运动对出生后多次异氟烷处理不同性别大鼠空间认知功能的影响及其分子学机制
2022-11-30房晓燕李萌萌郝济伟丘一诺张庆红
房晓燕,李萌萌,郝济伟,丘一诺,解 群,张庆红
2016年美国食品药品监督管理局发布药物警告:3岁以下婴幼儿接受手术或医疗操作,重复或长时间(3 h)使用全身麻醉及镇静药物,对大脑发育可能有一定影响[1]。本研究组及以往研究也证实,发育期多次异氟烷处理对大鼠成年后的认知功能有影响[2];有规律运动对大脑发育及神经功能修复可产生积极且深远的影响[3-6]。但不同性别间的认知差异尚未见报道。本研究旨在探讨规律运动对生后多次异氟烷处理后不同性别大鼠空间认知行为学的影响,并在探索其分子学机制。
1材料与方法
1.1 实验材料 生后7 d 的SD大鼠110只(北京华阜康生物科技股份有限公司,动物合格证编号:11401300035796,1140130035957, 11401300028504, 11401300037563)。按随机数字表法将大鼠分为正常对照组(n=28)、运动组(n=30)、异氟烷组(n=26)和异氟烷(n=26)运动组。新生大鼠和母鼠单笼饲养,21 d后按雌雄分笼。分别于出生后7、21和63 d测量大鼠体重,判断其生长发育状况。实验室温度(25±1)℃,相对湿度45%~55%,保持动物昼夜生物节律。行为学实验均在9:00~18:00进行。本实验方案通过医院动物伦理委员会批准,动物处置过程中所有操作符合动物伦理学标准。
1.2 异氟烷处理 异氟烷组及异氟烷运动组采用实验动物麻醉机(美国MATRX公司),以3%异氟烷(山东鲁南贝特,生产批号64140302)进行麻醉诱导,约5 min幼鼠进入麻醉状态,以1.5%异氟烷与30%氧气混合吸入维持2 h。麻醉过程中,通过检查幼鼠呼吸频率及每15 min夹尾确保其存活,ALC-HTP动物恒温系统(上海奥尔科特)维持体温在(36.5±0.5)℃,开启二氧化碳吸附器吸收废气[2]。异氟烷处理后幼鼠,各组取1只进行动脉血气分析,排除麻醉动物可能出现的高碳酸血症等因素对研究结果的干扰。异氟烷处理后其余幼鼠从诱导箱中取出, 3~5 min恢复爬行,待完全清醒后统一送回动物房饲养。
1.3 大鼠运动方案[7,8]运动组及异氟烷运动组从21 d开始进行有规律运动,训练时间为14:00-17:00,应用XR-PT-10A小动物跑台(上海欣软信息科技有限公司)。跑台由相互间隔的6条跑道组成,每条跑道有效空间为600 mm×95 mm×115 mm,跑台0°倾角不变,速度从4 m/min开始,每周增加2 m/min;6周速度增至14 m/min,每天训练30 min,每周5 d,剔除因受伤等未完成6周训练的大鼠。
1.4 八臂迷宫实验 运动结束后,各组大鼠进行八臂迷宫实验,亦称为辐射迷宫(radial arm maze),用于评估动物空间记忆能力[9]。实验所用八臂迷宫425 mm×145 mm× 225 mm。八臂迷宫实验期间,大鼠进行饲料限量喂养,不限水摄入,使其保持饥饿状态,体重维持在基础体重80%~85%。实验步骤分迷宫适应阶段、训练阶段、测试阶段。记录训练阶段完成实验的总耗时,即大鼠吃完所耗时间或10 min测试结束。测试阶段记录参考记忆错误、工作记忆错误、总耗时。
1.5 Morris水迷宫实验[10]本实验采用的水迷宫为直径150 cm,高60 cm的圆形水池,水深约35 cm,在其中一个象限中央放置平台(直径11 cm,高33 cm),平台于水下2 cm,实验过程通过水迷宫上方的摄像机进行录制。实验步骤分为定位航行实验、空间探索实验及反向空间实验,记录大鼠在目标象限耗时、目标象限潜伏期以及平台潜伏期。
1.6 免疫组化 各组大鼠在所有行为学试验结束后,腹腔注射3%戊巴比妥(5 mg/kg),待麻醉后,开胸以4%多聚甲醛经左心室进行全身灌注固定,完整取出大鼠海马组织,进行免疫组化染色。各指标的积分光密度值均应用麦克奥迪数码医学图像分析系统获得。
1.7 蛋白质印迹法(Western-Blot) 海马组织取材液氮中保存,随后进行蛋白质印迹实验。

2结 果
2.1 一般情况 饲养及运动训练过程中剔除因受伤或拒绝运动大鼠18只(雄鼠11只,雌鼠7只),每组大鼠为10~15只。随日龄增长,各组SD大鼠体重增加,组间比较差异无统计学意义。异氟烷处理后,各组幼鼠动脉血气分析结果比较,差异均无统计学意义。脑组织取材后称重,各组大鼠脑部重量比较无统计学差异(表1)。
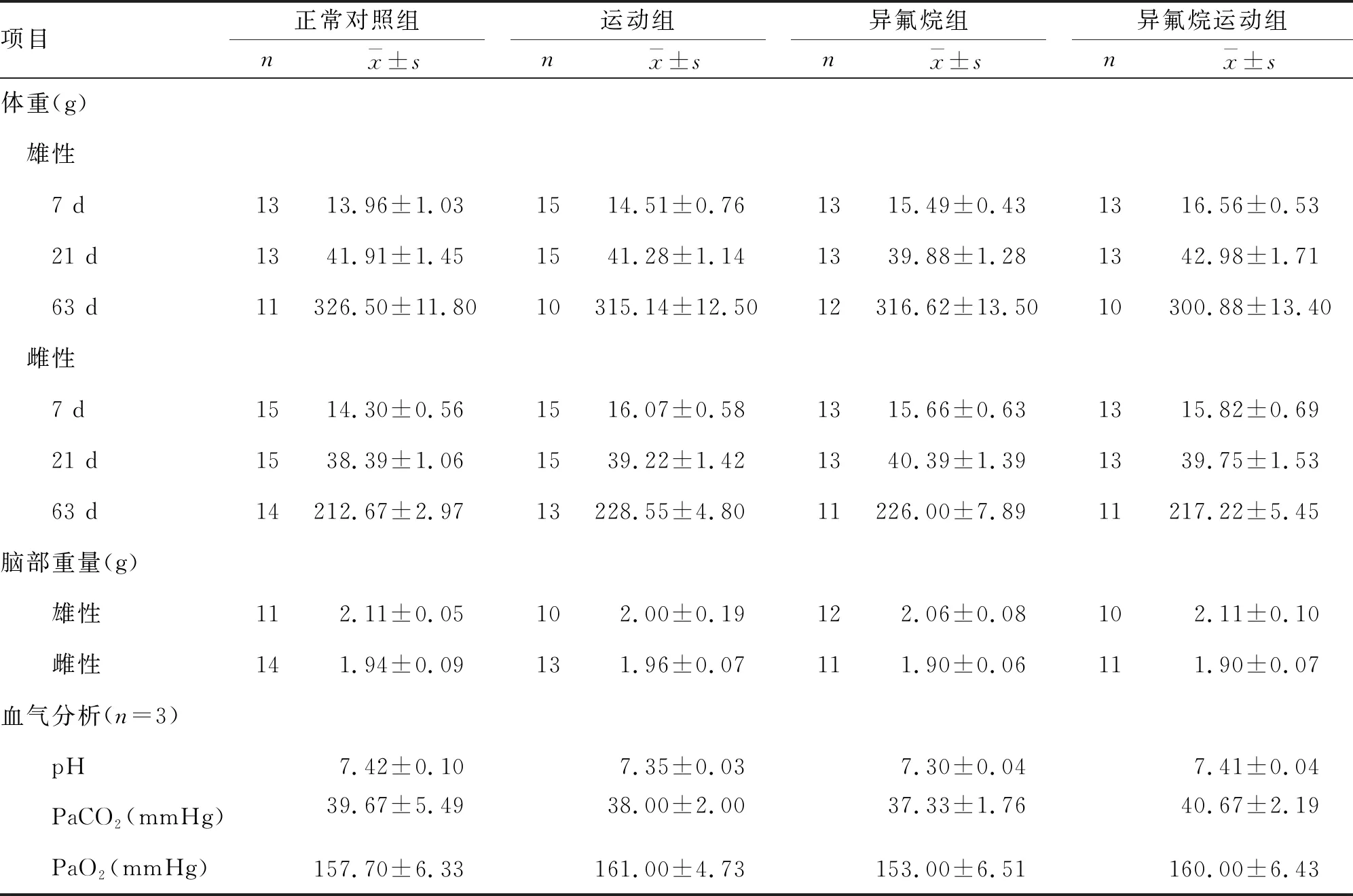
表1 各组SD大鼠一般情况
2.2 八臂迷宫实验 雌、雄大鼠在八臂迷宫训练阶段的总耗时,随训练次数增多呈现逐渐下降趋势,差异具有统计学意义(P<0.05);运动组雌、雄大鼠的表现明显优于非运动组,差异均具有统计学意义(P<0.05);尽管非运动组大鼠训练10次也能达到测试标准,但学习次数与总耗时之间无稳定的学习进步曲线,而进行规律运动的大鼠,其学习进步曲线清晰,尤其是雄鼠在训练第2次基本达到测试标准,雌鼠在第3,4 次逐渐稳定(图1A,B);异氟烷处理不同性别大鼠间比较,雌鼠在第5次达到测试标准,明显早于雄鼠(图1C);规律运动后发现,雄鼠在第2次即达到测试标准,而雌鼠在第4次训练才达到(图1D)。提示,异氟烷处理可能对雄鼠学习能力有一定影响,但规律运动后明显改善,即雄鼠的运动获益较大。

图1 运动对不同性别SD大鼠成年后在八臂迷宫训练阶段的影响
八臂迷宫测试中,采用单因素方差分析雄鼠各组间参考记忆错误及总耗时组间比较,差异具有统计学意义(P<0.001);进一步两两比较(Tamhane)发现, 与非运动组相比,运动两组参考记忆错误次数及总耗时减少,差异具有统计学意义(P<0.05,表2);分析雌鼠参考记忆错误及总耗时,组间比较均具有统计学差异(均为P=0.000);两两比较发现,与正常对照组相比,异氟烷组的参考记忆错误次数及总耗时显著增多,差异具有统计学意义(P<0.001),运动组参考记忆错误次数及总耗时降低,与正常对照组相比,差异无统计学意义;另外,异氟烷运动组与异氟烷组相比,在参考记忆错误次数及总耗时明显降低,差异具有统计学意义(均为P=0.000,表2)。
表2 运动对SD大鼠成年后八臂迷宫测试阶段的影响

表2 运动对SD大鼠成年后八臂迷宫测试阶段的影响
注:与正常对照组比较,①P<0.05;与异氟烷组比较, ②P<0.01
项目正常对照组运动组异氟烷组异氟烷运动组雄性 例数11101210 参考记忆错误(次)2.68±0.122.06±0.19①3.05±0.162.45±0.15 工作记忆错误(次)0.27±0.060.2±0.070.25±0.070.31±0.08 总时间(s)61.39±2.648.8±3.22①70.97±4.8250.1±2.89②雌性 例数14131111 参考记忆错误(次)2.4±0.092.08±0.133.09±0.11①2.17±0.15② 工作记忆错误(次)0.23±0.050.2±0.060.15±0.050.20±0.06 总时间(s)57.7±1.9752.5±2.1768.2±2.88①52.7±2.67②
2.3 Morris水迷宫实验 与正常对照组相比,异氟烷组雄鼠在目标象限潜伏期及平台潜伏期有延长趋势但差异无统计学意义;异氟烷组雌鼠在水迷宫空间探索实验的表现无明显差异;与非运动组相比,运动两组雄鼠在目标象限耗时增加,目标象限潜伏期及平台潜伏期均缩短,但无明显统计学差异(表3);反向空间实验中,异氟烷组雌鼠与正常对照组雌鼠相比,目标象限耗时减少(P<0.05,表4)。
表3 运动对SD大鼠成年后水迷宫空间探索实验的影响

表3 运动对SD大鼠成年后水迷宫空间探索实验的影响
分组正常对照组运动组异氟烷组异氟烷运动组雄性 例数11101210 目标象限耗时25.42±2.730.03±2.2426.08±2.4130.65±2.3 目标象限潜伏期4.65±0.533.91±0.665.48±1.924.22±0.83 平台潜伏期13.93±3.6010.17±2.3416.39±3.566.47±1.30雌性 例数14131111 目标象限耗时25.17±2.5729.88±2.9626.89±1.8124.12±3.01 目标象限潜伏期4.41±0.824.81±0.473.93±0.826.54±1.00 平台潜伏期9.02±2.0513.66±3.029.71±2.3614.84±3.72
表4 运动对SD大鼠成年后水迷宫反向空间实验的影响

表4 运动对SD大鼠成年后水迷宫反向空间实验的影响
注:与正常对照组比较,①P<0.05
分组正常对照组运动组异氟烷组异氟烷运动组雄性 例数11101210 目标象限耗时21.69±1.8620.95±2.821.17±1.6220.36±1.78 目标象限潜伏期4.09±0.313.34±0.544.15±0.343.81±0.43 平台潜伏期23.84±8.2720.1±8.5431.55±8.7623.75±7.22雌性 例数14131111 目标象限耗时25.58±1.1621.00±2.019.97±2.2①16.78±1.28 目标象限潜伏期4.55±5.244.9±0.755.46±1.115.24±0.91 平台潜伏期24.22±5.6227.03±6.6416.09±3.2229.13±4.83
2.4 运动对多次异氟烷处理后不同性别大鼠海马Ki-67,BDNF表达的影响 与非运动组雄鼠比较,运动组和异氟烷运动组雄鼠海马齿状回中可见Ki-67和BDNF阳性表达增多(图2A)。与异氟烷组相比,正常对照组及运动组雌鼠海马齿状回中均有Ki-67的阳性表达,而异氟烷组未见K-i67阳性细胞表达(图2B)。Western-Blot分析发现:与正常对照组相比,异氟烷组中雄鼠BDNF阳性表达水平较低;与异氟烷组相比,异氟烷运动组雄鼠BDNF表达增多;未见明显差异。与正常对照组相比,异氟烷组雌鼠中BDNF阳性表达水平均降低(P<0.05);与异氟烷组相比,异氟烷运动组雌鼠BDNF表达增多(P<0.05,图2C、D,表5)。

图2 运动对雄、雌SD大鼠海马组织Ki-67,BDNF表达的影响
表5 运动对SD大鼠海马组织BDNF表达的影响
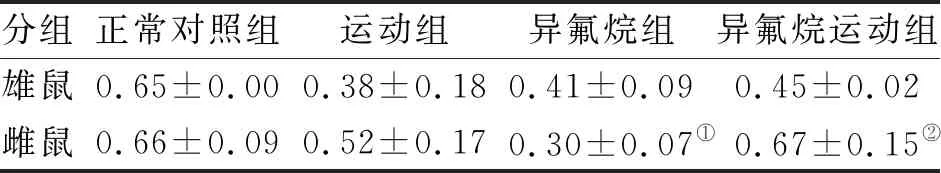
表5 运动对SD大鼠海马组织BDNF表达的影响
注:与正常对照组比较,①P<0.05;与异氟烷组比较,②P<0.05
分组正常对照组运动组异氟烷组异氟烷运动组雄鼠0.65±0.000.38±0.180.41±0.090.45±0.02雌鼠0.66±0.090.52±0.170.30±0.07①0.67±0.15②
3讨 论
有研究报道,全身麻醉药物对发育期大脑可产生短暂或长期影响,称为麻醉而致神经发育毒性(anesthesia-induced developmental neurotoxicity, AIDN)[11],其影响与大脑发育阶段、药物剂量、暴露时间及次数有关。发育早期(4岁以内)经历多次麻醉(3次)及手术暴露的儿童,认知损害风险增高[12]。但全身麻醉药物对发育期不同性别间动物认知功能的影响鲜少报道。以往基础研究多选用雄性动物,以避免雌孕激素水平波动而干扰实验结果,但认知水平的发展具有明显性别差异[13]。本研究发现,异氟烷多次处理对大鼠成年后空间认知功能有一定影响,学习能力降低,雄鼠表现在长期记忆,雌鼠表现在工作记忆方面。同时发现,发育期规律运动能够促进大鼠空间认知功能的发育,且雄鼠的运动获益强于雌鼠。English等[14]也指出,在外周神经损伤模型中,仅雄鼠在持续2周运动训练后轴突再生明显;Thompson等[15]在大鼠脑损伤研究中再次证实,雄鼠在持续平台训练的运动获益优于雌鼠,相关机制有待进一步研究。
3.1 异氟烷处理对大鼠成年后认知功能的影响 出生后7 d大鼠脑组织处于发育高峰,神经元树突、轴突及神经突触可塑性较强[2]。本研究发现,多次异氟烷处理对大鼠的空间记忆(八臂迷宫及水迷宫)有一定损害,尤其在八臂迷宫实验的训练阶段,不同性别异氟烷处理组大鼠学习能力均有所下降,尽管训练10次也达到测试标准,但学习次数与总耗时之间无稳定的学习进步曲线;而进行规律运动大鼠,其学习进步曲线清晰,尤其是雄鼠在训练第2次即达到测试标准。Cao等[16]也发现,老龄鼠暴露在1.2%异氟烷后2 h,可导致其学习过程延长,认知功能降低。另外还发现,异氟烷处理后大鼠参考记忆错误及总耗时均有所增加,提示大鼠的空间记忆中的长期记忆受到影响。随后研究发现,异氟烷处理后成年大鼠海马齿状回BDNF阳性表达均减少,提示发育期异氟烷暴露对随后大鼠的空间记忆编码,联想记忆等有一定影响[17-19]。
3.2 运动对异氟烷处理大鼠成年后认知的影响 运动已被证实能够促进其认知功能,能够促进大脑发育,智力水平,学习能力、执行和阅读能力[20-23],以发育期尤为显著。其机制与运动促进神经营养因子的表达及神经形成,神经细胞增殖、树突分枝形成、突触发生、长时程增强(long-term potentiation,LTP)和神经营养因子(BDNF)水平升高等相关[5,6,24,25]。本研究发现:发育期进行运动的两组大鼠在训练阶段,运动促进大鼠在八臂迷宫中的表现,总耗时呈稳定进步趋势,学习进步曲线稳定,且雄鼠获益更明显;然而,水迷宫的行为学测试中未发现运动优势。尽管两种行为学研究工具均是评测大鼠的空间认知功能,但对于不同性别间的研究结果差异尚需进一步探讨。
3.3 运动对异氟烷处理大鼠成年后海马的影响 大脑海马是与记忆相关的重要脑区,在空间记忆编码、联想记忆及学习能力方面起重要作用[18,19],运动引起海马改变与空间记忆任务中表现提高直接相关。本研究免疫组化结果发现,特异性海马增殖指标Ki-67,在雌雄大鼠运动两组(运动组及异氟烷运动组)的海马齿状回中表达增多,提示该区域细胞增殖活跃。与Sakalem 等[26]研究一致,运动及环境富集刺激可提高海马细胞和神经元的总体数量,从而提高空间记忆能力。大鼠在发育期进行规律运动,运动强度逐渐增加至中等强度,对其成年后认知功能有一定促进作用,与异氟烷组比较,异氟烷运动组大鼠BDNF表达也呈现增多趋势。进一步证实,运动改善认知能力与BDNF水平升高相关。
此外,我们将异氟烷和运动对大鼠成年后认知的影响分为学习和记忆两个阶段分别进行分析,发现异氟烷处理影响大鼠的学习能力,与梅奥医学中心发表的MASK临床研究结果相呼应[27],该研究发现多次接受全身麻醉的患儿的问题处理速度和精细运动能力下降。本研究发现运动能够改善异氟烷所造成的损害,这可能为未来进行认知功能改善提供参考。
综上,多次异氟烷处理对大鼠成年后学习认知功能有一定影响,且存在性别差异;雄鼠在长期记忆,而雌鼠在工作记忆方面有一定损害。同时发现,运动促进大鼠学习认知功能的发育,且雄鼠的运动获益更为明显。
