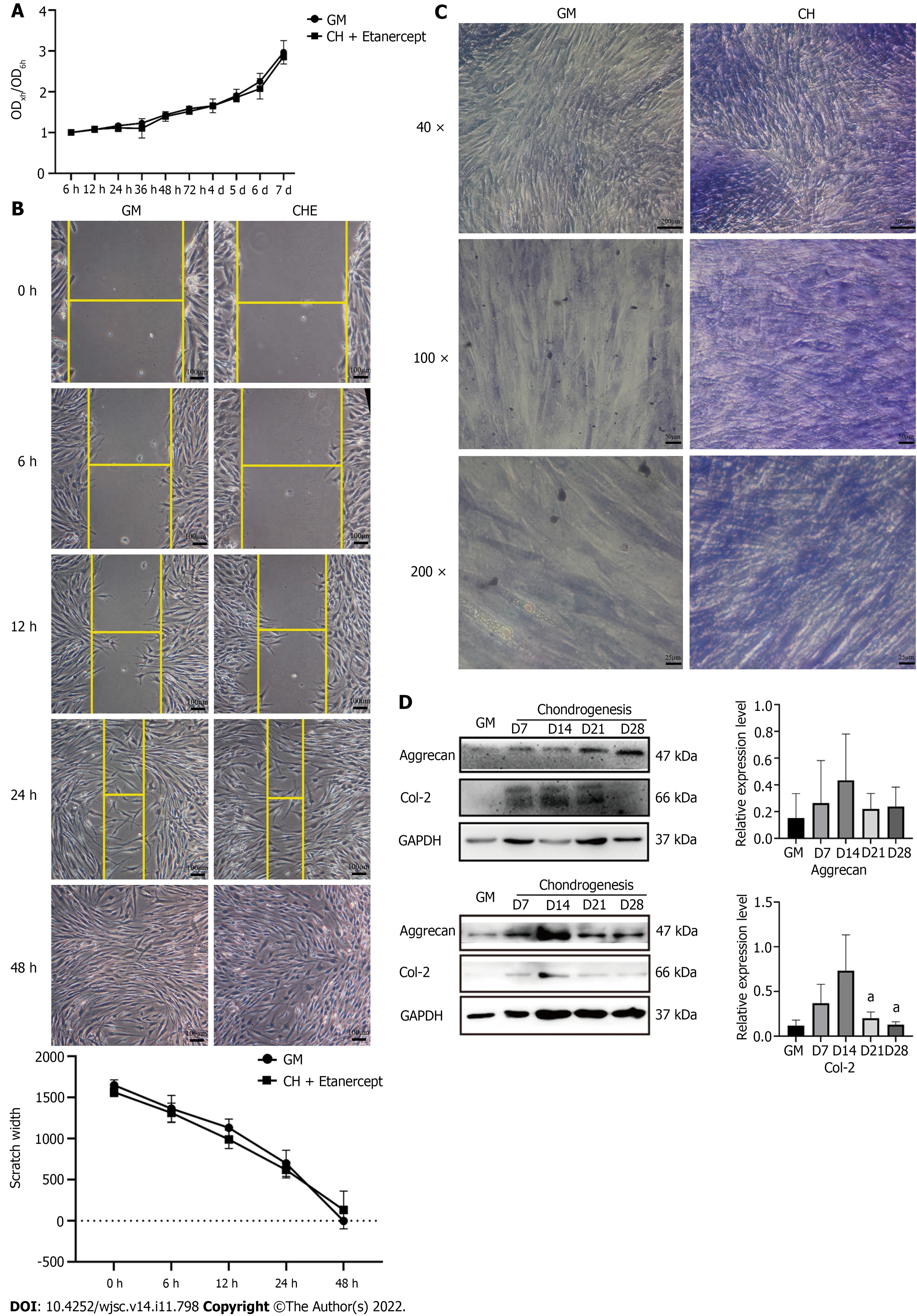
Figure 2 Etanercept cytotoxicity analysis and human adipose-derived stem cells chondrogenic differentiation ability identification. A: Thegrowth curves of human adipose-derived stem cells (hADSCs) treated with chondrogenic medium containing etanercept were measured using the CCK-8 method; B:Scratch test performed to determine the migration of hADSCs treated with chondrogenic medium containing etanercept; C and D: Toluidine blue staining and Western blot detection of chondrogenic differentiation marker proteins. aP < 0.05. GM: Human adipose-derived stem cells (hADSCs) grown in growth medium alone; CHE:hADSCs treated with chondrogenic differentiation medium and 1 μg/mL etanercept.
Etanercept and infliximab inhibited the binding of TNF-α to its receptor and delayed the decrease in the secretion of cartilage matrix by hADSCs via the NF-κB pathway
The levels of total and phosphorylated NF-κB p65 in hADSCs were increased during the induction of chondrogenic differentiation. After treatment with 1 μg/mL etanercept and 10 μg/mL infliximab, the expression level of NF-κB p65 did not significantly change, but the level of phosphorylated NF-κB p65 decreased on day 28 (Figures 4D, 5D and Supplementary Figure 1C). In addition, the expression level of MMP-3 decreased on day 28 in the CHE group.
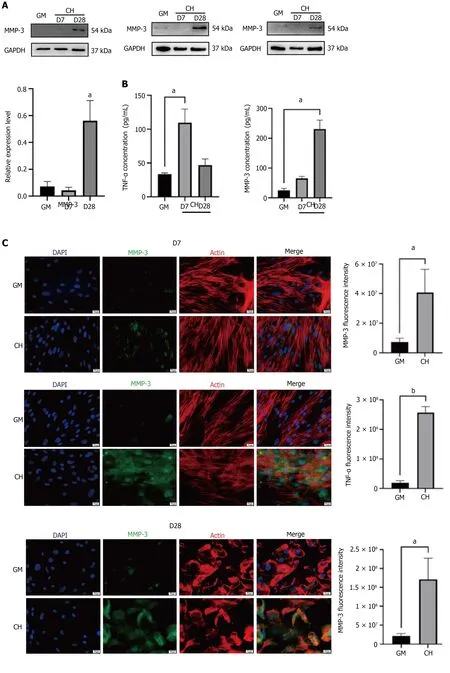
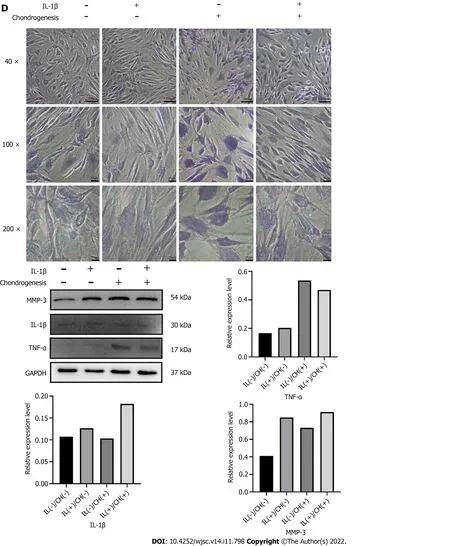
Figure 3 Expression levels of tumor necrosis factor-alpha and matrix metalloproteinase 3 were detected during chondrogenic differentiation of human adipose-derived stem cells. A: Western blot detection of MMP-3; B: Elisa detection of the concentrations of tumor necrosis factoralpha (TNF-α) and matrix metalloproteinase 3 (MMP-3) in the cell supernatant; C: Immunofluorescent staining of TNF-α and MMP-3; D: Toluidine blue staining and the Western blot detection of TNF-α and MMP-3 of human adipose-derived stem cells treated with chondrogenic medium containing interleukin-1β. aP < 0.05; bP <0.01. TNF-α: Tumor necrosis factor-alpha; MMP-3: Matrix metalloproteinase 3; IL-1β: Interleukin-1β; GM: Human adipose-derived stem cells (hADSCs) grown in growth medium alone; CH: hADSCs treated with chondrogenic differentiation medium.
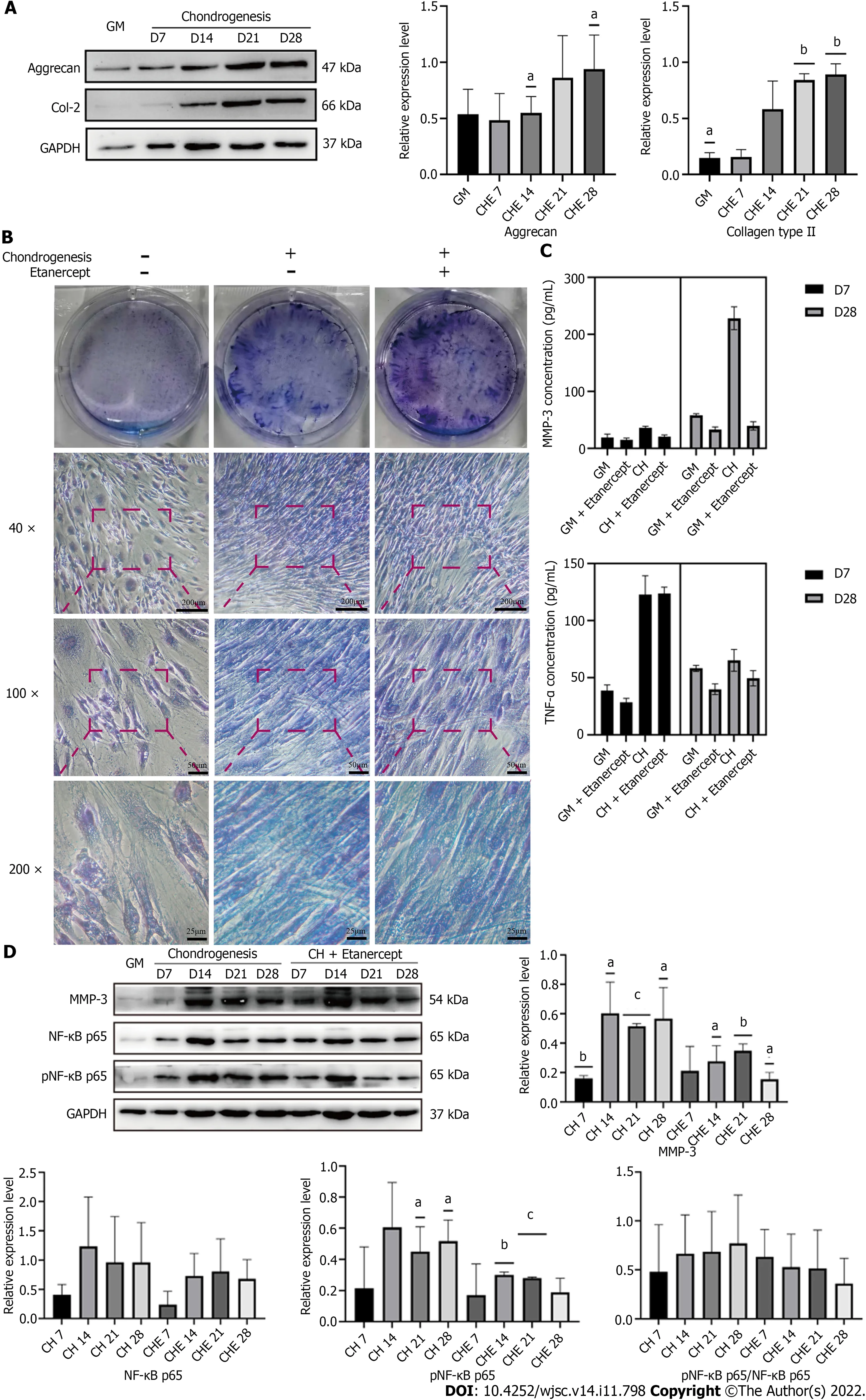
Figure 4 Detection of human adipose-derived stem cells chondrogenic differentiation marker proteins and tumor necrosis factor-alpha and matrix metalloproteinase 3 performed after treatment with chondrogenic differentiation medium containing etanercept. A: Western blot detection of Col-2 and Aggrecan of adipose-derived stem cells treated with chondrogenic differentiation medium containing etanercept; B: Toluidine blue staining; C:Elisa detection of the tumor necrosis factor-alpha and matrix metalloproteinase 3 (MMP-3) concentration in cell supernatant; D: Western blot detection of MMP-3 and NF-κB pathway. aP < 0.05; bP < 0.01; cP < 0.001. MMP-3: Matrix metalloproteinase 3; NF-κB: Nuclear factor kappa-B; GM: Human adipose-derived stem cells(hADSCs) grown in growth medium alone; CH: hADSCs treated with chondrogenic differentiation medium; CHE: hADSCs treated with chondrogenic differentiation medium and 1 μg/mL etanercept.

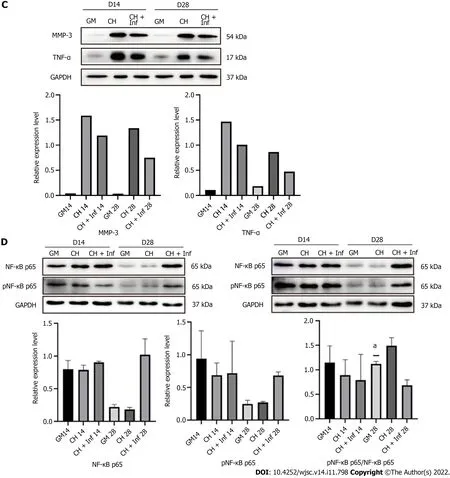
Figure 5 Detection of human adipose-derived stem cells chondrogenic differentiation marker proteins and tumor necrosis factor-alpha and matrix metalloproteinase 3 performed after treatment with chondrogenic differentiation medium containing Infliximab. A: Toluidine blue staining; B: Western blot detection of Col-2, Aggrecan and Sox-9 of adipose-derived stem cells treated with chondrogenic differentiation medium containing Infliximab;C: Western blot detection of matrix metalloproteinase 3 and tumor necrosis factor-alpha pathway; D: Western blot detection of nuclear factor kappa-B pathway. aP <0.05. TNF-α: Tumor necrosis factor-alpha; MMP-3: Matrix metalloproteinase 3; IL-1β: Interleukin-1β; GM: Human adipose-derived stem cells (hADSCs) grown in growth medium alone; CH: hADSCs treated with chondrogenic differentiation medium; CHE: hADSCs treated with chondrogenic differentiation medium and 1 μg/mL etanercept; CH+Inf: hADSCs treated with chondrogenic differentiation medium and 10 μg/mL infliximab.
DISCUSSION
In this study, we explored the possibility that the inflammatory environment caused by the elevated levels of TNF-α in the ADSC culture microenvironment at the early stage of chondrogenic differentiation induction affected the synthesis and secretion of cartilage matrix at the late stage of chondrogenic differentiation. Therefore, we used etanercept to inhibit TNF-α throughout the entire process of chondrogenic differentiation in vitro and observed changes in the cartilage matrix synthesis and secretion of ADSCs.
The induction of hADSCs to form cell lines that are capable of continuously secreting cartilage matrix in vitro is one approach for treating diseases that involve damaged cartilage. However, the decrease in cartilage matrix secretion in the late stage of chondrogenic differentiation is a major limitation to this research. Researchers have tried to solve this problem in several ways. Examples include: (1) 5-Aza cytidine (5-AZAC) was used to reduce the DNA methylation level[8]; (2) The expression level of cartilage matrix proteins was upregulated using noncoding RNA[16]; (3) A variety of cytokines, such as BMP and TGF-β, were added to culture[17]; and (4) A three-dimensional culture system was generated based on biomaterial scaffolds[18,19].
However, none of these methods could completely solve the problem that the production of cartilage matrix decreases during the late stage of the induced chondrogenic differentiation of stem cells in vitro.In this study, during the induced chondrogenic differentiation of hADSCs in vitro, the inflammatory factors TNF-α and MMP-3 accumulated in the culture system. Moreover, the ability of the cells to secrete cartilage matrix decreased. Therefore, we conducted a series of studies to determine whether there was a link between these two phenomena.
Long-term cartilage damage, which is characterized by local damage that is caused by the implant surgery, the immune reaction to implants, and the inflammatory factors that are inherent to the implants, can create an inflammatory microenvironment that is not conducive to stem cell therapy[20].Improving the inflammatory state of the microenvironment can enhance many therapeutic responses, as has been reported in stem cell transplantation therapy, rheumatoid arthritis and psoriasis treatment,and even depression treatment[21]. In a study on hematopoietic stem cell transplantation, the direct binding of donor-derived TNF-α to TNF-R1 impaired the survival and division of transplanted hematopoietic stem cells and progenitor cells[22]. Anti-TNF therapy has also been shown to increase the success of hematopoietic stem cell therapy in treating human adenosine deaminase deficiency. The response to anti-TNF therapy can be considered one of the indications of whether the patient is a candidate HSCT[23]. Under physiological conditions, nearby stem cells are the first to differentiate and repair the damage when the body is injured[24,25]. Therefore, a microenvironment that is similar to that of the target tissue can greatly facilitate stem cell-induced differentiation in vitro. In this study, we found that TNF-α in the culture system upregulated the expression of MMP-3, which degraded the extracellular cartilage matrix, resulting in a lack of external support for the induced differentiation of hADSCs.
The NF-κB pathway is the primary signaling pathway that is activated when TNF-α binds to TNFR[11,26]. This pathway is often associated with inflammation by researchers[27] and is widely involved in the homeostatic regulation of the musculoskeletal system[28]. This pathway has been described in many studies to block chondrogenic differentiation[29,30]. In this study, etanercept and infliximab successfully reduced the level of phosphorylated NF-κB p65 and the expression level of MMP-3. This study confirmed that the NF-κB pathway is one of the signaling pathways by which TNF-α blocks the chondrogenic differentiation of hADSCs. It is worth considering whether drugs that inhibit TNF-α can be used to improve the protocol of ADSC chondrogenic induction in vitro. It has been reported that some substances, such as hawthorn[31], saffron extract[13] and resveratrol[14], can not only inhibit the biological effects of TNF-α but also have antioxidant and antifibrosis abilities. Could these substances be more effective components for improving the chondrogenic induction medium? In addition, our study found that the continuous accumulation of inflammation level may be one of the reasons for the decreased cartilage matrix secretion. Are there other substances besides TNF-α involved in the inflam-matory response of the culture system? Can therapies targeting other inflammatory targets, such as TIMPs and NSAIDs, also be used to assist the chondrogenic differentiation of ADSCs in vitro? Further research is needed to find out the answers.
CONCLUSION
Through this study, we confirmed that TNF-α increased and inhibits the secretion of cartilage matrix by activating the NF-κB pathway. The use of TNF-A inhibitors, such as etanercept and infliximab, can maintain the cartilage matrix secretion of ADSCs at the late stage of chondrogenic differentiation. We hope to highlight an approach that combines stem cell therapy and targeted anti-inflammatory drugs to treat diseases that involve cartilage damage. The adverse effects associated with in vitro-induced chondrogenic differentiation can be eliminated and the therapeutic effect of hADSCs can be maximized with specific targeted drugs. Animal and in vivo studies remain to be conducted, which is the future direction of our team.
ARTICLE HIGHLIGHTS

Research motivation
In the previous study, we conducted single-cell sequencing of ADSCs before and after chondrogenic differentiation and found that the expression levels of Matrix metalloproteinase 3 (MMP-3) and Tumor necrosis factor receptor superfamily member 12A (TNFRSF12A) were significantly increased. Therefore,we hypothesized that the accumulation of inflammatory levels in THE culture system resulted in decreased cartilage matrix secretion at the late stage of differentiation and designed this study for this reason.
Research objectives
To investigate the changes of tumor necrosis factor-α (TNF-α) and MMP-3 Levels in the culture system of ADSCs before and after chondrogenic differentiation. To confirm that TNF-α increased and decreased cartilage matrix secretion of ADSCs by activating the NF-KB pathway. To confirm that adding TNF-α inhibitor to chondrogenic medium could improve the chondrogenic differentiation efficiency of ADSCs.
Research methods
Treat ADSCs with chondrogenic medium containing TNF- α inhibitors, such as etanercept and infliximab. Then observe the changes of cartilage matrix secretion and the level of inflammation in the culture system through western blot, Elisa, immunofluorescence and toluidine blue staining.
Research results
During the differentiation of ADSCs, the expression levels of TNF-α and MMP-3 increased gradually,and the activation of NF-κB signaling pathway increased. Adding TNF-α inhibitors, etanercept (1 μg/mL) or inflixib (10 μg/mL), to the chondrogenic medium can reduce the activation of NF-κB pathway, alleviate the inflammation and preserve the secretion of cartilage matrix of ADSCs.
Research conclusions
When TNF-α increases and binds to its receptor, activates NF-κB pathway and reduces cartilage matrix secretion of ADSCs. TNF-α inhibitors can block the above process and improve the chondrogenic differentiation efficiency of ADSCs in vitro.
Research perspectives
In future studies, the TNF-α inhibitor etanercept or infliximab used in this study could be combined with the scaffold material to optimize the growth environment of ADSCs and make the drug release more durable and gentle, so as to achieve higher chondrogenic differentiation efficiency of ADSCs. And obtain a transplantable cartilage engineering material with similar properties to the natural cartilage tissue for the repair and treatment of cartilage defects, eventually.
ACKNOWLEDGEMENTS
Thanks to Peking University Hong Kong University of Science and Technology Medical Center for providing the research instruments and workplace.
FOOTNOTES
Author contributions: Min SX and Wan JT conceived and designed the experiments; Wan JT, Qiu XS, and Fu ZH performed the experiments; Wan JT and Qiu XS analyzed the data; Wan JT and Huang YC composed the manuscript.
Supported by the Natural Science Foundation of Guangdong Province, China, No. 2017A030313564; and Shenzhen High-level Hospital Construction Fund to Peking University Shenzhen Hospital.
Institutional review board statement: This study was approved by the Ethics Committee of Peking University Shenzhen Hospital (No. 2022-076).
Conflict-of-interest statement: The author has no conflict of interest to declare.
Data sharing statement: No additional data are available.
Open-Access: This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non commercial. See: https://creativecommons.org/Licenses/by-nc/4.0/
Country/Territory of origin: China
ORCID number: Yong-Can Huang 0000-0001-8548-8233; Shao-Xiong Min 0000-0003-3334-6254.
S-Editor: Gong ZM
L-Editor: A
P-Editor: Cai YX
