氮代谢参与植物低氮胁迫研究进展
2022-11-30李佳佳徐翎清芮秀丽石俊婷刘大丽
李佳佳,徐翎清,赵 阳,芮秀丽,石俊婷,刘大丽
(1国家甜菜种质中期库/黑龙江大学,哈尔滨 150080;2黑龙江省普通高等学校甜菜遗传育种重点实验室/黑龙江大学现代农业与生态环境学院,哈尔滨 150080)
0 引言
植物生长依赖于无机氮,据报道,全世界每年向土壤中添加约8500万~9000万t氮肥[1]。人们试图通过施加大量的氮肥来实现作物高产,但事实上只有30%~40%的氮素会被植物吸收利用[2]。随之而来的是,未被利用的释放到环境中的含氮化合物会造成环境污染,如土壤酸化、土壤硬化或水的富营养化等[3]。那么,如何适量施氮,如何提高氮肥的有效利用率,如何在较低的氮水平下维持作物的正常生长,是急需解决的问题。因此,研究农作物在低氮胁迫下的响应机制是十分必要的。同时,植物对低氮胁迫氮代谢过程中的响应过程和适应机制也一直是学术研究的热点领域。提高氮肥的使用效率和限制氮肥的过度使用是保护环境和改善可持续生产性农业的一个重要挑战。
1 氮代谢概述
1.1 氮代谢概念
氮代谢是指氮的吸收、还原、氨基酸的代谢和运输,以及氮的转运和再利用等过程。它是植物的基本生理过程之一,也是植物参与地球化学循环的重要组成部分[4],对于植物抵抗逆境胁迫具有重要的意义。氮代谢与植物的生长和发育以及产量密切相关。
1.2 低氮胁迫过程中的氮素同化途径
氮的吸收、同化和再活化的分子机制已被广泛研究。植物对氮素的利用,包括吸收、易位、同化等过程,并受到遗传和环境因素等多种因素的控制[5]。植物氮的同化取决于谷氨酰胺合成酶-谷氨酸合成酶循环(GS/GOGAT)以及可用的能量和碳骨架。当氮供应不足时,植物的光合作用和呼吸作用受阻,植物自身补偿机制被相应的激活[6]。CURCI等[2]发现,在低氮胁迫下,参与小麦光合作用和光呼吸的大多数基因均被下调,包括光系统(PSI)、光系统(PSII)、ATP合酶和Rubisco(ribulose-1,5-bisphosphate carboxylase/oxygen ase)。研究表明,在低氮胁迫下,由于细胞色素中心吸收的激发能被耗散,PSII光化学效率随之降低[6]。低氮水平下,氮通过三羧酸循环同化进入氨基酸代谢途径,细胞质和线粒体经呼吸途径氧化产生更多的2OG(2-oxoglutarate)[7],为铵的再同化提供GS/GOGAT循环[8],来增强水稻对低氮胁迫的适应能力[9]。LAURENT等[10]也发现低氮胁迫时,2-OG可以在细胞内短暂积累,并作为氮缺乏时的信号。为达到最佳的氮同化速率,植物需要较高的光呼吸速率,通过叶绿体被膜或线粒体的苹果酸/草酰乙酸循环加强向细胞膜的氧化还原转移[7]。研究发现,呼吸速率与植物氮浓度呈正相关[11]。植物线粒体呼吸作用的底物供应与光合作用产生的碳水化合物有关[11]。硝酸盐还原酶(NR)和亚硝酸盐还原酶(NiR)将植物细胞吸收的NO3-还原成NH4+,在质体或叶绿体中被GS/GOGAT循环同化为谷氨酸。PSII还可以与2-OG形成复合物,并通过蛋白-蛋白相互作用相应地调节氮的同化[12]。其基本过程如图1所示。此外,研究还发现拟南芥和黄瓜在低氮胁迫下,参与光合作用的相关基因显著下调,进一步说明了光系统对低氮胁迫十分敏感[13-14]。
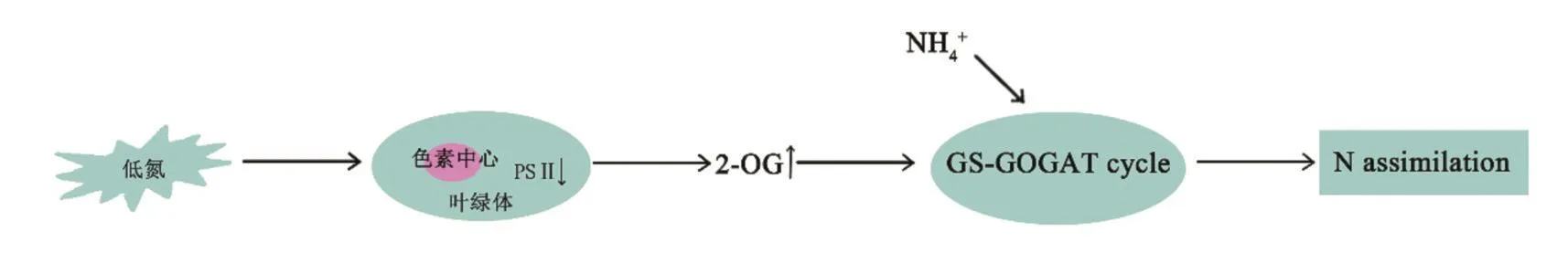
图1 植物体内基本氮代谢途径
2 植物应对低氮胁迫的研究进展
2.1 外部形态
根系不仅是植物吸收营养和水分的重要器官,也是植物激素和有机酸等许多物质的同化、转化和合成的重要场所[15]。当施入的养分不能满足植株生长要求时,植株本身会根据需要的氮量调整其根系结构,作为生存策略的一部分来应对生境变化[16]。JU等[17]认为氮高效水稻品种的根系生物量更大,根分布更深,更长,根密度更大。轻度缺氮可促进根系生长,有利于根系深层渗透;适当供氮可增加根系数量、生物量和密度;但过量供氮可抑制根系生长[18-19]。拓扑结构可以将根系分为很多类型,比如人骨类型,其只包含主轴和侧根的分支模式[20]。缺氮的幼苗会将其原有拓扑结构转化为二分体状根系[21],具有短、细、高发育等级的侧根[20]。本课题组前期等发现低氮胁迫会使甜菜植株生长势弱,不定根数量增多,叶色黄绿且薄,且推断甜菜可以通过改变根系构型和根系比例来获得氮素平衡[22]。
2.2 氮代谢相关酶
(1)硝酸还原酶
硝酸还原酶(nitrate reductase,NR)是植物氮同化的第一关键酶,它可以直接调控硝酸盐的还原[23],其活性高低与植物自身氮同化能力密切相关,因此NR的活性越强,植物的氮素代谢也越旺盛[24]。NR还是光合自养生物体内的一种诱导酶[25],光可诱导NR磷酸化,光度降低会导致NR活性迅速下降,进而降低植物对硝酸氮的吸收和同化[26]。低氮胁迫下,NR活性高的植株其耐低氮的能力较强[27]。高活性NR对植物生长发育和产量有积极的影响。
(2)谷氨酰胺合成酶
谷氨酰胺合成酶(GS)广泛存在于所有生物中,它利用ATP水解到ADP释放的能量催化氨和谷氨酸合成谷氨酰胺,是生物氮代谢的另一关键酶[28],在低氮胁迫中发挥重要作用。谷氨酰胺合成酶主要以两种不同亚细胞定位的同工酶形式存在:细胞质GS1和叶绿体/质体GS2[29]。GS1对植物正常的生长发育十分重要[30],GS2对叶绿体中N的光呼吸代谢十分重要[31]。GS在植物氮代谢途径中的关键作用已得到较为深入的研究,总体表现为不同的同工酶表达类型和生物学功能[32]。大量研究表明,GS可以调节植物的含氮量,间接影响光合作用和生理代谢,从而影响生长发育以及作物产量和品质[33]。
(3)谷氨酸合成酶
谷氨酸合成酶(glutamate synthase,GOGAT)是氨同化途径中的一种关键酶,能够与GS结合,完成氨同化。在高等植物中,主要的GOGAT同工酶是非光合作用组织中的NADH-GOGAT和光合作用组织中的铁氧化还原蛋白-GOGAT(Fd-GOGAT)[5]。后者占GOGAT总活性的95%,在植株叶片中处于主导地位[32]。Fd-GOGAT存在于叶绿体中,光诱导其生物合成,并同时与光合作用和光呼吸密切相关[34]。研究发现,低氮胁迫时草地早熟禾的NADH-GOGAT基因的表达量会增多[35]。GOGAT基因的表达受氮素、光照、作用部位等多种因素的影响[36]。
(4)谷氨酸脱氢酶
谷氨酸脱氢酶(glutamate dehydrogenase,GDH)是生物体中的一种多聚酶[37]。由于GDH催化的氨基酸脱氨作用产生2-酮戊二酸进入GS/GOGAT循环以提供底物,因此GDH在GS/GOGAT循环中起补偿[38]以及缓解铵中毒的作用[39]。GDH可以催化谷氨酸的合成,增加脯氨酸的数量,来提高植物的耐逆性[40]。
2.3 低氮胁迫对植物体初级代谢的影响
碳、氮代谢是植物最基本的两条代谢途径[41]。植物碳氮代谢的动态变化直接影响着光合产物的形成和转化,以及矿质营养物质的吸收和蛋白质的合成。氮是叶绿素分子的重要组分。氮的减少会导致基质蛋白和类囊体蛋白的减少,抑制叶绿素的合成,从而影响光合作用[42]。通过光合作用对氮的吸收和同化[43],确保了氮的吸收与碳的状态相关。碳代谢为氮代谢提供了碳源和能量[44],而氮代谢为碳代谢提供了酶和光合色素。植物的碳氮代谢彼此密切协调[45],需要共同的还原力、ATP和碳骨架[46]。
维持碳氮代谢的协调,以及碳水化合物与氮代谢物的适当平衡,也被称为“C/N平衡”,对植物的生长、发育和产量非常重要[47]。RuBisCo占C3植株叶片中可溶性蛋白总量的50%,占C4植株叶片中总量的20%[48]。在C3植物中,RuBisCo的氧化导致二氧化碳和光呼吸氨的释放[49]。研究发现RuBisCo和2-酮戊二酸(2-oxoglutarate)分别作为碳和氮饥饿信号[10],它们的浓度比可能间接反映C/N代谢平衡的状态[50]。光呼吸和三羧酸循环(TCA)不仅是重要的代谢途径,而且在C/N代谢控制的信号传导中起关键作用[50]。XIN等[9]发现,TCA循环可以使各种氨基酸的氮分配趋向合理,可提供更多的2-OG为GS/GOGAT循环提供NH4+,从而增强水稻对低氮胁迫的适应能力。在长时间缺氮条件下,由于RuBisCo的分解,光合活性下降,叶片衰老普遍加速,植物生长最终受到抑制[51]。双标记(13C/15N)硝酸铵和核磁共振(NMR)分析表明,甘蓝型油菜用于白天GS/GOGAT合成的α-酮戊二酸(2-OG)来源于夜间储存的有机酸(可能是苹果酸或柠檬酸),因此碳代谢对氮同化具有重要意义[52]。ABC1/OsFd-GOGAT通过调节氮同化和C/N平衡,在水稻的生长发育中发挥着至关重要的作用[8]。因此,调控碳氮平衡可以促进含碳、氮营养物质的吸收、同化和代谢效率,对提高低氮水平下作物产量具有指导意义。
2.4 低氮胁迫对植物体次级代谢的影响
当植物受到非生物胁迫时,体内会积累活性氧物质(ROS),并对植物细胞造成严重损害。然而,植物可以通过非生物胁迫诱导的抗氧化酶系统和次生代谢产物来消除过量的ROS,从而保护植物免受胁迫。苯丙烷代谢途径是重要的次级代谢途径之一。苯丙烷代谢物的生物合成和多样性是通过加氧酶、氧化还原酶、连接酶以及多种转移酶实现的,这些酶可通过酰化、甲基化、糖基化和羟基化对代谢物的基本骨架进行化学修饰[53]。苯丙氨酸的合成来源于莽草酸,莽草酸是高等植物次生代谢的起始物[54],合成许多重要的芳香代谢物。因此,苯丙烷代谢途径跨越初级代谢和次级代谢的边界,并引导代谢通量从初级代谢转向次级代谢[55]。苯丙烷类衍生物代谢途径有两条重要的子途径:木质素生物合成途径和类黄酮生物合成途径[56]。
类黄酮代谢是苯丙素代谢的一个重要分支,产生了最大的一类多酚类代谢物,大约包含6000多个化合物[57-58],并可受到C代谢诱导[59]。黄酮类化合物是花、果实和种子的主要着色物质,在植物生长和植物-环境相互作用中起着重要作用[60],且分布广泛,可作为抗氧化剂、抗菌物质、植物与细菌、光受体、色素相互作用中的信号分子[61-62]。研究发现,在豆科植物中,缺氮可诱导黄酮类化合物和异黄酮类化合物作为信号,抑制Nod因子生物合成相关基因的转录[63]。SiWLIM2b基因的表达可以通过增加其次级代谢产物类黄酮的含量来影响苯丙烷次级代谢途径,从而增强转基因水稻的抗旱性[64]。在茶和苜蓿中,bHLH35与苯丙氨酸生物合成和类黄酮生物合成途径中的许多基因表达呈正相关[65-66]。
苯丙素代谢途径与木质素的合成密切相关[67],而木质素的生物合成途径与光合碳的利用密切相关[68]。木质素等苯丙类聚合物是植物生长的机械支撑,促进水和营养物质的长距离运输[69]。它们还有助于植物对非生物和生物胁迫的应答,并促进昆虫作为授粉和种子传播媒介的吸引[70]。在低氮条件下,光合作用中的碳可以通过糖酵解或磷酸戊糖途径(PPP)转移到木质素的合成中[9]。因此,苯丙烷代谢物对植物的生长发育至关重要。
2.5 氮代谢相关基因参与低氮胁迫的研究进展
WRKY是水稻中对氮缺乏反应最强烈的转录因子家族之一,研究发现,水稻中有12个WRKY家族成员在叶鞘中诱导表达[71]。而低氮胁迫下,WRKY在马铃薯幼芽中却呈现下调表达[72]。因此WRKY作为传递氮胁迫耐受的关键因子,其参与的信号网络十分复杂,并在植物胁迫反应中起着重要的调节作用。LIANG等[73]发现,在缺氮水平下,参与苯丙烷生物合成和谷胱甘肽代谢途径的相关基因发生显著变化,如氮 转 运 体(NRT2.1、NRT2.2、NRT2.3、NRT2.4、NAR2.1、AMT1.3、AMT1.2和推测的AMT3.3)和氮同化体(NR2、GS1.1、GS1.2、GS1.3、NADH-GOGAT2和AS2)。SULTANA等[74]研究发现,小麦在氮胁迫下,下调的差异表达基因DEGs主要与碳水化合物代谢过程、光合作用、光捕获和防御反应有关,而上调的DEGs与核苷酸代谢、蛋白质水解和跨膜转运有关。氮饥饿条件导致根系总氮含量显著降低,在高亲和力的NRTs中,与拟南芥NRT2.5同源的基因在硬粒小麦中显著上调[2]。作为基因表达的调控元件,转录因子在外部环境中的氮浓度感知、氮信号转导和氮代谢的调控途径中发挥着重要作用。
3 展望
本研究综述了植物在应对低氮胁迫过程中所表现的形态、生理以及分子等方面的响应机制。为了提高农业生产的可持续性,适量的氮肥可以改善寒冷地区盐碱地土壤的理化性质,促进土壤微生物的生长,从而改善土壤质量。这种高效的“资源利用效率”需要对植物生理、植物吸收能力、植物代谢、植物对胁迫的响应以及土壤物理和化学性质等方面进行深入的研究。随着高通量测序技术、高分辨率质谱技术和信息处理技术的发展和应用,系统生物学研究已占据探索重要生物现象的主导地位。因此,本研究提出几点植物面临缺氮条件下未来的研究方向:(1)植物在低氮条件下的代谢网络不仅错综复杂,而且在植物之间、同一植物的不同生长期都存在着显著的差异;(2)转录组学已成为后基因组时代研究植物抗性最主要的学科之一。转录组学研究已经鉴定出大量基因在低氮胁迫下mRNA表达水平发生改变,但这些基因的表达产物之间的关系有待进一步研究;(3)科学家们可以从转录组学、功能基因组学、遗传学、生态生理学和土壤科学等角度来突破,系统研究植物对逆境胁迫后各种指标的变化,以期全面了解植物对低氮胁迫的分子响应机制。
