基于生长性状育种值和遗传多样性构建福建柏第1代核心育种群体
2022-11-29苏顺德
苏顺德
(福建省林业科学研究院 国家林业和草原局南方山地用材林培育重点实验室,福建省森林培育与林产品加工利用重点实验室,福建 福州 350012)
福建柏(Fokieniahodginsii)为柏科福建柏属唯一种,是我国第一批珍稀濒危二级保护植物,也是我国珍贵用材树种[1]。主要分布于福建、江西南部、浙江南部、广东北部、广西北部、云南中部及东南部、湖南南部、贵州和四川南部以及越南北部。水平呈东西窄条状分布区,垂直分布于海拔150~1 200 m[2]。福建柏是优良的用材树种,也是营建混交林的优良树种,具有生长快,适应性较强,材质优良的特点[3-5],且在种源间、家系间具有丰富的遗传变异6-8],并受到中度的遗传控制[9-14]。虽然在种源试验和优树自由授粉子代测定试验基础上,福建柏当前已完成种源筛选[12]、优树选择和第1代种子园营建[13,15-16],但关于育种群体的建设未见报道。育种群体是根据育种目标,从选择群体中筛选出的具有高遗传增益,负载较丰富遗传多样性,且相互间亲缘关系清晰可控的群体,是开展交配创制种质的主要群体,持续推动育种群体建设及其相关交配设计和杂交、测定工作是最为有效的林木育种途径[17]。国内外对主要树种均组建了育种群体[18-21]。得益于分子标记技术,依据表型遗传多样性分型和分子遗传距离分型构建具有亚结构的育种群体快速发展,如李梅[22]利用RAPD分子标记构建亚系结构的杉木育种群体,冯源恒等[23-24]利用SSR分子标记构建主群体——核心育种群体结构的马尾松第1代和具有10个亚系结构的第2代育种群体。因此,为了提高种质创新效率,持续推进福建柏轮回选择育种,本研究基于20年生优树自由授粉子代测定结果和优树EST-SSR分子遗传距离,构建了福建柏第1代核心育种群体,为交配制种提供亲本材料。
1 试验点概况
优树自由授粉子代测定林营建在福建省永安国有林场(福建省三明市永安市,福建中部)和福建省仙游溪口国有林场(福建省莆田市仙游县,福建东南部)。永安国有林场的试验林设在永浆工区,117°23′E,25°57′N。海拔195~250 m。山地红壤,Ⅲ类立地。前作为马尾松人工纯林。中亚热带海洋性季风气候,年平均降水量1 600 mm,年平均相对湿度83.0%,年平均气温19.3 ℃,极端高温40.5 ℃,极端低温-11.3 ℃,年平均无霜期295 d。仙游溪口国有林场的试验林设在场部管理区,118°57′E,25°37′N。海拔180~250 m。山地红壤,Ⅱ类立地。前作为杉木人工纯林。南亚热带海洋性季风气候,年平均降水量1 536 mm,年平均相对湿度76.4%,年平均气温20.0 ℃,极端高温39.4 ℃,极端低温-3.5 ℃,年平均无霜期318 d。
2 材料与方法
2.1 选择群体
选择群体为福建柏优树。1998-1999年在福建全省和湖南省道县选择优树175株[12]。1999年采种,2000年1月播种培育出127个家系足量苗木。根据各家系苗木数量,将优树子代测定试验分为试验1和试验2。试验1参试家系80个,以福建仙游福建柏母树林混种苗木(CK1)为对照。试验2参试家系47个,以福建仙游福建柏母树林混种苗木(CK1)和福建大田福建柏人工林混种苗木(CK2)为对照。
2.2 育种值测定方法
以优树自由授粉子代测定生长性状育种值,田间试验设计均为完全随机区组设计(RCB)。试验1参试处理81个(含CK1),10次重复,5株单列小区;试验2参试处理49个(含CK1和CK2),8次重复,6株单列小区。初植密度2 500株·hm-2,株行距2.0 m×2.0 m,穴规格50 cm×40 cm×30 cm,挖明穴,回表土,不施基肥。试验1在2个试验点均实施,试验2只在永安国有林场实施。2001年2月1 a生苗木造林,苗木平均苗高19.5 cm,平均地径0.30 cm。造林后前3 a每年春秋2季各锄草1次,之后每年春秋2季各劈草1次至第6年。2020年底(20 a林龄)每木调查树高(H)和胸径(D)。单株立木材积(V)估算[12]:V=0.000 056 85D1.629 996H1.261 954。
2.3 DNA提取方法
取样品的叶片在液氮中研磨粉碎,采用DNAsecure Plant Kit新型植物基因组DNA提取试剂盒(天根生化科技有限公司)提取各优树基因型DNA,并用1%琼脂糖凝胶电泳对样品的合格性进行检测。
2.4 PCR反应方法
利用筛选的11对EST-SSR引物(表1)对DNA样品PCR扩增。PCR试验采用50 μL体系,配比为:Dream Taq Green PCR Master Mix (2x)25 μL(赛默飞世尔科技公司Thermo Fisher Scientific)、F端引物1 μL、R端引物1 μL、DNA模板2 μL、ddH2O 21 μL。PCR反应程序为:94 ℃预热2 min,94 ℃变性20 s,52 ℃退火20 s,72 ℃延伸10 min,变性-退火-延伸设置40个循环。通过2%TAE琼脂糖凝胶进行电泳检测PCR产物。

表1 PCR扩增所用11对微卫星引物
2.5 统计分析方法
2.6 第1代核心育种群体构建方法
依据单株立木材积育种值和EST-SSR分子标记测定的遗传距离,从后向选择出的优树中利用改进的LTTB(largest triangle three buckets)算法将采样[27]挑选第1代核心育种群体,要求挑选出来的群体的数量尽可能少,但最大程度维持遗传多样性和高育种值。每株优树有12个特征值,第1-第11个特征值为11对EST-SSR分子标记引物大小,第12个特征值为单株立木材积育种值。
3 结果与分析
3.1 优树自由授粉子代生长性状表型变异
表2列出了3片优树自由授粉子代测定林20 a林龄时树高、胸径和单株立木材积的均值、年均生长量以及家系间的表型变异系数。20 a林龄时平均树高、胸径和单株立木材积分别为10.96 m、16.93 cm和0.129 00 m3,年均生长量分别为0.55 m、0.84 cm和0.006 45 m3,生长迅速。树高、胸径和单株立木材积在家系间的表型变异系数分别为5.87%、8.60%和14.89%,家系间生长性状表型变异丰富,且单株立木材积的表型变异较树高和胸径丰富。
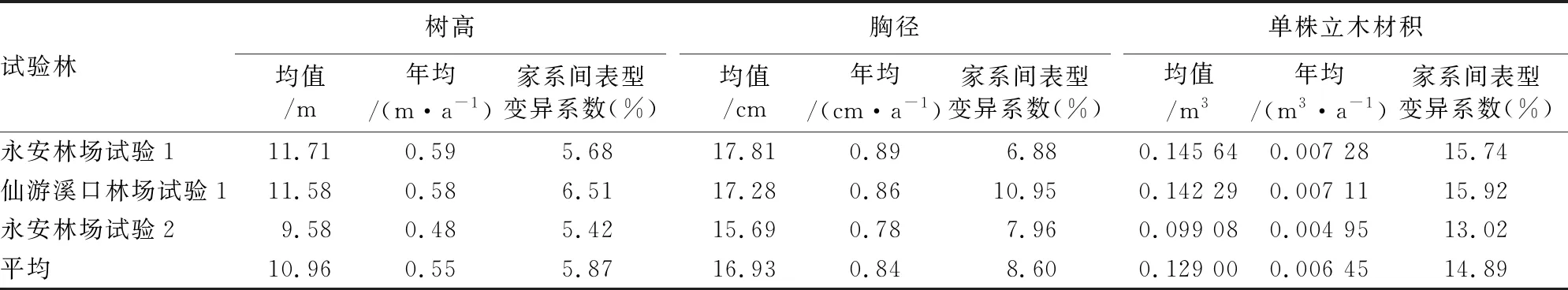
表2 优树自由授粉子代测定林平均生长
3.2 优树自由授粉子代生长性状遗传变异
表3列出了试验林20 a林龄时生长性状的方差分析和遗传参数估算结果,遗传参数汇总成表4。树高、胸径和单株立木材积在家系间差异极显著,狭义遗传力均值介于0.214 7~0.308 5,受到中度偏下的遗传控制。就树高而言,就树高而言,家系间遗传变异系数均值2.77%,家系遗传变异占群体均值的2.77%,家系间遗传分化系数均值19.75%,也即遗传变异的19.75%源自家系间,狭义遗传力与广义遗传力比值均值为0.587 5,加性和非加性遗传效应对树高起着较为均衡的遗传控制作用。就胸径而言,家系间遗传变异系数均值4.57%,家系遗传变异占群体胸径均值的4.57%,家系间遗传分化系数均值32.38%,也即遗传变异的32.38%源自家系间,狭义遗传力与广义遗传力比值均值为0.472 7,加性和非加性遗传效应对胸径也起着较为均衡的遗传控制作用。就单株立木材积而言,家系间遗传变异系数均值7.86%,家系遗传变异占群体单株立木材积均值的7.86%,家系间遗传分化系数均值28.56%,也即遗传变异的28.56%源自家系间,狭义遗传力与广义遗传力比值均值为0.464 1,加性和非加性遗传效应同样对单株立木材积起着较为均衡的遗传控制作用。
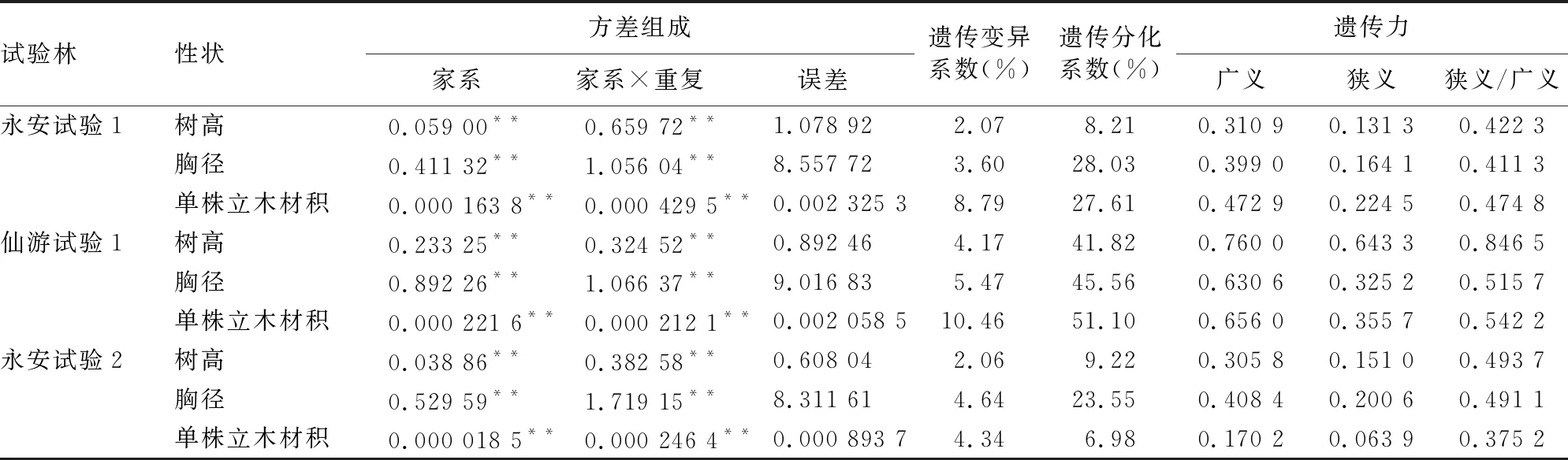
表3 生长性状方差分析及遗传参数

表4 生长性状遗传参数均值
3.3 第1代育种群体筛选
由于树高、胸径和单株立木材积均受到加性和非加性遗传效应共同的、较为均衡的遗传控制,使后向选择优树能获得稳定的生长遗传增益。相比于树高和胸径,单株立木材积家系遗传变异系数与狭义遗传力的乘积更大,对单株立木材积进行选择可以获得更为有效的遗传增益。因此,以单株立木材积育种值为指标对优树后向评价,并依据育种值排名淘汰单株立木材积育种值低于群体平均育种值(0)减去1个标准差的优树,选出111株优树为福建柏第1代育种群体。树高、胸径和单株立木材积平均育种值分别为0.03 m、0.05 cm和0.000 89 m3,观察杂合度(Ho)为0.359 2,Shannon信息指数(I)为1.305 3,等位基因数(Na)为6.181 8,有效等位基因数(Ne)为3.279 2(表5)。
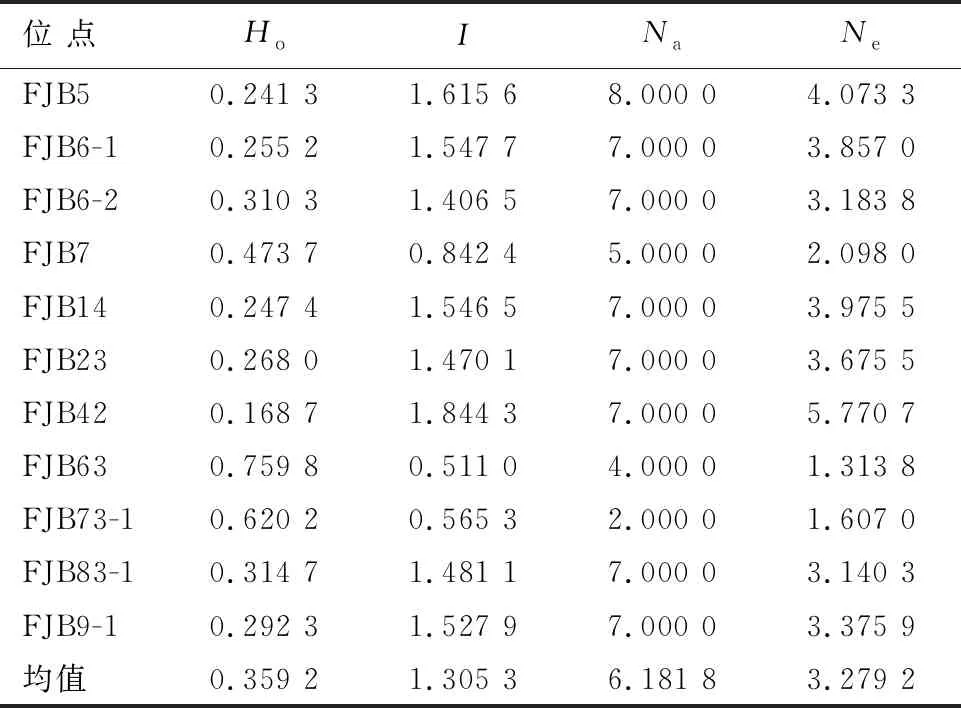
表5 第1代育种群体遗传多样性分析
3.4 第1代核心育种群体构建
对第1代育种群体的111株优树抽样,经迭代程序初步确定抽样数40左右组成的最佳群体具有较高的遗传多样性和单株立木材积育种值(图1A)。根据程序输出的不同抽样数量的最佳组合结果,绘制抽样数40前后10%的抽样数组合的遗传多样性占总多样性的比例曲线,以及育种值与总育种值的比值曲线(图1B)。由图1可以发现,抽样数为41时挑选的最佳组合,其遗传多样性和单株立木材积育种值曲线相交于最高点,也即同时具有较丰富遗传多样性和较高材积育种值。输出的41个最优家系组合为第1代核心育种群体(表6),保留了第1代育种群体73.58%的遗传多样性,单株立木材积育种值是第1代育种群体的1.63倍(图1B)。核心育种群体子代20 a林龄时单株立木材积、育种值、遗传增益平均值分别为0.132 41、0.002 75 m3和1.87%。第1代核心育种群体的遗传多样性参数见表7,观察杂合度(Ho)为0.359 8,Shannon信息指数(I)为1.322 8,等位基因数(Na)6.545 5,有效等位基因数(Ne)为3.306 9。

表6 福建柏第1代核心育种群体

表7 第1代核心育种群体遗传多样性分析
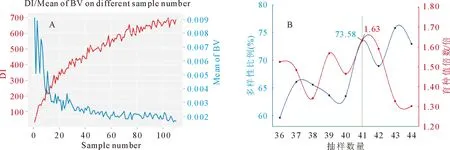
图1 核心育种群体迭代筛选
4 结论与讨论
4.1 结论
福建柏优树自由授粉子代20 a林龄时平均树高、胸径和单株立木材积分别为10.96 m、16.93 cm和0.129 00 m3,年均生长量分别为0.55 m、0.84 cm和0.006 45 m3。树高、胸径和单株立木材积具有丰富的遗传变异,狭义遗传力介于0.214 7~0.308 5,受到中度偏下的遗传控制,分别有19.75%、32.38%和28.56%的遗传变异源自家系间。同时,树高、胸径和单株立木材积狭义遗传力与广义遗传力比值介于0.464 1~0.587 5,表明加性和非加性遗传效应均较为均衡地控制着优树的生长性状,使后向选择优树能获得稳定的生长遗传增益,奠定了从优树中筛选福建柏第1代育种群体的基础。相比于树高和胸径,单株立木材积遗传变异系数与狭义遗传力的乘积最大,具有最大的选择增益,提供了以单株立木材积育种值后向评价优树的可行性。因此,本研究依据育种值排名筛选出111株优树为福建柏第1代育种群体,树高、胸径和单株立木材积育种值分别为0.03 m、0.05 cm和0.000 89 m3,观察杂合度(Ho)为0.359 2,Shannon信息指数(I)为1.305 3,等位基因数(Na)为6.181 8,有效等位基因数(Ne)为3.279 2。
经迭代抽样筛选出41株具有最高的遗传多样性和单株立木材积育种值的优树组合构成的第1代核心育种群体,观察杂合度(Ho)为0.359 8,Shannon信息指数(I)为1.322 8,等位基因数(Na)6.545 5,有效等位基因数(Ne)为3.306 9。保留了第1代育种群体73.58%的遗传多样性,单株立木材积育种值是第1代育种群体的1.63倍。其自由授粉子代20 a林龄时单株立木材积均值、育种值和遗传增益分别为0.132 41、0.002 75 m3和1.87%。
4.2 讨论
在林木遗传测定中,基因的显性作用和上位作用等非加性遗传效应往往对数量性状具有影响。对于育种材料选择来说,却更关注加性遗传效应对性状的作用,其是可稳定遗传的效应。由此,狭义遗传力与广义遗传力的比值在选择决策中就特别重要。其理论值介于0~1,越接近0,表明性状主要受非加性遗传效应控制,此时的选择重点应在无性繁殖材料筛选上;如果其值接近1,表明加性遗传效应对性状起着主要的控制作用,此时筛选育种材料才能获得稳定的遗传增益。本研究中加性和非加性遗传效应对福建柏优树的树高、胸径和单株立木材积生长均起着较为均衡的控制作用,是开展第1代育种材料选择的基础。
林木育种群体与林木核心种质的最大区别在于其主要强调了育种目标[17],或者说林木育种群体是来源于某一树种种质资源中,在某个或多个目标性状中具有优良表现和遗传多样性的种质集。虽然林木育种群体和核心种质在包含种质数量上并没有明确的比较关系,但从育种实践来看,育种群体包含的种质数量一般比核心种质多。构建林木育种群体的主要目的是推动种质创新及品种选育,构建林木核心种质的主要目的在于物种遗传多样性保护。而关于育种群体的容量,300~400是许多森林遗传学家提出的林木育种群体大小的准则和建议,但根据树种不同,有时育种群体的大小不一定会符合这个准则。如加拿大在美洲山杨的育种中,曾经建立的育种群体大小为150份种质[25]。有效大小为20~40的育种群体可以有效支撑几个选育世代的高遗传增益,特别是对于具有亚结构的育种群体,核心育种群体大小和亚系大小设置在20~40是适宜的。因此,育种群体及核心育种群体的容量因树种的育种策略、资源基础不同而异。
虽然福建柏种内具有丰富的遗传多样性[28],但由于过度砍伐等原因,其天然植株在自然界丰度较小,属于我国二级保护植物[1,6]。对其开展育种研究及应用,扩大人工林面积,对保护其遗传多样性及发挥其木材经济效益均有关键作用。但由于天然遗传资源匮乏,构建其第1代育种群体时不可避免地会面临种质资源缺乏的问题。本研究中收集的优树仅为127株,构建的第1代育种群体容量仅为111份,正是缘于此。但却构建了福建柏长期育种的遗传多样性和遗传增益的源泉。基于生长性状育种值和群体遗传多样性构建的41份核心育种群体保留了第1代育种群体73.58%遗传多样性,使单株立木材积育种值大幅提高,虽然丧失了部分遗传多样性,但却获得了较高的生长遗传增益,更是奠定了高效利用福建柏种质资源开展种质交配创新的基础。
