水肥耦合对蓝靛果忍冬幼苗细根生长及根抗氧化酶的影响
2022-11-29刘洋张启昌张璐李玉灵
刘洋,张启昌,张璐,李玉灵
(1.河北农业大学林学院,河北 保定 071001;2.吉林农业科技学院农学院,吉林 吉林 132101;3.北华大学林学院,吉林 吉林 132013)
蓝靛果忍冬(Lonicera caerulea)是兼有食用价值、经济价值、医用价值的浆果类落叶灌木,果实含有氨基酸、黄酮、维生素、酚醛类物质等,具有抗氧化、清热解毒的功效,已经成为新一代小浆果树种,深受人们的喜爱[1-4]。
细根可以反映植物对土壤水分和养分的适应状况[5],对整株植物的生长具有重要影响。良好的根系发育可以缩短缓苗时间,使植株提早进入快速生长期[6]。在土壤水肥极度缺少或过量时,根系生理特性也会出现相应变化以维持植物正常生长[7-8]。植物处在逆境环境下,会产生自由基等有害物质,自由基会破坏细胞质膜,使相对电导率升高,这时植物体内保护酶系统开始启动,清除过剩自由基等有害物质从而维持植物正常生长。研究植物体内抗氧化酶活性可以判断植物抵抗逆境的能力[9-10]。林兴军[11]对番茄体内抗氧化酶的研究发现,干旱胁迫会引起超氧化物歧化酶(soperoxide dismutase,SOD)、过 氧 化 物 酶(peroxidase,POD)、过氧化氢酶(catalase,CAT)、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)等抗氧化酶活性升高,施肥对番茄叶片SOD和CAT均产生显著影响,干旱胁迫下增加施肥量可以使酶的活性降低。根系直接吸收水分和养分,受水肥的影响最敏感,当受外界环境胁迫时,细胞内大量自由基的累积导致细胞的膜脂过氧化,根系会做出积极响应,应对环境的变化[12]。张希吏等[13]研究了干旱胁迫对沙芥幼苗根系抗氧化酶活性的影响,发现POD活性、CAT活性、抗坏血酸活性会随着胁迫的加剧而升高,SOD活性则出现了先上升后下降的变化趋势,说明幼苗根系在干旱胁迫下受到了一定程度的伤害。里程辉等[14]研究淹水胁迫下不同中间砧对苹果岳冠根系抗氧化酶活性的影响,表明根系中SOD、POD、CAT、APX活性均先上升后下降,胁迫初期虽然诱导了酶保护系统,减轻自由基对根系细胞的伤害,但是随着胁迫程度的加剧,超过了植物的承受能力,抗氧化酶活性降低。水肥耦合对植物细根生长和根系抗氧化酶的影响研究多集中在蔬菜和作物栽培中,在小浆果类树种中研究较少。近几年对蓝靛果忍冬生理特性的研究也受到广泛关注,从目前研究来看,对其生理特性影响的研究主要集中在低温、水分、养分、盐分胁迫的研究[15-19]。水肥是影响蓝靛果忍冬幼苗生长的重要环境因子,目前蓝靛果忍冬幼苗根系水肥耦合效应研究较少,对细根生长及根系抗氧化酶的影响研究未见报道。因此,通过对蓝靛果忍冬细根生长和根系抗氧化酶系统对水肥耦合的响应进行研究,认识蓝靛果忍冬细根生长和根系生理特性与土壤环境的作用机理和响应规律,以期为蓝靛果忍冬苗期立地筛选、水肥精准管理提供理论依据。
1 材料与方法
1.1 试验区概况
试验在吉林省吉林市北华大学东校区进行。该地区属中温带大陆性季风气候,四季分明。年无霜期120~130 d,年日照时数2 400~2 600 h,年平均气温4℃,最热月(7月)月平均气温21℃,最冷月(1月)月平均气温1℃。年平均降水量450~670 mm,80%降雨量集中在夏季。
基质为农田土,有机质含量为15.23 g·kg-1,全氮(N)含量2.87 g·kg-1,全磷(P)含量1.08 g·kg-1,全钾(K)含量25.88 g·kg-1,速效N含量132.68 mg·kg-1,速 效P含 量38.69 mg·kg-1,速 效K含 量288.69 mg·kg-1,pH 6.24。测定培养土田间最大持水量(field capacity,FC)为28.29%。
1.2 试验材料
试验材料为蓝靛果忍冬“北华1号”1年生扦插苗,选择根系生长一致的幼苗。试验采用盆栽试验,每盆栽植1株,共20个处理,每个处理8盆,共160株。试验用花盆规格为30 cm(上口内径)×25 cm(高),上盆前用3%硫酸亚铁对土壤进行消毒处理。缓苗期25 d,至4月30日缓苗结束,缓苗期间进行正常养护管理。
1.3 试验设计
试验采用三因素五水平二次旋转回归组合设计,试验因素分别为土壤水分(W)、氮肥施入量(N)和磷肥施入量(P)。各试验因素水平编码见表1。
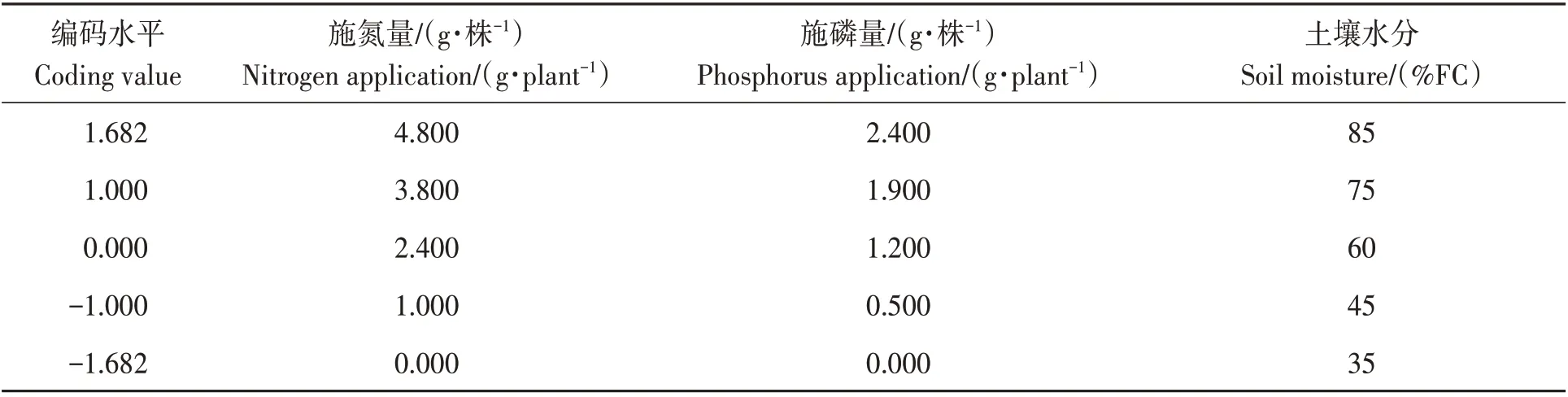
表1 试验因素水平编码Table 1 Level coding of experimental factors
土壤水分:35%FC为重度干旱胁迫,45%FC为中度干旱胁迫,60%FC为轻度干旱胁迫,75%FC为适合水分条件,85%FC为过量灌溉。根据1年生蓝靛果忍冬幼苗收获时含氮量、含磷量结合常规生产施肥量,确定了施氮量和施磷量。施钾量为常规生产施肥量。20种处理实际值见表2。
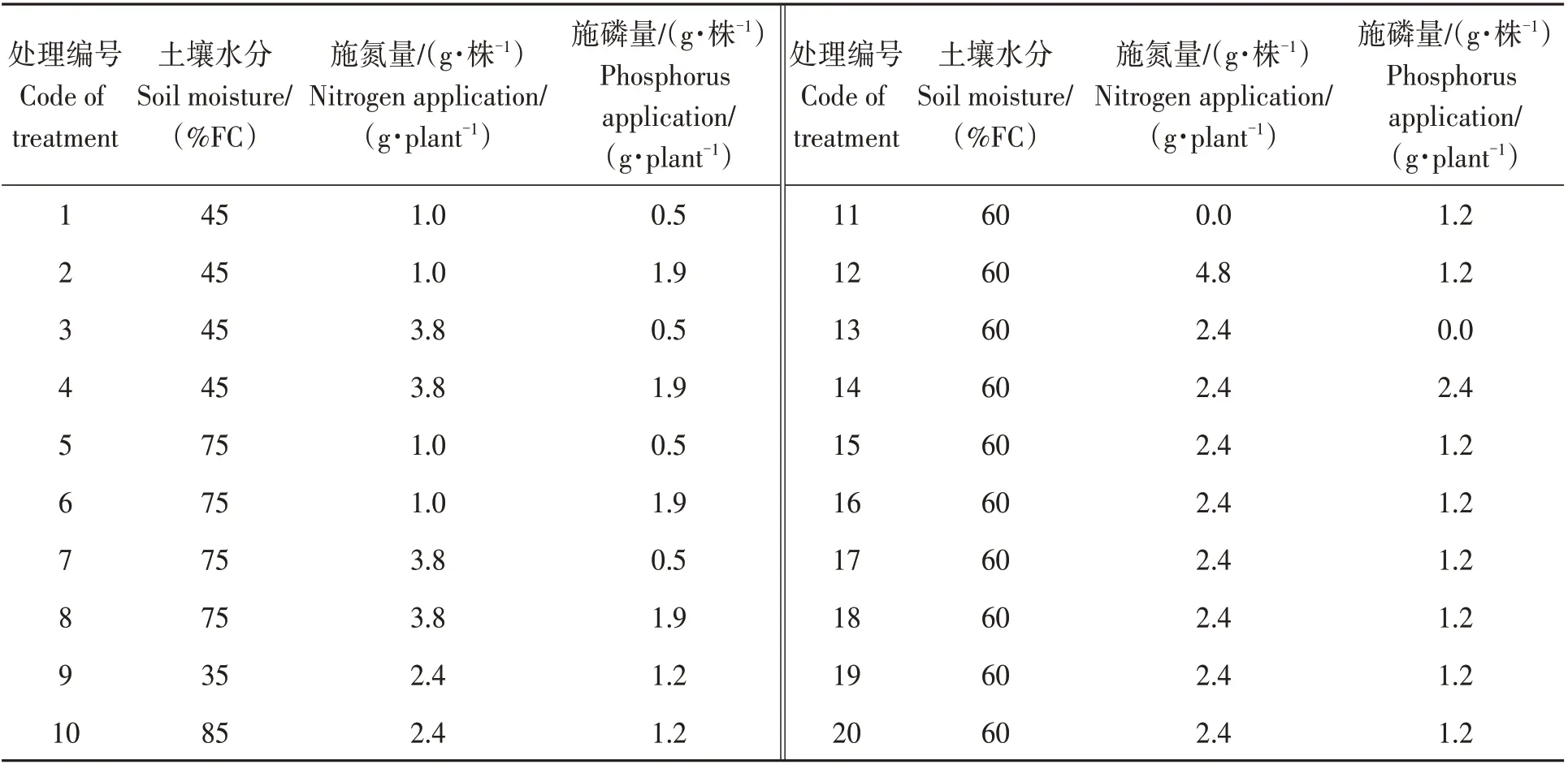
表2 不同处理土壤水分、施氮量、施磷量实际值Table 2 Soil moisture,nitrogen application and phosphorus application under different treatments
1.4 试验因子控制方案
氮肥用尿素(CH4N2O,含N为46%,山西阳煤丰喜肥业),磷肥用分析纯过磷酸钙((CaH2PO4)2·CaSO4·H2O,含P为16%),钾肥用分析纯硫酸钾(K2SO4,含K为50%)。氮肥共分3次施入(5月26日、6月20日、7月15日),磷肥根据计算量于5月26日一次性施入,钾肥(3 g·株-1)定量于5月26日一次性施入。施肥方式:分别在幼苗周围4个方位距根茎5 cm处挖穴埋施,每穴施肥量相等,施肥于浇水当日进行。
土壤水分控制采用整盆称重法结合便携式土壤湿度、温度和盐度计(FOM/mts)进行校订。缓苗期结束后进行控水试验,控水时间为4月30日至9月30日,每2 d于傍晚补水1次。每个处理任意挑选不连续摆放的3盆进行称重,称重时轻拿轻放于2 min内完成,补水采用1 000 mL量筒(精确度10 mL)。根据随机挑选的3盆幼苗灌水量的平均值确定该处理其他苗盆的灌水量。

1.5 测定指标
1.5.1 细根生长指标测定 按照Pregitzer等[20]的方法对根样进行根系分级(图1)。分级后,将各级根用Epson数字化扫描仪扫描,将扫描后的图像存入计算机,用于扫描仪配套的WinRHIZO(Pro2004b)根系图像分析系统软件对各级根系形态进行定量分析。扫描后的各级根系分别装入信封,在105℃烘箱中杀青1 h,然后再置于65~70℃烘箱中烘干至恒重,用天平对其称重,记录的数据即为生物量干质量。
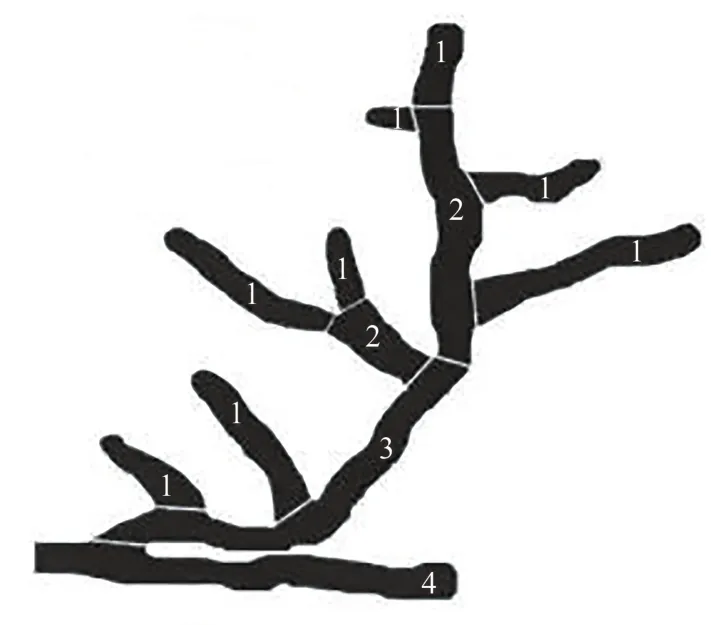
图1 根序分级示意图[20]Fig.1 Schematic diagrams of root identification[20]
根据根系图像分析系统软件可以直接测量和计算细根的根长。比根长按照下列公式计算。

1.5.2 根系抗氧化酶指标测定 依照蔡庆生[21]的测定方法对蓝靛果忍冬幼苗根系SOD、POD、谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)进行测定;参照Nakano等[22]的方法测定APX活性。
1.6 数据处理与分析
采用Microsoft Excel 2019进行数据处理,SPSS 25.0和SAS9.0进行ANOVA方差分析、回归模型建立和回归系数检验,利用Sigmaplot 12.0绘图,利用Lingo18.0进行多目标决策模型分析。本研究中各指标与土壤水分、施氮量、施磷量回归分析采用以下模型。
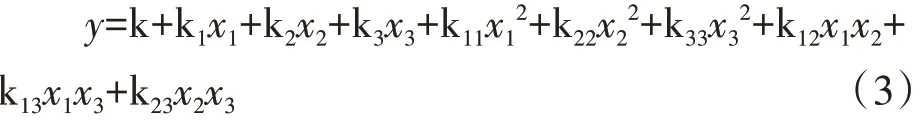
式中,y表示响应变量,x1表示土壤水分(W),x2表示施氮量(N),x3表示施磷量(P),k表示模型的回归系数。试验中,各响应变量的回归系数已标准化,各影响因素的水平已经过无量纲线性编码代换。其系数正负号表示各因素对变量的影响方向,直接比较其系数绝对值可以判断各因素对响应变量的影响程度。
2 结果与分析
2.1 水肥耦合对蓝靛果忍冬幼苗细根根长的影响
蓝靛果忍冬幼苗一共有5级根,根据根序法进行根系分级,把蓝靛果忍冬幼苗的1~3级根定为细根。对蓝靛果忍冬幼苗细根根长进行测定,结果如图2,不同处理1~3级根根长变化范围分别为122.24~338.69、75.99~159.65、28.44~86.21 cm。处理8明显高于其他处理,1级根根长是处理3的2.77倍;2级根根长是处理2的2.11倍,3级根根长是处理4的3.03倍,均达到显著差异(P<0.05),说明适当的水肥处理有利于蓝靛果忍冬细根长度的增加。
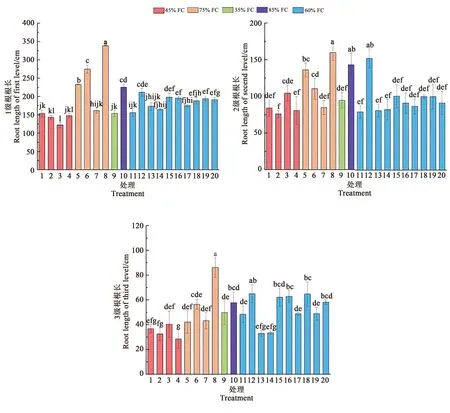
图2 蓝靛果忍冬1~3级根根长Fig.2 Length for the 1~3 order roots in Lonicera caerulea
2.2 水肥耦合对蓝靛果忍冬幼苗细根比根长的影响
如图3所示,不同处理蓝靛果忍冬幼苗1~3级根的比根长变化范围分别是76.93~225.26、21.48~53.22、4.03~16.24 m·g-1,说明不同水肥处理对蓝靛果忍冬幼苗细根的比根长有明显影响。其中,处理8的1级、2级、3级根比根长显著高于其他处理(P<0.05),分别是比根长较低的处理9的2.93、1.79、3.57倍,是处理11的2.39、2.42、3.37倍。说明适当的水肥配施能明显增加蓝靛果忍冬细根的比根长,有利于根对营养物质的吸收。

图3 蓝靛果忍冬1~3级根比根长Fig.3 Specific root length for the 1~3 order roots in Lonicera caerulea
2.3 水肥耦合对蓝靛果忍冬幼苗根系SOD活性的影响
如图4所示,不同处理蓝靛果忍冬幼苗根系SOD活性变化范围在7.62~32.99 U·g-1·min-1,处理4的SOD活性最高且与其他处理差异显著(P<0.05),处理1和处理9的SOD活性也较高。处理7、处理8和处理15~20的SOD活性较低。处理4是处理15~20的3.6~4.3倍,SOD活性在轻度胁迫下较稳定,当水肥胁迫程度加剧时,SOD活性升高,当达到重度胁迫时,SOD活性又降低。说明水肥配施有利于维持蓝靛果忍冬根系SOD活性的稳定,水肥不足会引起抗氧化酶活性升高。

图4 不同处理蓝靛果忍冬根系超氧化物歧化酶活性的变化Fig.4 Changes of SODactivity in the roots of Loniceracaerulea under different treatments
2.4 水肥耦合对蓝靛果忍冬幼苗根系POD活性的影响
如图5所示,不同处理蓝靛果忍冬幼苗根系POD活性变化范围在10.44~39.26 U·g-1·min-1。处理3、4、13、14的POD活性较高;处理5、7、15~20和处理8的POD活性较低。其中,处理13的POD活性是处理15~20的2.6~3.8倍,且达到显著差异(P<0.05),在土壤水分和施氮量充足的条件下以及土壤水肥轻度胁迫下POD活性较稳定,当中度干旱胁迫时,POD活性升高,当磷肥施入不足或过量时也会引起POD活性升高,进一步说明水肥配施有利于维持蓝靛果忍冬根系POD活性的稳定,水肥不足或过量灌溉施肥影响了POD活性的稳定。
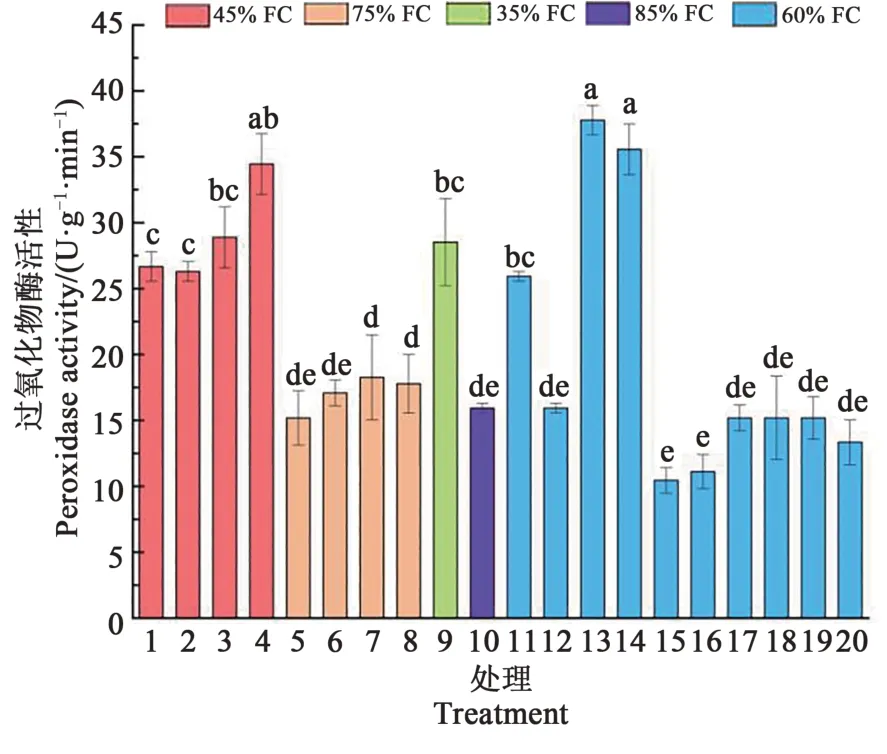
图5 不同处理蓝靛果忍冬根系过氧化物酶活性的变化Fig.5 Changes of PODactivity in the roots of Loniceracaerulea under different treatments
2.5 水肥耦合对蓝靛果忍冬幼苗根系APX活性的影响
如图6所示,不同处理蓝靛果忍冬幼苗根系APX活性变化范围在44.00~194.00 U·g-1·min-1。处理15~20、处理5~8和处理10的APX活性较低;处理2、3、9、11和处理13的APX的活性较高。根系APX的活性对土壤水肥较为敏感,当土壤水肥条件合适时,根系APX活性稳定,当中度干旱胁迫和重度干旱胁迫时均引起APX活性升高,未施磷肥的处理13和未施氮肥的处理11的APX活性也较高。另外,处理4为中度干旱胁迫但氮磷施入适量,虽然APX活性升高,但由于氮磷的合理施入有效缓解了干旱胁迫。

图6 不同处理蓝靛果忍冬根系抗坏血酸过氧化物酶活性的变化Fig.6 Changesof APX activity in the roots of Loniceracaerulea under different treatments
2.6 水肥耦合对蓝靛果忍冬幼苗根系GSH-Px活性的影响
从图7可以看出,不同处理蓝靛果忍冬幼苗根系GSH-Px活性变化范围在1.43~4.09 U·g-1·min-1。处理1的GSH-Px活性最高,且与其他处理达到了显著差异(P<0.05),处理11和处理6的GSH-Px活性也较高;处理10、15~20的GSH-Px活性较低。水肥胁迫影响了GSH-Px活性,适水适肥条件下蓝靛果忍冬幼苗根系GSH-Px活性稳定。处理1与3达到显著差异(P<0.05),二者只是施氮量不同,处理6和处理8达到显著差异(P<0.05),二者也是施氮量不同,说明施氮量对GSH-Px活性有显著影响。
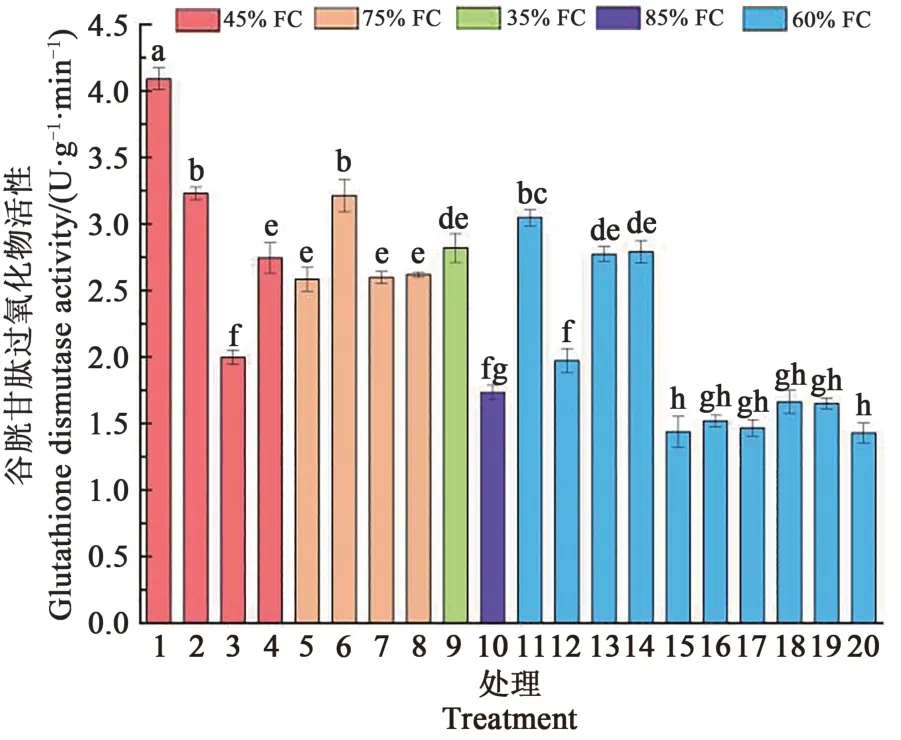
图7 不同处理蓝靛果忍冬根系谷胱甘肽过氧化物酶活性的变化Fig.7 Changes of GSH-Px activity in the roots of Loniceracaerulea under different treatments
2.7 水肥因子效应分析
2.7.1 蓝靛果忍冬幼苗细根根长 分别以蓝靛果忍冬幼苗1~3级根根长y1~y3为响应变量,土壤水分(x1)、施氮量(x2)、施磷量(x3)为自变量,对回归模型进行检验分析,3个回归模型均达到了显著性水平(P<0.05),说明模型与实际情况拟合较好,响应变量与土壤的水分、施氮量、施磷量有较强的相关性(R2范围在0.556~0.732)。在剔除不显著因素后,各响应变量与土壤水肥的模型如下。

模型4中,x1和x3的回归系数均为正值,且|x1|>|x3|,说明土壤水分、施磷量正向作用于1级根根长,其中土壤水分是主效应。耦合效应方面,土壤水分和施氮量、施氮量和施磷量对1级根根长有显著耦合正效应。模型5中,x1和x2的回归系数均为正值,且|x1|>|x2|,说明土壤水分和施氮量正向作用于2级根根长,主效应为土壤水分,耦合效应分析,施氮量和施磷量对2级根根长有显著耦合正效应。模型6中,x1和x2回归系数均为正数,且|x1|>|x2|,说明土壤水分为主效应,耦合效应分析施氮量和施磷量对3级根根长有显著耦合正效应。综合分析模型4~6,土壤水分、施氮量、施磷量均对蓝靛果忍冬细根根长有显著影响,其中土壤水分是主要影响因子,耦合效应分析施氮量和施磷量对细根根长耦合效应最显著。
2.7.2 蓝靛果忍冬幼苗细根比根长 分别以蓝靛果忍冬幼苗1~3级根比根长y1~y3为响应变量,土壤水分(x1)、施氮量(x2)、施磷量(x3)为自变量,对回归模型进行检验分析,3个回归模型均达到了显著性水平(P<0.05),说明模型与实际情况拟合较好,响应变量与土壤水分、施氮量、施磷量有较强的相关性(R2范围为0.435~0.726)。在剔除不显著因素后,各响应变量与土壤水肥的回归模型如下。

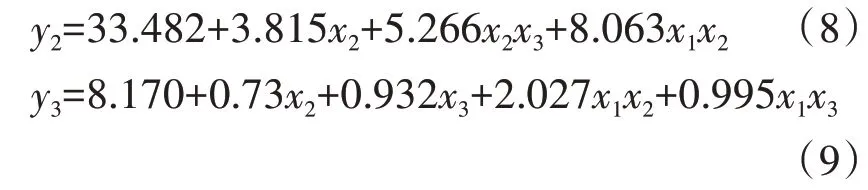
模型7~9中,综合分析x1、x2、x3、x1x2、x1x3、x2x3的回归系数,土壤水分、施氮量、施磷量均正向作用于细根比根长,土壤水分为主效应。耦合效应方面,土壤水分和施氮量耦合效应对细根比根长有显著耦合正效应。
2.7.3 蓝靛果忍冬幼苗根系抗氧化酶 分别以蓝靛果忍冬幼苗根系超氧化物酶活性(ySOD)、过氧化物酶活性(yPOD)、抗坏血酸过氧化物酶活性(yAPX)、谷胱甘肽过氧化物酶活性(yGSH-Px)为响应变量,土壤水分(x1)、施氮量(x2)、施磷量(x3)为自变量,对回归模型进行检验分析,4个回归模型均达到了显著性水平(P<0.01),说明模型与实际情况拟合较好,响应变量与土壤的水分、施氮量、施磷量有较强的相关性(R2范围为0.712~0.871)。在剔除不显著因素后,各响应变量与土壤水肥的回归模型如下。

根据回归模型10~13,总结了土壤水肥因子对根系抗氧化酶活性的影响(表3)。各测定指标对水肥耦合的响应略有差别,土壤水分、施磷量对蓝靛果忍冬幼苗根系抗氧化酶活性均有显著负效应,其中施磷量对POD影响最显著,施氮量对SOD、APX、GSH-Px影响最显著。耦合效应方面,施氮量和施磷量耦合效应对所测定的抗氧化酶活性影响最显著。

表3 蓝靛果忍冬幼苗根系抗氧化酶活性土壤水肥因子效应综合分析Table 3 Effects analysis of soil and fertilizer application on antioxidant enzyme activity of root of Lonicera caerulea seedlings
2.8 对模型进行寻优
通过模型优化计算及综合分析结果得出,蓝靛果忍冬幼苗细根生长和维持根系抗氧化酶活性稳定的最优水肥组合为:土壤田间最大持水量的69%,施氮量为3.3 g·株-1,施磷量为1.2 g·株-1,在此处理下蓝靛果忍冬幼苗细根总根长为521.46 cm、细根总比根长为300.51 m·g-1;蓝靛果忍冬根系SOD活性、POD活性、APX活性、GSH-Px活 性 分 别 为13.84、15.41、67.56、1.73 U·g-1·min-1。
3 讨论
蓝靛果忍冬幼苗细根各生长指标对土壤水肥因子的响应规律相似,合理的水肥施入量及其耦合效应有利于促进蓝靛果忍冬幼苗细根生长。其中,处理8幼苗细根生长最好,这与程奇云[23]的研究施肥会显著影响果树根系生长且有助于苗木生长的研究结论一致。本研究表明,在适水适氮适磷处理下,蓝靛果忍冬幼苗细根根长和比根长明显高于低水处理、无氮处理和无磷处理。蓝靛果忍冬幼苗细根在生长期对土壤水分敏感,其水氮耦合、水磷耦合、氮磷耦合对细根根长和比根长有显著正效应,氮磷耦合对细根根长影响最显著,而水氮耦合对细根比根长影响最显著。
植物体内抗氧化酶的含量变化是表征植物逆境下应对胁迫的重要指标。植物处在胁迫条件下,植物体内活性氧增多会产生伤害植物生长的氧化产物,这些氧化产物会破坏植物细胞质膜完整性,导致细胞损伤,从而影响植物生长[24-26]。为保护植物正常生长,在植物遭受胁迫时,植物体内会产生一系列的抗氧化酶来清除自由基[27]。本研究表明,水肥适合的处理7、处理8和轻度胁迫处理15~20蓝靛果忍冬幼苗根系抗氧化酶活性较低,处理1~4(中度干旱胁迫)、处理9(重度干旱胁迫)、处理11(无氮)和处理13(无磷)蓝靛果忍冬根系中抗氧化酶活性均升高,这与邹养军[28]的研究苹果长时间处在缺水条件下根系SOD、POD的活性会明显增高的结果基本一致,与侯栋等[29]以2年生香玲等4个核桃品种为试材不同干旱胁迫处理对其POD活性的影响结果也基本一致。在主效应和耦合效应分析中,施氮量为主效应,施磷量和施氮量耦合效应对幼苗根系抗氧化酶影响最为显著。以上表明合理施肥更有利于维持抗氧化酶活性的稳定性,过度灌溉、施肥或水肥缺失下,植物体内产生大量自由基使其正常代谢受到干扰,细胞处于逆境时植物通过升高抗氧化酶活性以清除这些氧化物,使细胞质膜所受伤害降低,植物体内过氧化酶升高说明细胞环境稳定性受到了破坏。需要注意的是,处理15~20抗氧化酶活性较低,轻度胁迫下抗氧化酶活性并未升高,说明在轻度逆境下蓝靛果忍冬幼苗根系抗氧化酶系统未启动,蓝靛果忍冬对于逆境有一定的抵抗能力,但随着水分和养分胁迫的加剧,抗氧化酶系统启动,抗氧化酶活性升高,而当超过了阈值时,如处理9和处理11,抗氧化酶活性又降低,说明抗氧化酶系统遭到了严重破坏,严重影响了植物的正常生长。另外,在研究中还发现不同的酶在不同水肥条件下活性有差别,GSH-Px活性在施氮量不足时活性最高、POD活性在施磷量不足时活性最高,说明不同水肥条件下不同的酶的主导地位不一样,此消彼长、协同作用于植物体以维持细胞正常的代谢平衡保护质膜结构不被破坏。
