外源Ca2+对模拟酸雨胁迫下不同抗性水稻根系氮吸收的影响
2022-12-02张元棋梁婵娟
张元棋,梁婵娟,2*
(1.江苏省厌氧生物技术重点实验室,江南大学环境与土木工程学院,江苏 无锡 214122;2.江苏水处理技术与材料协同创新中心,苏州科技大学,江苏 苏州 215009)
尽管近年来全球酸雨态势有所减缓,但我国仍有4.66×107hm2的国土面积为酸雨区[1]。酸雨会抑制植物根系生长[2],破坏其吸收养分的能力[3];引起活性氧过量积累,造成氧化损伤[4-5];破坏叶绿体微观结构,减少叶绿素含量,降低光合作用能力[6-7]。酸雨还会通过淋溶土壤中的,使土壤贫瘠[8]、农作物生长缓慢,引起农业减产,提高作物耐受性可能是缓解酸雨区农业减产的一种可行方法。
已有研究发现外源Ca2+能提高植物对盐、干旱及低温等非生物胁迫的耐受能力[9-12]。酸雨胁迫下,外源Ca2+能通过提高抗氧化酶活性、同工酶浓度及基因表达缓解酸雨造成的氧化胁迫[4-5],通过调节细胞质膜组分及胞内Ca2+形态促进质膜稳定性[13],提高植物对酸雨的耐受性。同时有研究发现外源Ca2+缓解酸雨胁迫下大豆品质与调控其营养吸收有关[14]。然而外源Ca2+和酸雨胁迫下植物营养吸收与植物耐酸性的关系仍不明确,明确这一点将有助于提高植物耐酸性,从而为缓解酸雨对作物产量及品质的影响提供良好的生长基础。氮是限制植物生长最重要的因素之一[15]。水稻主要以NO-3和的形式吸收无机氮,这一过程需要H+-ATPase 水解ATP提供能量,同时向胞外泵出H+维持胞内pH 稳定,保障氮吸收的稳定和平衡是维持水稻生长和发育的基础。明晰外源Ca2+如何通过H+-ATPase 调节酸雨胁迫下植物氮吸收,将为丰富植物耐酸性机制提供新的理论支撑,同时也为缓解酸雨造成的土壤肥力下降提供思考方向。
为了提高植物耐酸性,同时进一步探明外源Ca2+调控机理和调控效果的品种间差异,本研究选取了两个广泛种植的水稻品种——五优308(抗性种)和南粳9108(敏感种)。首先,探究外源Ca2+对酸雨胁迫与恢复后水稻根系生长与氮(NO-3和)含量的影响,明晰外源Ca2+调控酸雨胁迫下植物氮吸收与耐受性的关系;其次,测定根系 NO-3、吸收速率,ATP 含量和质膜H+-ATPase 活性,探究外源Ca2+调控酸雨胁迫下植物根系质膜H+-ATPase 活性对氮吸收的影响;最后,从质膜H+-ATPase 磷酸化水平角度探析外源Ca2+调控酸雨胁迫下植物根系质膜H+-ATPase 活性的内在机制。通过对比外源Ca2+与酸雨处理下两个水稻品种生长、氮吸收及质膜 H+-ATPase 活性与磷酸化的差异,客观评价外源Ca2+调控植物耐酸性的效果,为采取有效措施减轻酸雨对农业生产的影响提供理论依据,为缓解酸雨引起的粮食安全问题提供新的选择。
1 材料与方法
1.1 试验材料
试验试材为水稻(Oryza sativaL.),品种为五优308(抗性种)和南粳9108(敏感种)。品种筛选是根据郭莉莉等[16]的研究结果,考虑长江中下游地区种植的常见水稻品种,并在模拟酸雨(pH 4.5 或3.0)下通过相对生长速率、根系质膜透性以及叶片叶绿素含量指标筛选比较得出的抗性差异。将种子浸泡于0.1%的HgCl2中10 min后取出,使用去离子水冲洗干净后,置于去离子水中浸泡24 h。种子转移至蛭石中,于25 ℃恒温萌发,长至两叶时转移至24 孔泡沫板并置于6.88 L 周转箱培养,培养条件参考马永佳等[13]的方法。营养液配制按照国际水稻研究所常规水稻营养液配方,营养液每3 d更换一次。
1.2 试验方法
1.3 指标测定方法
试验材料使用去离子水冲洗,105 ℃烘30 min 后80 ℃烘至质量恒定,测定根系干质量。采用水杨酸-硫酸法和靛酚蓝比色法分别测定根(以鲜质量计)中NO-3和含量[18-19]。
采用GAO 等[20]的方法测定并计算NO-3和的吸收速率,计算公式分别为:
NO-3吸收速率=(初始营养液中NO-3浓度-处理5 d后营养液中NO-3浓度)×营养液体积(/根鲜质量×5)
质膜H+-ATPase 活性使用质膜蛋白提取试剂盒(贝博生物科技有限公司,上海)提取测定。参考LIANG 等[21]的方法使用定磷试剂测定反应体系中生成的无机磷含量,并根据以下公式进行计算:

式中:E为酶活,µmol·mg-1·min-1;C为标准曲线上查得的无机磷含量,µg·mL-1;t为反应时间,min;Wp为膜制剂中的蛋白含量,mg·mL-1;n为稀释倍数。
H+-ATPase 磷酸化水平使用Western-Blot 方法测定,使用8%浓缩胶和6%分离胶在120 V、30 min 和80 V、55 min 条件下电泳以分离目标蛋白。使用半干法转膜后用5%BSA溶液封闭PVDF膜2 h,以1∶2 000比例4 ℃静置过夜孵育一抗(14-3-3 蛋白),用PBST溶液清洗3 次,再以1∶1 000 比例加入二抗(HRP-驴抗兔 IgG 抗体)室温孵育 2 h,最后用 PBST 清洗 3 次,显影后进行拍摄。
根系ATP 含量(以鲜质量计)测定参考文献[22],使用高效液相色谱法测定。具体条件为C18柱(250 mm×4.6 mm,5 µm),柱温25 ℃,流速1.2 mL·min-1,UV 254 nm,进样量 20 µL,流动相 A 为 0.06 mol·L-1Na2HPO4、0.04 mol·L-1KH2PO4,pH 7.0,流动相B 为纯乙腈。洗脱程序:0 min 100% A;2 min 95% A 和5%B;4 min 80% A 和 20% B;5.3 min 75% A 和 25% B;6 min 100%A。
1.4 数据处理
采用SPSS 18.0 对数据进行显著性分析,不同字母表示处理间差异显著(P<0.05)。数据分析后使用Origin 2021 绘图,绘图数据均为3 次独立试验数据的平均值±标准误差(Mean±SD)。
2 结果与分析
2.1 外源Ca2+对模拟酸雨下不同抗性水稻幼苗根系生长和根中NO-3、含量的影响
如图 1 所示,胁迫期,与 CK 相比,SAR1 处理下,五优308 幼苗根系干质量以及NO-3和含量无显著变化(P>0.05);南粳9108 幼苗根系干质量显著降低(P<0.05),NO-3含量无显著变化(P>0.05),含量显著增加(P<0.05)。SAR2 处理下,两个品种水稻根系干质量、NO-3和含量均低于CK(P<0.05),五优308的降幅分别为15%、24%、19%,南粳9108的降幅分别为31%、42%、20%。单一外源Ca2+处理对两个品种水稻根系生长以及根中NO(-3除五优308)和含量均无显著影响(P>0.05)。Ca2++SAR1 处理下,两个品种水稻幼苗根系干质量、根中NO-3和含量与 CK 均无显著差异(P>0.05),但两个品种水稻的根干质量均显著高于 SAR1 处理(P<0.05)。Ca2++SAR2 处理下,两个品种水稻幼苗根干质量、根中NO-3和含量均低于CK(P<0.05),但显著高于SAR2 处理(P<0.05),五优308 上述3 个指标分别达到CK 的93%、88%、92%,南粳9108 分别达到CK 的77%、80%、88%。恢复期,五优308 各处理组中根系生长、NO-(3除Ca2++SAR2 处理)和含量均恢复至CK 水平(P>0.05)。在南粳 9108 中,SAR2 处理和 Ca2++SAR2 处理下根系生长和含量仍低于CK(P<0.05),但Ca2++SAR2 处理高于SAR2 处理(P<0.05)。SAR2 处理下南粳9108根中NO-3含量低于CK(P<0.05),但在Ca2++SAR2 处理下恢复至CK水平(P>0.05)。

图1 外源Ca2+对模拟酸雨下不同抗性水稻根干质量和根中NO-3、含量的影响Figure 1 Effects of exogenous Ca2+on root dry mass,NO-3 and contents of roots in different resistant rices to simulated acid rain stress
2.2 外源Ca2+对模拟酸雨下不同抗性水稻根系NO-3和吸收速率的影响
如图 2 所示,胁迫期,SAR1 处理下,五优 308 的NO-3和吸收速率高于 CK(P<0.05),南粳 9108 的NO-3吸收速率无显著变化(P>0.05),吸收速率高于CK(P<0.05)。SAR2 处理下,两个品种水稻根系NO-3和吸收速率均低于 CK(P<0.05),五优 308 降幅分别为42%、27%,南粳9108 降幅分别为45%、33%。单一外源Ca2+处理下五优308 的NO-3吸收速率高于 CK(P<0.05),吸收速率无显著变化(P>0.05);南粳9108的NO-3和吸收速率均无显著变化(P>0.05)。Ca2++SAR1处理下,两个品种水稻根中NO-3和吸收速率与CK 无显著差异(P>0.05),NO-3吸收速率与SAR1 处理无差异(P>0.05),吸收速率低于 SAR1 处理(P<0.05)。Ca2++SAR2 处理下,两个品种水稻的NO-3和吸收速率均低于CK 水平(P<0.05),但高于SAR2处理(P<0.05),五优308中分别为CK 的 83%、87%,南粳 9108 中分别为 CK 的 82%、80%。恢复期,五优308除在SAR2处理下NO-3吸收速率仍低于CK 外(P<0.05),其余处理均恢复至CK 水平(P>0.05),吸收速率在所有处理下均恢复至CK水平(P>0.05)。南粳9108在SAR2处理下NO-3和吸收速率仍低于CK 水平(P<0.05),Ca2++SAR2 处理下NO-3吸收速率低于CK但高于SAR2处理(P<0.05),其他处理下NO-3和吸收速率均恢复至CK水平(P>0.05)。
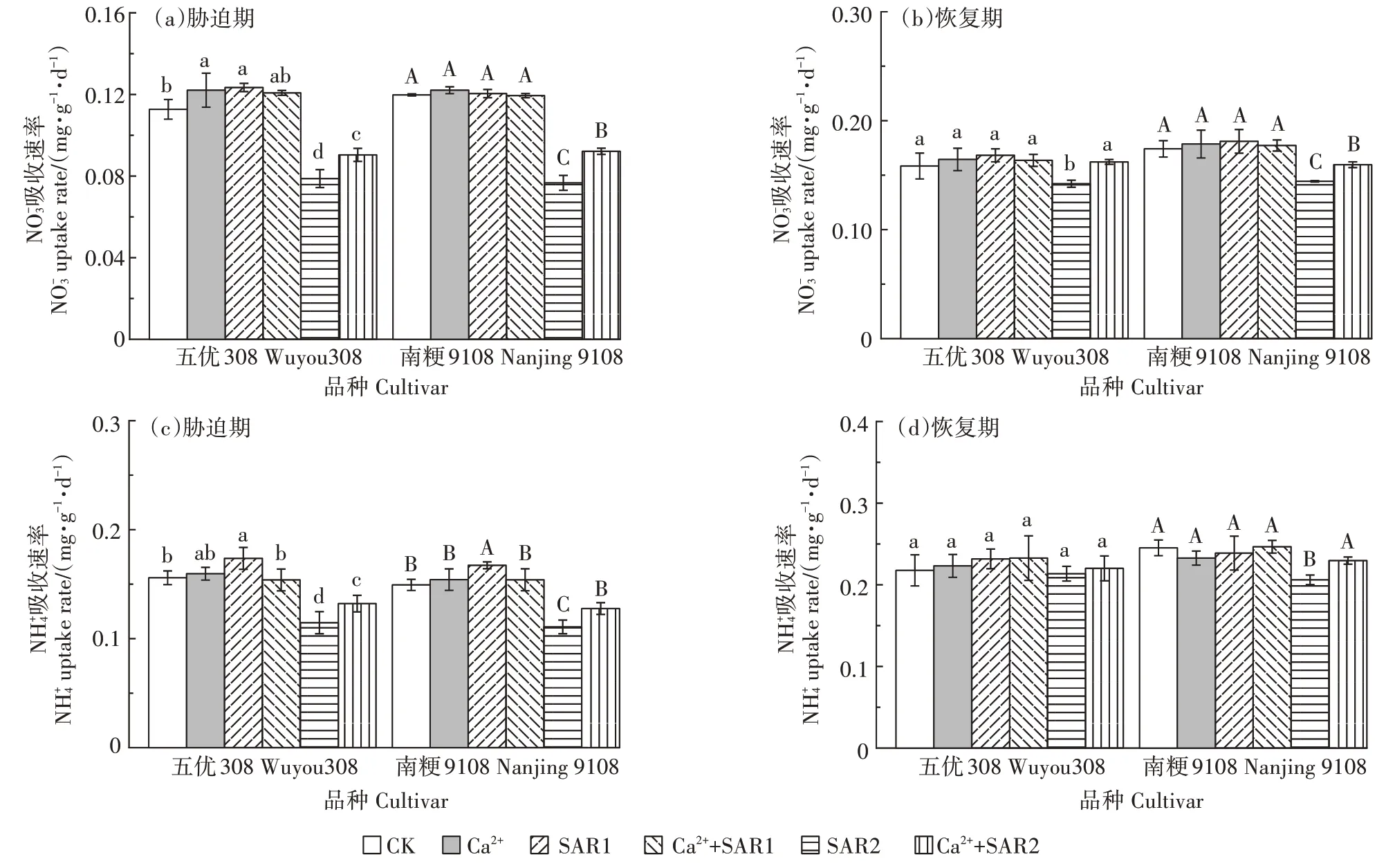
图2 外源Ca2+对模拟酸雨下不同抗性水稻根系NO-3和吸收速率的影响Figure 2 Effects of exogenous Ca2+on NO-3 and uptake rates of different resistant rices root under simulated acid rain stress
2.3 外源Ca2+对模拟酸雨下不同抗性水稻根系质膜H+-ATPase活性和根中ATP含量的影响
如图3 所示,胁迫期,SAR1 处理下两个品种水稻根系质膜 H+-ATPase 活性高于 CK(P<0.05),ATP 含量均降低(P<0.05),五优 308 和南粳 9108 中 H+-ATPase 活性增幅分别为16%、20%,ATP 含量降幅分别为25%、31%。SAR2 处理下,两个品种水稻H+-ATPase 活性以及 ATP 含量均低于 CK(P<0.05),五优308 降幅分别为32%、31%,南粳9108 降幅分别为43%、50%。单一外源Ca2+处理下两个品种水稻根系H+-ATPase 活性与 CK 无差异(P>0.05),根中 ATP 含量低于CK(P<0.05)。Ca2++SAR1处理下,两个品种水稻H+-ATPase活性与CK无显著差异(P>0.05),其中在五优308中与SAR1处理无差异(P>0.05),在南粳9108中低于SAR1处理(P<0.05);两个品种水稻根中ATP含量均显著低于CK(P<0.05),在五优308中高于SAR1处理(P<0.05)。Ca2++SAR2 处理下,两个品种水稻H+-ATPase活性和ATP含量均低于CK水平(P<0.05),五优308中分别达CK水平的82%、78%,南粳9108中分别达到 CK 水平的 80%、69%,但均高于 SAR2 处理(P<0.05)。恢复期,五优 308 H+-ATPase 活性和ATP 含量在所有处理下均恢复至CK水平(P>0.05)。南粳9108在SAR2处理下H+-ATPase活性和ATP含量仍低于CK水平(P<0.05),Ca2++SAR2处理下ATP含量仍低于CK但高于SAR2处理(P<0.05),H+-ATPase活性恢复至CK水平(P>0.05)。南粳9108在其他处理下H+-ATPase活性和ATP含量均恢复至CK水平(P>0.05)。

图3 外源Ca2+对模拟酸雨下不同抗性水稻根系质膜H+-ATPase活性和ATP含量的影响Figure 3 Effects of exogenous Ca2+on H+-ATPase activities and ATP contents of different resistant rices root under simulated acid rain stress
2.4 外源Ca2+对模拟酸雨下不同抗性水稻根系质膜H+-ATPase磷酸化水平的影响
如图4所示,胁迫期,SAR1处理下,五优308根系质膜 H+-ATPase 磷酸化水平高于CK(P<0.05),南粳9108 H+-ATPase 磷酸化水平与CK 无差异(P>0.05);SAR2处理下,五优308根系质膜H+-ATPase磷酸化水平与CK无差异(P>0.05),在南粳9108中显著低于CK(P<0.05)。单一外源 Ca2+处理下,五优 308 的H+-ATPase磷酸化水平与CK无差异(P>0.05),在南粳9108中则显著增加(P<0.05)。Ca2++SAR1处理下,五优308的H+-ATPase 磷酸化水平与CK 无差异,但低于SAR1处理(P<0.05),南粳9108 H+-ATPase磷酸化水平与CK和SAR1处理均无差异(P>0.05)。Ca2++SAR2处理下,五优308中H+-ATPase磷酸化水平与CK和SAR2处理均无显著差异(P>0.05),南粳9108中H+-ATPase磷酸化水平与CK 无差异(P>0.05),但高于SAR2 处理(P<0.05)。恢复期,五优308的H+-ATPase 磷酸化水平在所有处理下均恢复至CK水平(P>0.05),而南粳9108的H+-ATPase 磷酸化水平在SAR2处理下仍低于CK(P<0.05),但其余处理均恢复至CK水平(P>0.05)。
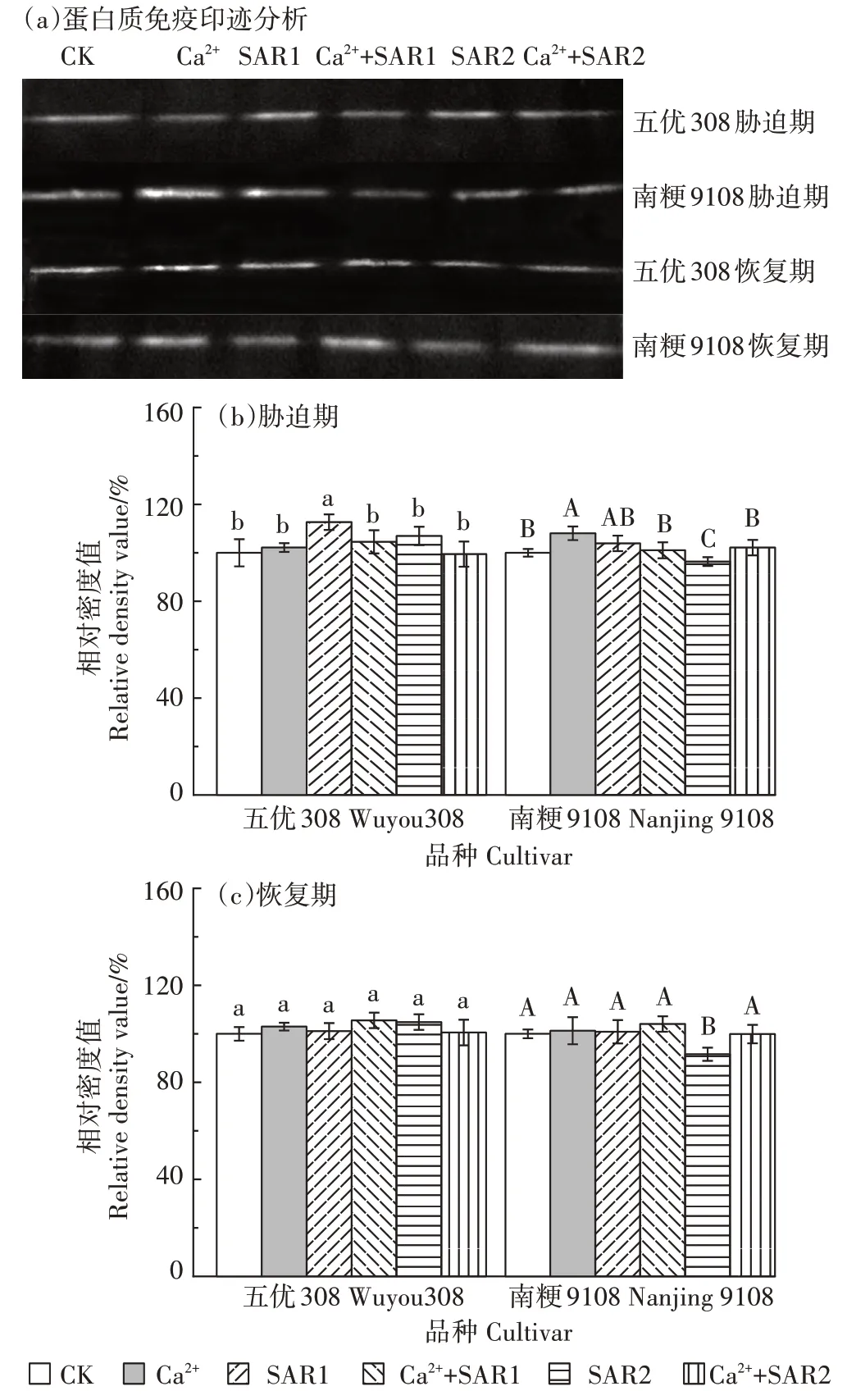
图4 外源Ca2+对模拟酸雨下不同抗性水稻根系质膜H+-ATPase磷酸化水平的影响Figure 4 Effects of exogenous Ca2+on phosphorylation levels of H+-ATPase in roots plasma membranes of rice under simulated acid rain stress
3 讨论
3.1 外源Ca2+对模拟酸雨下不同抗性水稻幼苗生长和氮吸收及供能的影响
根系是植物从环境中获取养分的重要器官,其对植株整体的生长状况有重要影响。NO-3和是植物必需的无机氮,NO-3的吸收需要H+-ATPase 水解ATP提供质子驱动力,的吸收伴随着胞内pH 增加,也需要H+-ATPase 泵出多余的H+以维持细胞pH 稳定[23],然而过量积累会引起铵毒效应[24-25]。本研究发现SAR1 处理均刺激了两个品种水稻根系H+-ATPase 活性,促进了ATP 水解,增加了供能,促进了两个品种水稻的吸收和五优308 的NO-3吸收。该处理下五优308 中NO-3和含量没有显著变化,根系生长没有受到影响,南粳9108 中含量显著增加,根系生长受到抑制。在南粳9108幼苗根系中积累,需要向胞外排出H+以维持胞内pH 稳定,导致根系环境pH 降低[26],从而加剧了对根系生长的抑制。此外,的增加破坏了水稻根系环境中阴阳离子平衡[23],这也不利于南粳9108 根系生长。SAR1 处理尽管增加了NO-3的吸收,但并未引起根中NO-3含量的显著增加,这可能是NO-3向上转运,分配到叶中的量增加所致。本研究还发现SAR2 处理抑制了两个品种水稻根系质膜H+-ATPase 活性,减少了ATP 含量,供能减少导致NO-3和吸收和积累减少,根系生长受到抑制,其中对南粳9108 的抑制程度更大。与本研究结果一致,有研究发现在盐胁迫下,低强度盐胁迫会刺激H+-ATPase 活性增加,促进营养元素的吸收,而高强度盐胁迫则抑制H+-ATPase 活性,减少对氮的吸收,从而使植物生物量减少[22]。因此推测低强度(pH 4.5)酸雨胁迫下水稻根系质膜H+-ATPase 活性的增加可能是一种适应性反应,以此提高植物对胁迫的耐受能力,高强度(pH 3.0)酸雨则超出了水稻的耐受范围,引起H+-ATPase 活性的下降。此外,铝胁迫下,耐受性品种玉米中氮素吸收受抑制程度小于敏感性玉米[27],这与本研究结果发现的酸雨胁迫下品种变化的差异类似。
研究表明外源Ca2+能缓解重金属、低氧以及盐胁迫对植物NO-3吸收的抑制,从而缓解对植物生长的抑制[20,28-29]。本研究结果显示,Ca2++SAR1 处理可使两个品种水稻根系质膜H+-ATPase 活性稳定在CK 水平,有利于分解ATP,稳定NO-3和吸收,从而缓解了根系生长受到的抑制。这与我们先前的研究结果一致,低浓度(pH 4.5)酸雨下,外源Ca2+降低大豆H+-ATPase 活性至对照水平,调节其营养吸收平衡,有效缓解了酸雨对生长的抑制[14]。本研究结果还显示,Ca2++SAR2处理对两个品种水稻根中H+-ATPase活性降低和ATP 含量减少有缓解作用,但缓解效果有限,其中对五优308 的缓解效果更好。这可能是因为Ca2++SAR2 处理对NO-3和的吸收高于 SAR2 但仍低于CK,根系生长仍受到抑制,但五优308 根系生长更接近CK 水平。研究表明胞外Ca2+的存在能增加膜厚度,降低细胞膜的流动性,提高H+-ATPase 活性[30]。低强度(pH 4.5)酸雨不改变植物根系质膜稳定性,而高强度(pH 3.0)酸雨降低了根系质膜稳定性[13]。外源Ca2+可能通过调节模拟酸雨下膜稳定性调节H+-ATPase 活性,且对SAR1 和SAR2 处理下膜稳定性的调节和对H+-ATPase活性的调节效果一致。Ca2+与模拟酸雨处理下,外源Ca2+的调控使水稻根中和NO-3含量更接近CK 水平。这与低温胁迫下外源Ca2+缓解小麦生长和氮水平下降的结果一致[31],表明外源Ca2+有利于缓解非生物胁迫对植物根系生长的抑制。本研究还发现外源Ca2+对SAR1处理下两个品种水稻的调控效果优于SAR2处理,对五优308的调控效果优于南粳9108。ZHANG 等[32]也发现外源Ca2+对盐胁迫下耐受性苦荞的调控效果更好。对比分析外源Ca2+调控作用的共性及差异性,扩大外源Ca2+调控作用的共性,合理利用品种差异性筛选耐受基因并结合外源Ca2+调控作用,将有利于缓解酸雨对农作物生长的抑制,减轻酸雨造成的危害。
恢复期结果显示SAR2 对两个品种水稻NO-3的吸收均造成了不可恢复性伤害,但外源Ca2+减小了SAR2 造成的不可恢复性伤害的程度,更有利于五优308幼苗根系NO-3吸收和根中NO-3含量的恢复。SAR2处理下,由于H+-ATPase 活性和ATP含量受到了不可逆性伤害,导致南粳9108 NO-3和吸收也受到不可逆的伤害。
3.2 外源Ca2+对模拟酸雨下不同抗性水稻根系质膜H+-ATPase磷酸化水平的影响
H+-ATPase 活性受多种方式调控,包括转录水平调控、翻译后修饰调控和信号分子调控。磷酸化是一种翻译后修饰的调控方法,通过14-3-3 蛋白与H+-ATPase C 端自抑制结构结合从而激活H+-ATPase[33]。研究表明胞外H+浓度增加会引发根系质膜H+-ATPase 的磷酸化水平增加[34],促进 H+-ATPase 活性增加。这与本研究结果中SAR1 导致水稻根系环境酸化,两个品种水稻根系H+-ATPase 磷酸化水平增加引起活性增加相同。SAR2 处理下两个品种水稻H+-ATPase活性均降低,其中南粳9108降幅更大,同时五优308 的H+-ATPase 磷酸化水平没有显著变化,南粳9108 磷酸化水平显著降低。这表明SAR2 处理下,五优308 中的H+-ATPase 磷酸化水平可能不是H+-ATPase 活性降低的主要原因。南粳9108 中H+-ATPase 磷酸化水平的降低是导致其H+-ATPase 活性降低的原因之一。高强度酸雨会降低水稻质膜H+-ATPase 10 个编码基因的转录水平,造成H+-ATPase活性降低[35]。此外,质膜H+-ATPase 作为一种膜结合蛋白,需要磷脂才能发挥活性[36]。笔者先前的研究发现,高强度酸雨会导致水稻根系质膜中总磷脂含量的降低[13],这可能也是导致H+-ATPase 活性降低的原因之一。南粳9108 中H+-ATPase 磷酸化水平的降低可能是其活性降低的原因之一,同时其转录水平、细胞膜磷脂含量可能均降低,共同引起了南粳9108中H+-ATPase 活性更大程度的降低。研究表明铝胁迫下耐受性强的蚕豆H+-ATPase 磷酸化水平高于敏感性蚕豆[37],这与本研究中高强度酸雨胁迫下耐受性水稻五优308 中H+-ATPase 磷酸化水平高于敏感性水稻南粳9108的结果一致。
本研究结果显示,Ca2++SAR 处理下外源Ca2+有利于维持水稻根系质膜H+-ATPase 磷酸化水平的稳定,从而保障其活性的稳定。研究表明,Ca2+通过磷酸化影响H+-ATPase 的活性,且这种调节作用并不依赖于钙调蛋白[38]。腺苷5′-单磷酸抑制根系H+-ATPase 磷酸化后会降低植物对铝的耐受能力[39]。而在本研究结果中,外源Ca2+可维持酸雨胁迫下水稻根系H+-ATPase磷酸化水平稳定,这可能是外源Ca2+保障酸雨胁迫下水稻H+-ATPase 活性稳定,提高水稻耐受性的原因之一。此外,本研究也发现酸雨胁迫下,外源Ca2+能缓解酸雨引起的H+-ATPase编码基因转录水平的下降[40],同时外源Ca2+还能缓解酸雨引起的质膜磷脂含量下降[13],这可能也是外源Ca2+调控H+-ATPase活性的原因。在研究两个耐盐性差异的水稻品种时也发现,Ca2+增强了耐盐性水稻根系H+-ATPase 磷酸化,而盐敏感性水稻磷酸化水平没有变化[41]。这与本研究中发现外源Ca2+对酸雨胁迫下抗性水稻质膜H+-ATPase 磷酸化水平调控效果更好的结果相似,这可能是外源Ca2+对酸雨胁迫下抗性水稻五优308 的H+-ATPase 活性调控效果更好的原因之一。恢复期,SAR2 处理下五优308 根系质膜H+-ATPase 磷酸化水平恢复至CK 水平,但南粳9108 并未恢复,这与SAR2处理下两个品种水稻根系质膜H+-ATPase 活性的恢复情况一致,因此五优308 根系质膜H+-ATPase 磷酸化水平没有遭受不可逆伤害是其活性可恢复的原因之一。外源Ca2+减小了SAR2 处理对南粳9108 根系质膜H+-ATPase 磷酸化水平的不可逆伤害程度,从而减少了对H+-ATPase活性不可逆伤害程度。
4 结论
(1)pH 4.5 模拟酸雨处理下,五优 308 和南粳9108两个品种水稻H+-ATPase磷酸化水平增加,提高了H+-ATPase 活性,促进了ATP 分解,同时也均促进了对的吸收,然而仅在南粳9108的根中过量积累从而抑制了根系生长,五优308 根中NO-3和含量没有显著变化,根系生长正常。pH 3.0模拟酸雨处理下,南粳9108的磷酸化水平下降导致H+-ATPase活性大幅降低,ATP 水解供能减少,NO-3和吸收及积累减少,根系生长减慢;而五优308 保持了H+-ATPase 磷酸化水平从而避免了H+-ATPase 活性大幅降低,且NO-3、吸收和积累及生长降幅较小。
(2)外源Ca2+能在模拟酸雨(pH 4.5和3.0)处理下保障两个品种水稻根系质膜H+-ATPase 磷酸化水平,有利于维持H+-ATPase 活性稳定,增加ATP 含量,保障H+-ATPase功能稳定,调节NO-3和吸收和积累,缓解根系生长抑制,降低模拟酸雨(pH 3.0)处理造成的不可逆伤害程度。相比于南粳9108,外源Ca2+对五优308的各项指标调控效果更好。
