新疆三种独尾草的适宜分布及其对未来气候变化的响应
2022-11-26董苏君马松梅何大俊
董苏君,马松梅,张 丹,何大俊,张 林,闫 涵
1 石河子大学生命科学学院, 绿洲城镇与山盆系统生态兵团重点实验室, 石河子 832000
2 石河子大学理学院, 干旱区景观生态重点实验室, 绿洲城镇与山盆系统生态兵团重点实验室, 石河子 832000
物种地理分布格局(空间分布特征)的变化是对气候变化最清楚、最直接的反映[1]。预测气候变化对植物多样性的影响一直是植物地理学和植物生态学研究的热点和重点之一。气候变化对分布范围较窄的植物来说更具有抑制性和选择性,因为这类植物对环境气候的特定性相对较高[2]。大尺度气候变化会对植被分布造成一定影响,局部环境的差异也影响植物产生局部环境适应,造成很多植物类群分布特征相似但环境生态位产生一定分化[3]。相关研究利用遗传变异分析和生态位分析表明温度变化和较低冬季均温以及高度特异性的生境可能是造成昌都韭(AlliumchangduenseJ.M.Xu.)与梭沙韭(AlliumforrestiiDiels)分化的重要原因[4];温度和降水的显著差异造成不同倍性的猕猴桃(中华猕猴桃(Actinidiachinensisvar.chinensis)、美味猕猴桃(Actinidiachinensisvar.deliciosa))、3种云杉(沙地云杉(Piceamongolica)、红皮云杉(Piceakoraiensis)、白扦(Piceameyeri))产生明显的生态位差异[5—6]。新疆独特的山脉-盆地-绿洲-沙漠景观,孕育了丰富的中国新疆仅分布属,如角果藜属(CeratocarpusLinn.)、新疆藜属(AelleniaUlbr)、盐蓬属(Halimocnemis)等,以及中国新疆仅分布种,如沙生柽柳(Tamarixtaklamakanensis)、天山云杉(Piceaschrenkianavar.tianschanica)等,以及国内仅分布于新疆的类短命植物阿尔泰独尾草(Eremurusaltaicus)、粗柄独尾草(Eremurusinderiensis)和异翅独尾草(Eremurusanisopterus)。
独尾草属(EremurusM.Bieb.)隶属百合科(Liliaceae),在我国共有4种,阿尔泰独尾草(Eremurusaltaicus(Pall.) Stev)、粗柄独尾草(Eremurusinderiensis(M.Bieb) Regel)、异翅独尾草(Eremurusanisopterus(Kar.et Kir) Regel)和独尾草(EremuruschinensisFedtsch),前3种仅分布于新疆[7]。其中,阿尔泰独尾草为典型山地植物,主要分布于伊犁河谷地带和准噶尔盆地西北缘,多生长于山地草原和草甸草原;粗柄独尾草和异翅独尾草为典型的沙生植物,均分布于准噶尔盆地西南部,常见于平原固定沙丘、半固定沙丘和沙地[8]。新疆3种独尾草均为早春多年生类短命植物,利用早春季相对充足的水分在2个月左右迅速完成生活史,随后地上部分枯萎、而地下部分进入休眠期以躲避炎热夏季[9]。目前,对于3种独尾草已开展的研究主要体现在解剖学[10]、繁育生物学[11]、化学成分分析[12]等方面。但是,3种独尾草在新疆的生态适宜性如何?适宜的分布范围与空间格局如何?哪些环境因子主要影响了3种植物的适宜分布?以及3种植物对未来气候变化的可能响应,这一系列科学问题都并不清楚,成为限制新疆独尾草资源利用和科学保护的瓶颈。
物种分布模型(SDMs)是评估气候变化对物种分布影响的主要方法,通常用于物种的潜在地理分布研究。常见的物种分布模型主要有最大熵模型(MaxEnt)、广义加性模型、广义线性模型、分类回归树等[13]。其中,最大熵模型基于熵最大原理,利用不完整的物种分布信息,反映物种潜在分布区与环境因子的关系,并构建其地理尺度上的空间分布模型[14]。由于机理清晰、建模简单、模拟结果连续、预测效果较好等特点,成为预测物种潜在地理分布的常用模型之一[15],已被广泛运用于典型荒漠植物的历史分布格局模拟[16]、外来物种的入侵风险模拟[17]、种下谱系分组模拟[18]等研究领域。因此,本研究基于3种独尾草的地理分布信息和环境数据,利用MaxEnt模型结合地理信息系统ArcGIS模拟不同时段3种植物在新疆的适宜分布,利用R程序包、统计学和空间统计学方法分析不同气候因子对3种独尾草适宜分布的影响。旨在解决以下科学目标:(1)基准气候情景下,3种独尾草在新疆的适宜分布范围与空间分布特征;(2)限制独尾草分布的关键环境驱动因子及其适宜的数值范围;(3)3种独尾草的适宜分布对未来气候变化的可能响应。
1 研究数据与研究方法
1.1 分布数据和环境数据的来源与处理
本研究通过检索中国数字植物标本馆(CVH;http://www.cvh.org.cn/)、全球生物多样性信息网络(https://www.gbif.org/)结合已发表文献,共获得114个独尾草属的自然分布点,对有经纬度记载的数据直接使用,对仅有位置记录的分布点则通过BIGMAP地图下载器和中国地名录确定详细地理坐标,并基于R中的raster和biomod2程序包进一步剔除重复和模糊的分布点,矫正分布点偏差。最终,获得64条准确有效的分布记录(阿尔泰独尾草29条、粗柄独尾草18条、异翅独尾草17条)用于模型的模拟,覆盖了3种独尾草属植物在新疆的自然分布范围(图1)。

图1 MaxEnt模型预测的3种独尾草基准、未来气候情景下的适宜分布
本研究中,19个气候变量来源于IPCC生物气候数据集(http://www.worldclim.org),选取2001—2040年、2041—2080年SSP2- 4.5共享社会经济路径联合国政府间气候变化专门委员会(SSPs)下CMIP6模型的气候数据。共享社会经济路径是IPCC描述未来不同气候政策不同社会发展状况下的排放情景。其中包括SSP1—SSP5共5种不同路径。 本研究选择代表温室气体排放中等路径SSP2-4.5。坡度、坡向数据利用ArcGIS10.5空间分析工具从中国科学院地理空间数据云平台(http://www.gscloud.cn/)提供的数字高程模型(DEM)中提取。模型模拟使用的中国省级行政区划底图来源于国家基础地理信息系统网站(http://www.ngcc.cn/)。考虑空间环境变量对物种的限制作用,降低变量的空间纬度避免过拟合现象,运用R软件的Hmisc程序包对22个环境因子进行Pearson相关性分析,剔除相关性高(>80%)的变量,最终筛选获得8个环境变量(表1)。
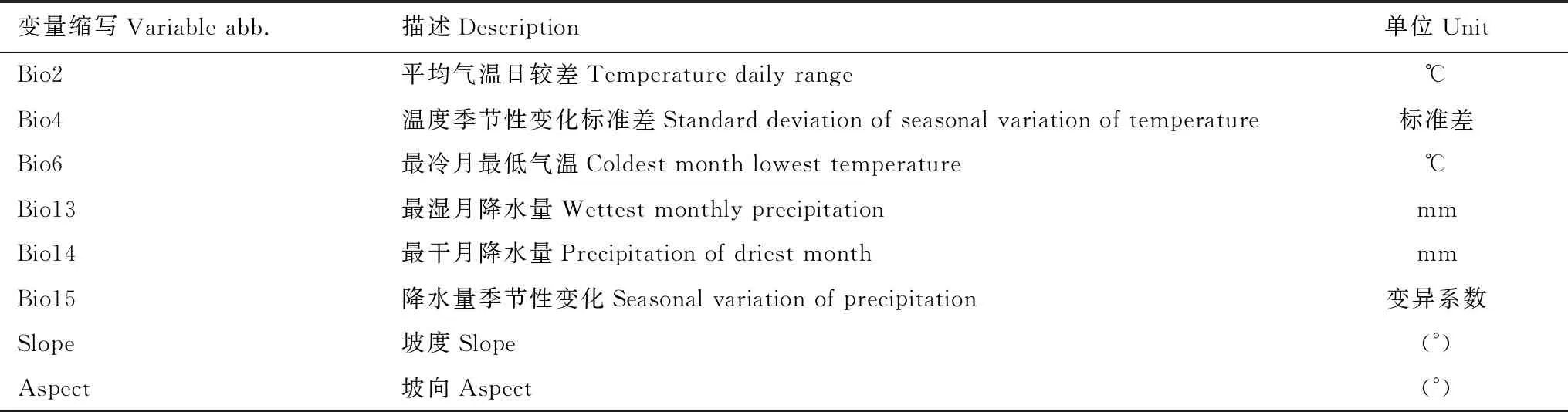
表1 8个环境变量及其相关信息
1.2 最大熵模型构建
利用MaxEnt模型构建,随机选择75%的分布数据作为训练集,剩余25%数据作为测试集,用于检验模型的准确性,重复运算10次。采用受试者工作特征曲线(ROC)下方面积(AUC)值对模型模拟的准确性进行评价[19]。AUC的取值范围为0—1,值为0.8—0.9,表示模型模拟精度较高,为0.9—1.0,表示模型模拟精度极高[20]。为了方便模型模拟结果的可视化表达,参照已发表文献[21],利用ArcGIS空间分析工具计算所有已知分布点在潜在分布图上的适宜性,并计算这些值的标准差σ和平均值μ,选择p=σ-μ作为阈值,区分“适生区”和“不适生区”;选择p=σ+μ作为阈值,区分“适生区”和“高度适生区”。
1.3 多元环境相似度面和最不相似变量分析
利用多元环境相似度面和最不相似变量分析基准时段与未来时段研究区的气候变化及其主要变化因子。利用MaxEnt.jar文件中的density.tools.Novel工具分别计算2001—2040年和2041—2080年研究区内6个气候因子(2个地形因子变化太小而忽略不计)的相似度(S)得到多元环境相似度面。当S值为正值时,S值越小表示该区域气候因子的差异越大,S值为100.00时,表示无差异;当S值为负值时,表示该区域至少有1个气候因子的不相似度超出了参考范围,该区域的环境差异极大[22]。同时,提取相似度最低的气候因子,得到最不相似变量,利用ArcGIS 10.5进行可视化处理。利用MaxEnt模型生成的环境变量贡献率和置换重要值分析各环境因子对模型模拟的贡献以及模型对各环境因子的依赖[23]。
1.4 8个环境因子的主成分分析
为了进一步评价3种植物分布区的环境变量差异以及造成环境差异的主要驱动因子,本研究基于3种独尾草的64个自然分布点和筛选后的8个环境变量进行主成分(PCA)分析,PCA分析利用R4.0.2的ggplot2包完成。
2 研究结果与分析
2.1 3种独尾草的适宜分布及其对未来气候变化的响应
基准及未来气候情境下,3种独尾草模拟的MaxEnt模型的平均AUC值均大于0.935,表明模型模拟结果较准确。沙生植物粗柄独尾草与异翅独尾草适宜分布范围有较大部分的重叠,阿尔泰独尾草与两种沙生独尾草的分布区具有明显差异(图1)。基准气候下,阿尔泰独尾草的适宜分布占研究区总面积的19.46%,集中于北疆伊犁河谷、天山北麓、阿尔泰山中段和准噶尔西部山地(图1)。其中,高度适生区为小面积的破碎斑块,集中于准噶尔西部山地、伊犁河谷,占研究区总面积的2.97%。沙生植物粗柄独尾草与异翅独尾草的适宜分布主要集中于天山北坡古尔班通古特沙漠西南部沙地,适生区面积占比分别为3.06%、1.16%(表2)。具体说,粗柄独尾草的适生区,西起准噶尔盆地西端婆罗科努山的山前沙丘,东至天山中段的古尔班通古特沙漠南缘,其中高度适生区主要呈带状分布于天山北坡中段的绿洲-荒漠过渡带,占研究区总面积的0.60%。异翅独尾草的适生区较小且破碎程度更高,高度适生区呈块状分布于婆罗科努山中段的沙漠南缘,占研究区总面积的0.19%(图1,表2)。
2001—2040年和2041—2080年气候情景下,3种独尾草植物的适生范围整体均呈缩减趋势(图1)。2001—2040年,阿尔泰独尾草以山地适宜分布区边缘及沙漠边缘斑块缩减或消失为主,适宜分布区占比将缩减9.925%,分布整体呈向天山北麓和准噶尔西部山地缩减趋势;2041—2080年的适宜分布将进一步破碎化。粗柄独尾草的适生区在2001—2040年将缩减20.984%,主要表现为适宜分布区内部及边缘破碎化;2041—2080年适生区将向古尔班通古特沙漠南缘显著缩减58.112%。异翅独尾草的适生区2001—2040年将缩减71.607%,整体向沙漠西端婆罗科努山的山前沙丘北坡缩减,且高度适生区基本消失;2041—2080年适生面积将略微增加(图1,表2)。
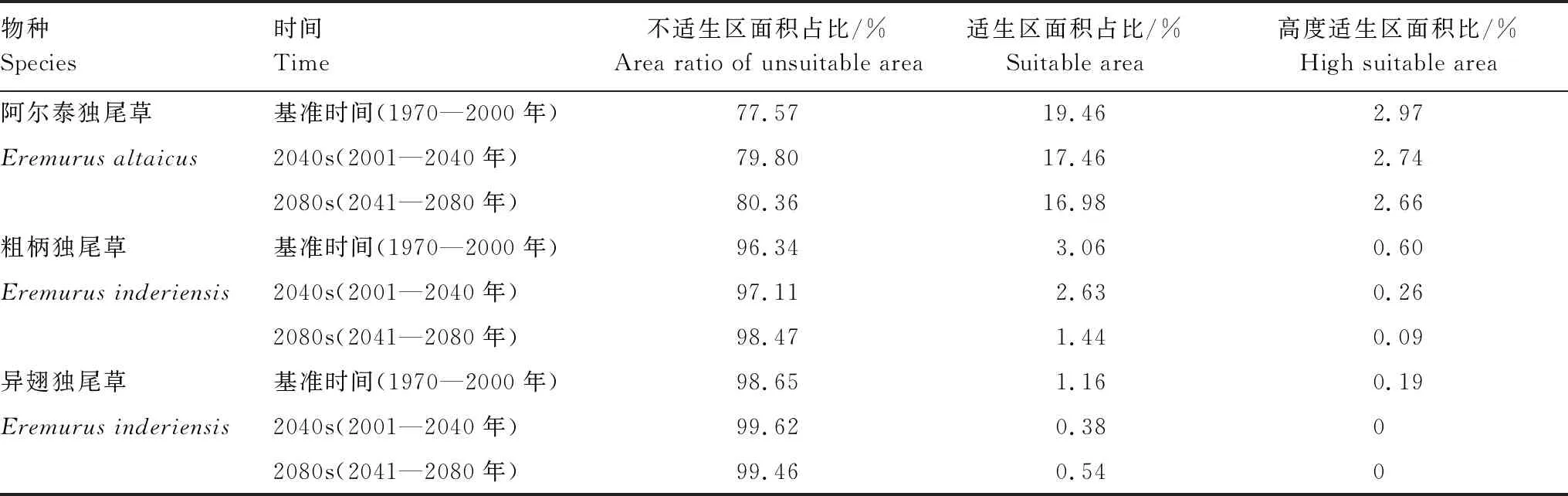
表2 基准、未来气候情景下3种独尾草不同等级适生区面积的变化
2.2 影响3种独尾草适宜分布的环境因子分析
(1)MaxEnt模型模拟的驱动因子及其累积贡献率
最大熵模拟的环境因子分析表明,极端降水因子主要影响了阿尔泰独尾草的适宜分布,最干月降水量和最湿月降水量的累积贡献率达74.8%,另外坡向也对该山地植物的分布具有一定影响,贡献率为10.9%。粗柄独尾草的生态适宜性主要受温度季节性变动指数和最干月降水量的影响,异翅独尾草主要受温度季节性变动指数和坡度的影响,累积贡献率分别为84.7%、83.5%(表3)。
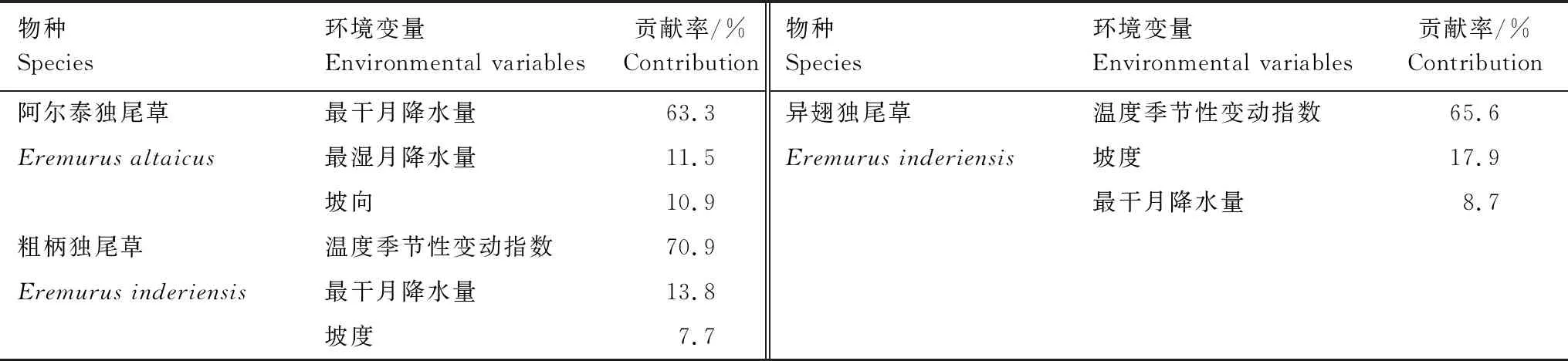
表3 影响3种独尾草分布的主要环境变量及其贡献率
(2)研究区未来气候相对基准气候的波动及其主导因子
多元环境相似度面预测结果表明,2001—2040年和2041—2080年,研究区大部分区域将发生明显的气候波动(图2)。2001—2040年,研究区内气候因子相似度低于20的区域占比为84.03%,相比于基准气候,伊犁河谷、准噶尔盆地南部、塔里木盆地西南部和吐鲁番盆地等均表现出强烈的气候波动(图2)。阿尔泰独尾草、粗柄独尾草和异翅独尾草在基准气候下的绝大部分适宜分布区均将发生明显的气候波动,适生区内气候因子的平均相似度分别为10.00、1.23、0.77(图2)。2041—2080年研究区的气候波动与2001—2040年较相似,但波动范围和程度略微增加,气候因子相似度低于20的区域占比达87.41%,3种植物适生区内气候因子的平均相似度分别为2.87、5.75、6.00(图2)。最不相似变量显示,2001—2040年最冷月最低温(Bio6)、降水量季节性变化(Bio15)、平均日较差(Bio2)、温度季节性变动指数(Bio4)均将发生显著变化,是造成研究区内81.91%的区域发生气候波动的主要因子(图2)。最干月降水量、降水量的季节性变化和最湿月降水量,也是影响阿尔泰独尾草62.07%的分布点发生气候变化的主导因子。温度季节性变动指数是影响粗柄独尾83.33%和异翅独尾草88.24%的分布点气候发生变化的主要因子(图2)。

图2 2001—2040年、2041—2080年气候情景下影响3种独尾草分布的6个驱动因子的多元环境相似度面和最不相似变量
2.3 3种独尾草的环境生态位差异及其驱动因子分析
PCA分析结果显示,基准气候下,3种独尾草植物各适生区内8个环境驱动因子的数值范围存在明显差异,尤其是山地独尾草和沙生独尾草显著分化为两组(图3)。最干月降水量(Bio14)、温度季节性变动指数(Bio4)、最湿月降水量(Bio13)和降水量季节性变化(Bio15)主要导致了3种植物环境生态位的显著差异(图3)。
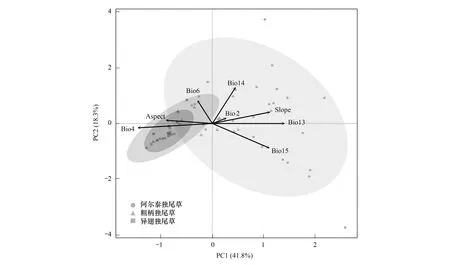
图3 影响3种独尾草分布的8个环境因子的PCA分析
3 结论与讨论
3.1 3种独尾草的适宜分布及其环境驱动机制
基准与未来时段,本研究模型模拟的阿尔泰独尾草和两种沙生独尾草的分布范围均存在明显差异(图1),阿尔泰独尾草的适宜分布集中于天山北麓的山地草甸,两种沙生独尾草主要分布于古尔班通古特沙漠南缘,这与文献记载一致[24]。而且,根据本研究,阿尔泰独尾草与两种沙生独尾草的环境生态位存在较显著的差异(图3)。基准气候下,除在新疆天山和阿尔泰山的适宜分布外[10—11],模型还识别出阿尔泰独尾草在伊犁河谷和准噶尔西部山地具有高度适宜性(图1),相关研究也表明早春类短命植物从最北部的阿尔泰山到最南部的伊犁河谷均有分布[25],这些区域均属新疆重要的水源涵养区,常年植被茂盛,适合山地植物、短命植物等生长[26]。两种沙生植物粗柄独尾草和异翅独尾草的适宜分布有较大范围的重叠,主要集中于准噶尔盆地西南部沙区婆罗科努山的山前沙丘(图1)。准噶尔盆地西南部荒漠拥有丰富的热量资源、相对均匀的降水条件,能够为沙生植物提供较适宜的生长环境,梭梭(H.ammodendron)[27]和白梭梭(H.persicum)[28]等荒漠植物物种均在该区域具有较适宜的分布。两种沙生类短命植物长期适应沙漠恶劣生存环境,进化出了一系列生活史对策,如:花期仅重叠1—2天、阴雨天显著延长散粉期、花药雨天不开裂且已裂花药因复水作用闭合[8,29]。两种沙生独尾草的适宜分布范围相比,粗柄独尾草适生区较大且集中,这可能是因为粗柄独尾草在破碎化生境中形成了较好的局部环境适应性,其种子变轻、种翅增大,以增强种子的散布能力从而在不利生境中扩大其生存范围[30]。
新疆3种独尾草适宜分布区内的降水和温度均存在显著差异,阿尔泰独尾草受降水因素影响较大;而两种沙生独尾草则受气温因素影响较大(图2,图3)。其中,最干月降水量(Bio14)主要影响了阿尔泰独尾草的分布基准气候下该植物适生区内最干月降水的适宜范围是1—25mm,2001—2040年和2041—2080年的降水范围将略有增加。适宜的降水区间是植物对外界环境变化的缓冲应对,过多降水可能超出植物降水需求量的缓冲区间,降低植物生长的适宜性,从而将造成分布范围缩减。未来时段,阿尔泰独尾草适宜分布区减少的区域,如天山北麓和准噶尔盆地西部山地等,最干月降水量(Bio14)均有所增加,说明该因子的波动对其适宜分布具有主导性的限制(图2)。温度季节性变动指数(Bio4)主要影响了两种沙生独尾草的分布,基准气候下该因子的平均适宜范围为1150—1672,2001—2040年和2041—2080年分别为1490—1712和1524—1751。两种沙生类短命植物应对夏季高温,存在夏眠,未来气候情景下温度变动区间升高将导致两种植物适宜的生境缩减。相关研究也表明古尔班通古特沙漠短命植物的种子发芽受温度和光线的强调节作用[31]。因而,荒漠夏季酷热、冬季严寒的温度波动对两种沙生独尾草的生长、分布存在较大限制。2001—2040年和2041—2080年古特沙漠南缘的温度季节性变动指数(Bio14)显著增加(图2),两种独尾草的适宜分布也在这些区域呈破碎化减少趋势,尤其是异翅独尾草的高度适宜分布区将基本消失(图1)。温度季节性变动指数(Bio4)和最干月降水量(Bio14)也在一定程度上主导了山地植物阿尔泰独尾草与两种沙生独尾草环境生态位的分化(图3)。此外,本研究选取的地形因子坡度和坡向对新疆3种独尾草的适宜分布也有一定影响(表3)。阿尔泰独尾草多分布于天山北麓的阳坡(生长于南坡),常伴随天山云杉(生长于北坡)[11];异翅独尾草生态适应幅较宽不仅可与粗柄独尾草同生长于沙丘北坡或坡底丘间平地,也可生于坡顶[24]。
3.2 3种独尾草的适宜分布对未来气候变化的可能响应
气候变化将引起气温和降水量空间分布格局的改变,若超过了植物生态幅可能会使植物适生区面积减少[32]。例如,极端降水可能导致植物种子储存在土壤中或已经死亡[33],过量光热会对植株产生光破坏或光抑制,进而限制植物的生长[34]。在全球气候变化背景下,降水的波动变化和极端干旱事件也将对沙漠植被,特别是对短命草本种群产生较大的影响[35]。
未来气候变化情景下,3种独尾草的适宜分布面积均呈缩减趋势,阿尔泰独尾草为经度方向上的缩减,两种沙生独尾草为纬度方向上的缩减(图1)。2001—2040年和2041—2080年相比,气候变动较相似,均以准噶尔西部山地降水量增加、古尔班通古特沙漠南部温度变动加剧为主(图2)。以2001—2040年分析,阿尔泰独尾草62.07%的分布点发生气候波动,将造成适生区主要向天山北麓和准噶尔西部山地缩减;粗柄独尾草83.33%的分布点发生气候波动,将造成适生区向古尔班通古特沙漠南缘缩减;异翅独尾草88.24%的分布点发生气候波动,将造成适生区整体向沙漠南缘西端婆罗科努山的山前沙丘北坡缩减(图1)。相关研究也表明其它山地、荒漠植物未来气候变化情景下也呈现分布范围缩减趋势,如山地植物天山花楸在西北地区的适宜分布将向高海拔地区缩减[36];荒漠植物裸果木在我国西北荒漠区的适宜分布范围和最适宜分布区都将明显减少[37]。本研究3种植物适生分布区的变化与水温条件的变化紧密相关:2001—2040年和2041—2080年,研究区气候波动较大的区域,如准噶尔盆地中南部,将造成山地植物阿尔泰独尾草在古尔班通古特沙漠边缘的斑块适宜分布区消失,阿尔泰山中段区域边缘也呈现破碎化,而天山北麓、准噶尔西部山地未来时段气候波动较小,为该植物提供了较稳定、较集中的适宜生境,可作为资源管理和保护的重点区域(图1);古尔班通古特沙漠南缘,婆罗科努山中段区域,为沙生植物粗柄独尾草和异翅独尾草植物提供了连续、稳定的适宜生境(图1,图2)。
本研究模拟的新疆3种独尾草植物是新疆山地、荒漠区域的生态平衡与修复的先锋物种,3种植物分布的生态适宜性变化及其环境驱动机制研究具有重要的理论与现实意义。根据本研究,未来气候情境下,由于研究区气候波动,新疆3种独尾草的适宜分布总体均将呈缩减趋势,以适生区边缘及沙漠边缘斑块破碎化或消失为主,本研究的结果可为国内仅在新疆分布的3种独尾草植物资源的科学保护提供科学数据。
