盐度对香螺行为、鳃和肾抗氧化酶活性与组织结构的影响
2022-11-25张思研王绍军杨佳程朱建业毛俊霞王许波常亚青鹿志创郝振林
张思研, 王绍军, 杨佳程, 朱建业, 田 莹, 王 荦, 毛俊霞, 王许波, 常亚青, 鹿志创, 郝振林
盐度对香螺行为、鳃和肾抗氧化酶活性与组织结构的影响
张思研1, 王绍军2, 杨佳程1, 朱建业1, 田 莹1, 王 荦1, 毛俊霞1, 王许波1, 常亚青1, 鹿志创3, 郝振林1
(1. 大连海洋大学 农业农村部北方海水增养殖重点实验室, 辽宁 大连 116000; 2. 山东省渔业发展和资源养护总站, 山东 济南 250000; 3. 辽宁省海洋水产科学研究院, 辽宁 大连 116000)
为明确不同盐度胁迫下香螺的生理生态学响应及其适应机制, 本文采用行为学、组织学和生物化学手段研究了不同盐度条件下(16、20、24、28、32、36、40、44、48)香螺的行为、病理损伤以及抗氧化酶活性。结果显示, 在高盐和低盐胁迫下, 香螺出现应激反应, 摄食率下降, 死亡率升高。随盐度升高, 香螺的总抗氧化能力先升高后降低, 在盐度32时达到峰值; 香螺鳃的过氧化氢酶(CAT)活力和超氧化物歧化酶(SOD)活力呈先降低后升高的趋势; 肾的CAT和SOD活力呈先升高后降低的趋势。组织切片结果显示, 在高盐和低盐胁迫下, 香螺嗅检器和鳃丝的形态呈现显著变化, 黏液细胞数量均增加。上述结果表明, 盐度显著影响香螺的生理生态反应, 且香螺可通过行为变化、调节抗氧化酶以及改变组织形态结构从而适应外界盐度的变化。
香螺; 盐度; 生理胁迫; 行为; 抗氧化酶活性; 病理损伤
盐度是水产养殖重要的环境因子之一, 与养殖动物的生长、发育等生理活动息息相关, 外界环境的变化(如阴雨、干旱天气)都会一定程度上影响养殖水体的盐度, 从而影响养殖生物的生理状态[1-5]。盐度的变化会导致水产动物的生长存活、呼吸代谢等一系列生理反应发生变化。适当的盐度变化可以提高水产动物的生长繁殖及其肉质, 但当盐度超过耐受范围, 会引发应激反应甚至导致死亡[6-9]。因此, 探明盐度变化对水产动物的影响对开展人工增养殖具有重要的意义。
香螺()属软体动物门(Mollusca), 腹足纲(Gastropoda), 新腹足目(Neogastropoda), 蛾螺科(Buccinida), 主要分布在我国渤海和黄海海域, 朝鲜、日本海也有分布[10-11]。香螺肉质鲜美, 营养丰富, 市场需求量巨大, 目前市场价格已超过200元/kg, 对香螺野生资源的捕捞强度日益增加, 从而使香螺产量锐减。然而, 目前香螺规模化人工增养殖尚未突破, 其来源主要依靠野生资源, 因此, 探明香螺适宜生长的环境因子, 制定合理的增养殖模式, 尽快实现香螺的全人工增养殖迫在眉睫。目前我国关于香螺的研究主要集中在营养成分[12-13]、壳体[14-15]、繁殖[16]以及遗传多样性[17-18]等方面。国外对香螺的相关研究较少, 日本学者研究了北海道香螺与繁殖和摄食相关的迁移方式, 另外还有学者开展了有关香螺的繁殖习性的研究[19-20]。关于盐度对香螺行为等相关生理生态影响的研究尚未见报道。本研究拟采用生理学、组织学等手段研究了不同盐度条件下香螺的行为、相关组织抗氧化酶活性的变化以及组织形态的变化, 检测了香螺对盐度变化的耐受能力以及在不同盐度下的生理生态学响应, 旨在为明确香螺适应盐度变化的调节机制, 并为后期开展香螺的人工增养殖提供理论参考。
1 材料与方法
1.1 实验材料
实验用香螺取自辽宁省大连市旅顺口区黄海海域, 潮湿状态下运回大连海洋大学重点实验室暂养。香螺个体湿质量为(85.3±9.3) g。选取27个规格为50 L的水族箱(58 cm×44 cm×34 cm), 每个水族箱暂养100只香螺。暂养期间水温保持12 ℃, 水箱内含氧量为7.5 mg/L, 盐度为32。
1.2 实验方法
实验设置9个盐度组, 盐度分别为16、20、24、28、32(对照)、36、40、44、48, 将暂养在32盐度下的香螺直接放入相应盐度处理组中, 每个盐度设置3个平行, 每个平行组随机放入100只健康活跃的香螺。实验前将香螺断食2 d, 实验过程中尽量避免外界因素(如声音、光照等)对实验的干扰。香螺适应环境2 h后, 每个实验组内投入2 kg新鲜三角帆蚌()肉, 观察不同盐度下香螺的行为特征(持续观察120 h), 在第6 h时测定各实验组的摄食率, 并在各组内随机取20只香螺, 取其鳃、肾和嗅检器组织样品, 以为后期开展组织学和相关抗氧化酶活性研究。
1.3 测定指标与方法
1.3.1 行为学指标测定
摄食率: 香螺摄食量占香螺体重的百分比。
1.3.2 鳃和嗅检器石蜡切片观察
取新鲜的鳃和嗅检器保存于波恩氏液, 然后进行常规石蜡组织切片, HE染色等, 并在光学显微镜(Nikon E-CLIPSE)下观察组织形态学变化, 拍照记录。
1.3.3 酶活性测定
香螺鳃和肾的总抗氧化能力(T-AOC)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)活力的测定均采用南京建成生物工程研究所试剂盒进行, 具体参考其说明书。
1.4 数据处理
使用SPSS22.0和OriginPro 2017软件对6 h内盐度变化所导致的香螺摄食率及鳃和嗅检器的组织切片进行分析, 用单因素方差分析统计香螺鳃丝间距、嗅检器丝间距以及鳃和肾抗氧化能力(T-AOC)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)的活力变化(<0.05为差异显著)。
2 结果
2.1 行为观察
2.1.1 香螺的摄食行为
6 h时不同盐度下香螺的摄食率见图1。当盐度高于16时, 香螺开始摄食, 摄食率随盐度上升逐渐升高; 当盐度为28时, 摄食率最高为50.11%, 随后香螺摄食率随盐度上升而下降, 当盐度高于44时香螺停止摄食, 摄食率为0。
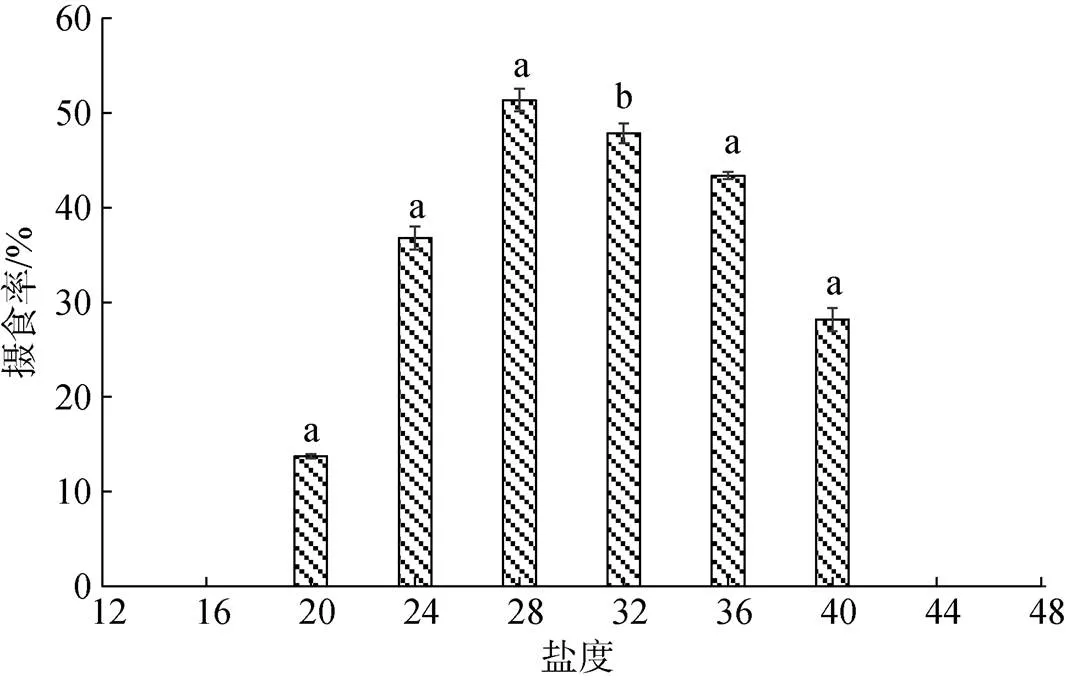
图1 6 h时不同盐度对香螺摄食率的影响
注: 不同小写字母表示不同盐度下香螺摄食率对于32盐度时差异显著(<0.05)
将不同盐度下香螺摄食率进行ks检验和正态分布拟合分析, 拟合方程为=0.000 84–0.098 63+ 4.149 72–68.31+382.12(其中为摄食率,为盐度,2值为 0.981 7)。拟合曲线见图2, 香螺的摄食率随盐度的升高呈先上升后下降的趋势, 由拟合方程可知, 当盐度为31.5时, 香螺摄食率最高。

图2 不同盐度对香螺摄食率影响曲线
2.1.2 香螺的死亡率
6 h时不同盐度下香螺的死亡率如图3所示。由表可知, 在高盐胁迫和低盐胁迫下, 香螺存在一定的应激行为。对照组中, 香螺比较安静, 喜欢待在水底。当香螺所处海水盐度为16、20、24时, 6 h后开始出现脱吸现象, 并分别在24 h、48 h、72 h全部死亡。脱吸状态时, 香螺足部出现水肿状态, 严重时软体部无法完全缩入壳中, 死亡状态时, 腹足僵硬, 翻出螺壳外, 吻管缩回壳内, 用玻璃棒轻触无生命迹象。在盐度为28时, 香螺在6 h内处于活跃期, 此时香螺活动量显著增大, 尤其喜欢向水面攀爬。当香螺处于海水盐度为44、48时, 6 h后出现脱吸, 并分别于96 h、24 h全部死亡。

图3 6 h时不同盐度对香螺死亡率的影响
2.2 主要抗氧化指标的变化
2.2.1 盐度胁迫下T-AOC的变化
6 h时不同盐度胁迫下香螺鳃和肾的抗氧化能力(T-AOC)见图4。当盐度为32时, 香螺鳃和肾的T-AOC值最高, 且显著高于其他盐度梯度水平(<0.01)。当盐度上升, 鳃和肾的T-AOC均呈先上升后下降的趋势, 且在盐度32时达到最大值。香螺鳃和肾的T- AOC在高盐低盐胁迫下的变化趋势一致, 但是在32与28, 32与36盐度水平下鳃的T-AOC存在极显著差异(<0.01), 肾的T-AOC在盐度为32与28之间不存在显著差异(>0.05), 在盐度为32和36之间存在极显著差异(<0.01)。
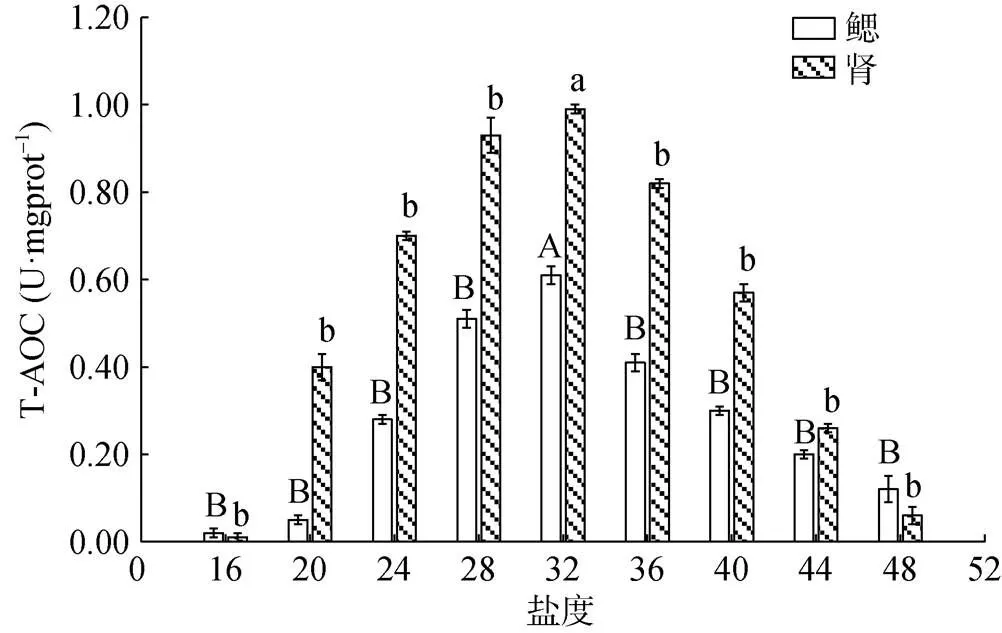
图4 6 h时不同盐度条件对香螺鳃和肾的抗氧化能力(T- AOC)的影响
注: 不同大小写字母表示不同盐度下鳃或肾对于32盐度时抗氧化能力(T-AOC)差异显著(<0.05)
2.2.2 盐度胁迫下CAT的变化
6 h时不同盐度胁迫下香螺的鳃和肾的过氧化氢酶(CAT)活力变化见图5。随着盐度逐渐上升, 鳃的CAT活力呈先下降后上升的趋势, 肾的CAT活力呈先上升后下降的趋势, 且均在盐度为32时达到最值。盐度为28和36时鳃的CAT活力与对照组32相比均存在极显著差异(<0.01)。盐度为28和36时肾的CAT活力与对照组32相比均存在极显著差异(<0.01), 盐度在28和36之间不存在显著差异(>0.05)。当盐度为32时, 鳃的CAT活力值最低, 且显著低于其他盐度梯度水平(<0.01), 肾的CAT活力值最高, 且显著高于其他盐度梯度水平(<0.01)。
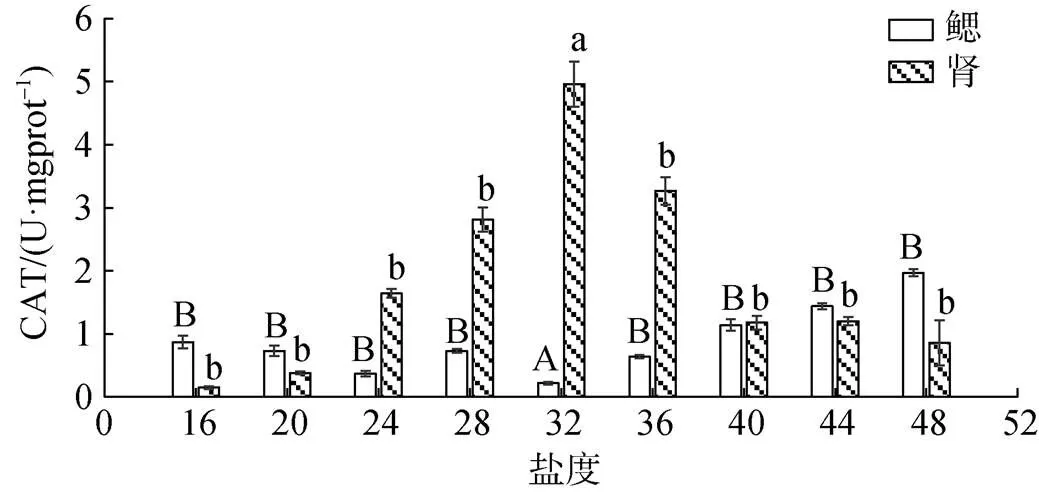
图5 6 h时不同盐度条件对香螺鳃和肾的过氧化氢酶(CAT)的影响
注: 不同大小写字母表示不同盐度下鳃或肾对于32盐度时过氧化氢酶(CAT)差异显著(<0.05)
2.2.3 盐度胁迫下SOD的变化
6 h时不同盐度胁迫条件下香螺鳃和肾的超氧化物岐化酶(SOD)活力变化见图6。随着盐度逐渐上升, 肾的超氧化物歧化酶(SOD)活力呈先上升后下降的趋势。当盐度为28时, 鳃的超氧化物歧化酶(SOD)活力值最低。盐度为28和36时鳃的超氧化物歧化酶(SOD)活力与对照组32相比均存在极显著差异(<0.01)。盐度为32和28时肾的超氧化物歧化酶(SOD)活力不存在显著差异(>0.05), 盐度为32和36时肾的超氧化物歧化酶(SOD)活力存在极显著差异(<0.01)。
2.3 组织学观察
2.3.1 盐度胁迫下鳃组织学的观察比较
6 h时不同盐度胁迫下香螺鳃的组织形态变化见图7。香螺鳃丝间距在高低盐条件下均呈先扩大后缩小的趋势, 16盐度水平下鳃丝间隔最小, 40盐度水平鳃丝间隔最大。香螺的鳃丝在32盐度水平下呈正常并拢状态, 鳃丝组织结构完整, 呼吸上皮细胞和泌氯细胞排列整齐。16盐度水平下香螺鳃丝顶端肿胀, 内有游离的泌氯细胞。在20、24和28低盐度水平下, 部分鳃丝细胞排列混乱, 鳃丝呼吸上皮细胞明显增厚, 鳃丝边缘模糊。在40、44和48高盐度水平下, 香螺鳃丝皱缩, 顶端略有肿胀, 部分鳃丝呼吸上皮细胞脱落、坏死, 鳃丝边缘呈锯齿状。黏液细胞数量在低盐度和高盐度条件下均增加。

图6 6 h时不同盐度条件对香螺鳃和肾的超氧化物歧化酶(SOD)的影响
注: 不同大小写字母表示不同盐度下鳃或肾对于32盐度时超氧化物歧化酶(SOD)差异显著(<0.05)
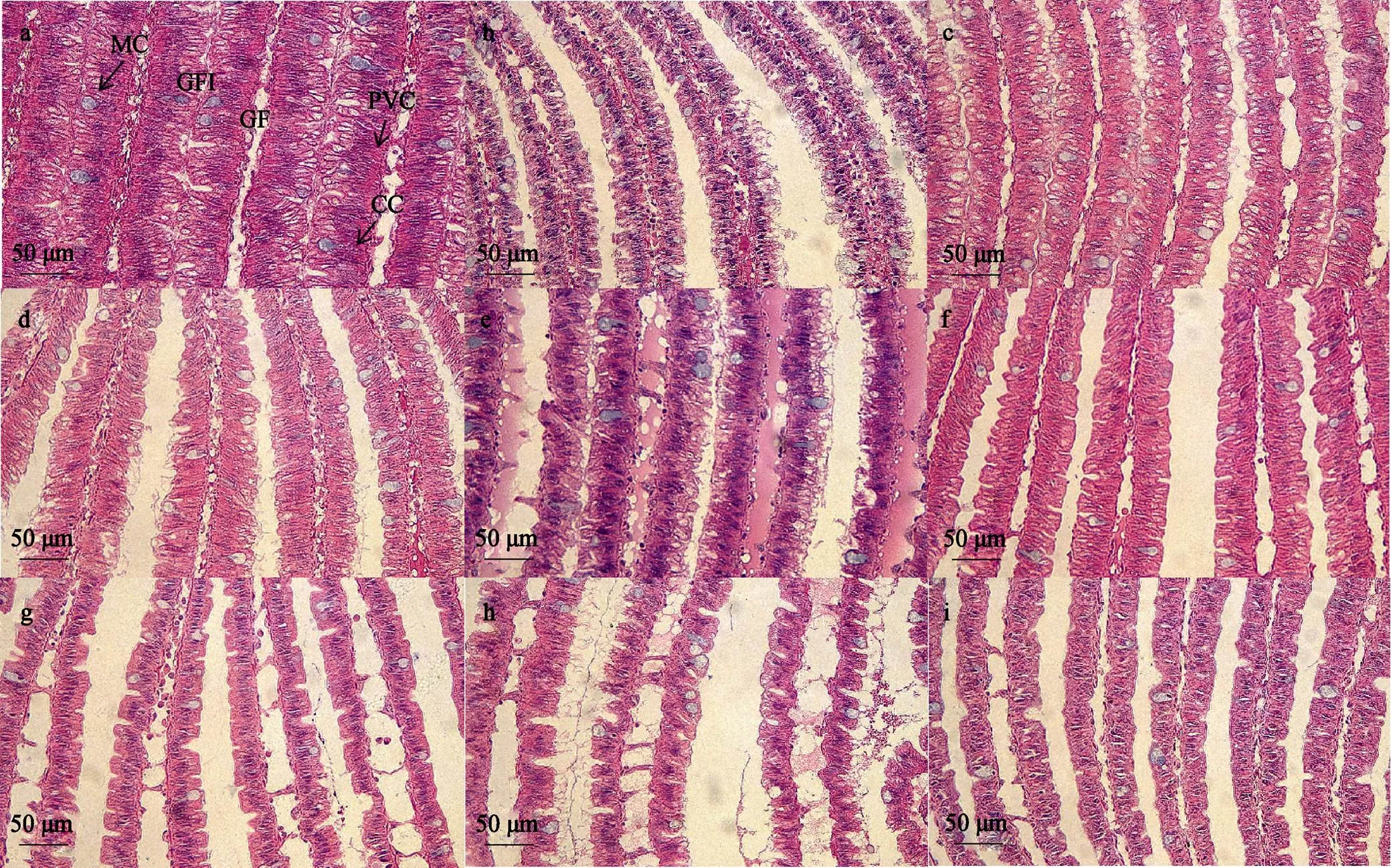
图7 6 h时不同盐度条件下香螺鳃组织形态
注: a: 盐度16; b: 盐度20; c: 盐度24; d: 盐度28; e: 盐度32; f: 盐度36; g: 盐度40; h: 盐度44; i: 盐度48
注: GF: 鳃丝; MC: 黏液细胞; GFI: 鳃丝间隔; PVC: 呼吸上皮细胞; CC: 泌氯细胞
2.3.2 盐度胁迫下嗅检器组织的观察比较
6 h时不同盐度胁迫下香螺嗅检器组织形态变化见图8。香螺嗅检器丝间隔随盐度上升而变大, 在20盐度水平下香螺嗅检器丝间隔最小, 40盐度水平下香螺嗅检器丝间隔最大, 在40、44、48高盐水平下, 嗅检器丝间隔无太大变化。鳃丝间的结缔组织随盐度增加而逐渐减少, 高盐条件下黏液细胞的体积变小。
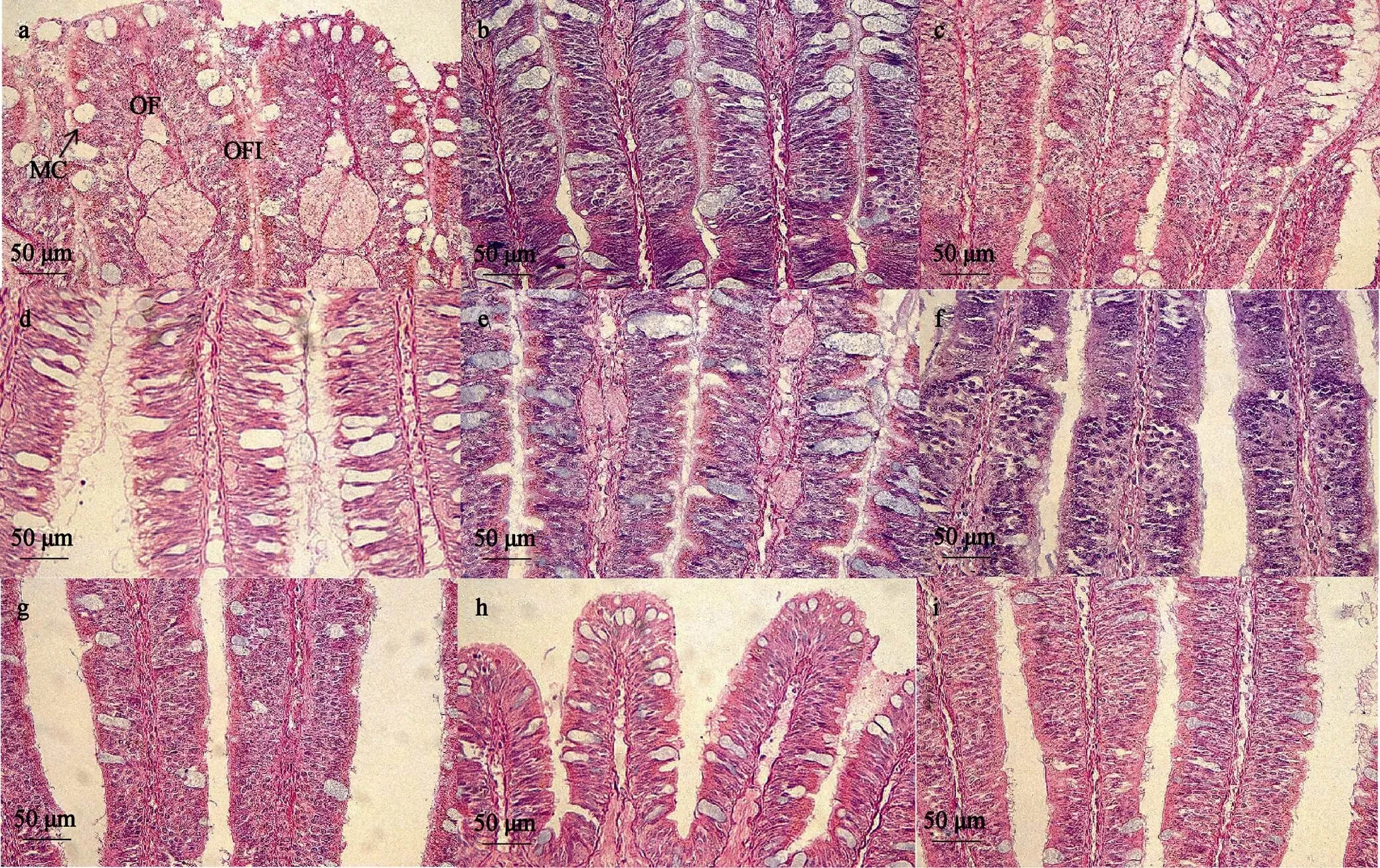
图8 6 h时不同盐度条件下香螺嗅检器组织形态
注: a: 盐度16; b: 盐度20; c: 盐度24; d: 盐度28; e: 盐度32; f: 盐度36; g: 盐度40; h: 盐度44; i: 盐度48
注: OF: 嗅检器丝; MC: 黏液细胞; OFI: 嗅检器丝间隔
2.3.3 盐度胁迫下黏液细胞的变化
6 h时不同盐度下黏液细胞数量变化见图9。鳃和嗅检器的黏液细胞数量在高盐和低盐条件下均呈现先上升后下降的趋势。当盐度为24时, 香螺鳃的黏液细胞数量最多。除盐度28和32水平外, 剩余盐度条件下香螺的鳃和嗅检器黏液细胞数量均存在极显著差异(<0.01)。
3 讨论
3.1 盐度对香螺行为的影响
盐度是海水一项重要理化指标, 也是贝类摄食、呼吸等生理活动的重要影响因子之一[21]。对水产动物而言, 盐度变化主要影响其渗透压调节, 进而影响其基础代谢, 水产动物对盐度均有一定的适应范围, 当超出这个范围, 机体会通过应急响应进行调节以适应盐度变化[22]。有研究发现, 低盐条件下, 贝类为了保护机体, 会关闭贝壳; 高盐条件下, 由于环境盐度超出贝类的承受范围, 机体无法进行正常的生理活动, 最终死亡[23-26]。本研究结果与之相似, 本研究发现, 香螺的摄食率随盐度从16上升而升高, 行动逐渐活跃。当盐度为28时, 摄食率最高, 此后随着盐度的升高香螺摄食率降低, 盐度44时停止摄食。推测认为出现这种现象的原因是香螺为适应低盐条件会降低自身的代谢速率, 并伴随闭厣、行动缓慢等行为, 与外界交换水量降低, 进而导致摄食率降低; 高盐胁迫下, 香螺部分组织细胞可能受损, 无法进行正常的摄食和代谢活动。盐度为28时, 香螺的摄食率最高, 推测有可能是因为其产生应激反应, 进行报复性摄食以维持机体渗透压平衡。通过拟合曲线可知, 在盐度为31.5时香螺摄食率最高为46.675%, 此时为其摄食的最适盐度, 这与周学家等关于香螺最适盐度为31~32.5的结论一致[27]。

图9 6 h时不同盐度条件下香螺鳃和嗅检器的黏液细胞数量变化
注: 不同大小写字母表示不同盐度下鳃或嗅检器对于32盐度时和嗅检器的黏液细胞数量变化差异显著(<0.05)
此外本实验通过对比不同盐度水平下香螺的死亡时间和死亡率发现, 香螺对高盐度的耐受性优于对低盐度的耐受性, 与管角螺()稚贝对高盐度有较强的耐受力一致[28], 分析认为出现这种现象的原因与物种间的生活习性差异有关, 香螺的栖息环境一般比较稳定, 因此较少会遇到由于陆地活动或者雨水影响导致盐度大幅度变化的情况。
3.2 盐度对香螺抗氧化能力的影响
盐度改变通常会导致生物体内的多种生理应激反应, 这种反应多与相关活性氧分子的过量产生有关, 而抗氧化能力的提高是消除超氧自由基的重要途径。总抗氧化能力(total antioxidant capacity, T-AOC)是用于衡量机体抗氧化系统状态的综合性指标[29]。SOD与CAT能有效清除体内的超氧阴离子自由基 (O2–) 、游离氧 (O) 和H2O2等活性氧物质, 减少细胞毒性对机体产生的危害, 是机体抗氧化防御系统的重要组成部分[30-31]。
不适宜的盐度环境会降低贝类的免疫力、抵抗力和新陈代谢率等[32-34]。多名学者开展了盐度对水生动物抗氧化能力的研究, 发现高盐或低盐胁迫对水生动物的抗氧化能力均有显著影响[35-36]。盐度的变化会影响水生动物的渗透压调节, 有研究表明, 渗透压的改变会影响水生动物的免疫酶活性, 进而影响免疫系统的功能。不良环境刺激时, 机体免疫系统响应, 其中CAT和SOD起重要作用。此外, 水生动物为适应非等渗环境需要耗能, 这一过程也会导致机体免疫酶活力的变化[37]。本实验中, 随着盐度的上升, 香螺鳃和肾的总抗氧化能力均呈先升高后降低的趋势, 且在盐度32时达到最高。可以初步判断在不适盐度时, 香螺处于被动应激状态, 不适的盐度环境会使香螺消耗大量能量来调整渗透压, 同时, 盐度胁迫使香螺体内大量ROS堆积, 机体受到氧化损伤, 酶活性受到抑制, 进而影响了香螺的总抗氧化能力。
香螺鳃的CAT和SOD活力随盐度的升高均呈先下降后上升的趋势。这与盐度对大竹蛏()、锯缘青蟹()等水生动物的酶活力影响相一致[38-39]。徐立红等研究表明, 在轻度逆境胁迫下, 生物体的CAT、SOD活性会被诱导, 而在重度逆境胁迫时, 其活性会被抑制[40]。推断认为在轻度盐度胁迫下, ROS含量增多, 机体为保持氧化系统的动态平衡而产生应激反应, CAT和SOD活力显著增加, 做出快速应答以清除ROS; 重度盐度胁迫下香螺机体处于过氧化状态, 受到氧化损伤, 部分相关酶合成异常, 活性降低。与对照组相比, 随着盐度升高, 香螺肾的CAT和SOD活力总体呈先上升后下降的趋势, 这与本实验中鳃的抗氧化酶活性变化相反, 推测这种差异出现的原因可能是香螺鳃和肾之间对盐度胁迫的应答具有一定的组织差异性, 具体的变化原因和作用机制还有待进一步验证。
3.3 盐度对香螺鳃和嗅检器组织形态的影响
鳃是贝类体内用以呼吸、摄食以及渗透压调节的重要器官。Carmona等研究发现, 鳃丝会随着外界盐度的改变而发生相应的适应性变化, 因此, 鳃的组织形态变化是研究盐度对水生生物影响的重要指标[41]。嗅检器是香螺用以判读环境变化的主要器官, 在贝类生物中起着重要的化学感受作用, 如摄食定位、调整呼吸、调控生殖、感光等[42-45]。很多学者开展了盐度对水产动物鳃的影响的研究, 在对紫石房蛤()、珍珠龙胆石斑鱼(♀×♂)幼鱼、红鳍东方鲀()等的研究中, 都发现了盐度变化显著影响水生生物鳃组织结构的现象[30, 46-47]。本研究中, 香螺在低盐胁迫下鳃的丝间距总体呈扩大趋势, 在高盐胁迫下丝间距呈先扩大后缩小的趋势, 与高盐度组相比, 低盐组的鳃丝间距更小, 鳃的呼吸上皮细胞增厚, 鳃丝略有肿胀。低盐胁迫下, 鳃丝的变化有利于鳃与水体接触面积增大, 从而大量吸收水中的无机盐离子, 以适应低渗环境; 高盐胁迫下, 鳃丝间距的增加可以增大滤水量, 从而增大水氧交换量, 以调节体内渗透压的平衡。鳃丝间距在盐度为16时出现最小值且显著低于其他组, 推测在此低盐胁迫下, 鳃组织已经受到了一定程度的破坏, 无法正常地进行调节。
黏液细胞是一种常见于水生动物体内的腺细胞, 可分泌黏液, 其中含有多种活性物质, 具有免疫作用, 并可以促进离子的交换, 黏液细胞的数量可以表示香螺的应激程度[48-51]。面对盐度胁迫, 黏液细胞会产生保护性反应, 但超出其保护限度时, 黏液细胞会受到破坏[52]。本研究中, 随着盐度从对照组升高或降低, 香螺鳃和嗅检器的黏液细胞数量均呈现先增加后减少的趋势。分析认为在轻度不适的盐度条件下, 香螺增加黏液细胞数量并分泌黏液, 通过离子交换适应外界渗透压的改变, 同时保护组织的完整性; 当香螺处于极端不适盐度时, 推测可能由于过度应激造成了不可逆的损伤, 导致其黏液细胞合成异常, 数量下降。
4 结论
盐度显著影响香螺的生理生态反应, 在高盐和低盐胁迫下, 香螺出现应激反应, 摄食率下降, 死亡率升高; 香螺鳃和肾的T-AOC、CAT以及SOD活力均产生显著变化; 香螺嗅检器和鳃丝的形态呈现显著变化, 黏液细胞数量均增加。因此香螺可通过行为变化、调节抗氧化酶以及改变组织形态结构从而适应外界盐度的变化。
[1] ZHAO Q. Effect of salinity on regulation mechanism of neuroendocrine-immunoregulatory network in[J]. Fish and Shellfish Immunology, 2016, 49: 396-406.
[2] 周忠凯. 拟穴青蟹几丁质酶基因克隆及其在发育, 蜕壳与环境胁迫中的表达分析[D]. 杭州: 浙江大学, 2018.
ZHOU Zhongkai. The chitinase genes cloning and expression analysis during development, molting and environment stresses in mud crab[D]. Hangzhou: Zhejiang University, 2018.
[3] LI C, LI N, DONG T T, et al. Analysis of differential gene expression inunder High salinity stress[J]. Aquaculture Reports, 2020, 18: 100423.
[4] 何竺柳. 低盐度胁迫对罗氏沼虾生长和肉质的影响[D].舟山: 浙江海洋大学, 2018.
HE Zhuliu. Effects of low salinity stress on growth and meat quality of[D]. Zhoushan: Zhejiang Ocean University, 2018.
[5] 马金武. 三疣梭子蟹不同盐度下血淋巴理化指标分析及Na~+/H~+-exchanger和基因的克隆和功能研究[D]. 上海: 上海海洋大学, 2016.
MA Jinwu. Physicochemical index analysis of haemolymph and cloning and functional study of Na+/H+- exchanger andgenes in swimming crab () under different salinity[D]. Shanghai: Shanghai Ocean University, 2016.
[6] 程亚美, 赵金良, 宋凌元, 等. 盐度、碱度对尼罗罗非鱼生长和肌肉品质的影响[J]. 水产科学, 2020, 39(3): 341-349.
CHENG Yamei, ZHAO Jinliang, SONG Lingyuan, et al. Effect of salinity and alkalinity on growth performance and muscle quality of Nile Tilapia[J]. Fisheries Science, 2020, 39(3): 341-349.
[7] 鲜博, 高建操, 徐钢春, 等. 盐度对刀鲚生长、抗氧化应激和渗透压调节能力的影响[J]. 海洋湖沼通报, 2020(2): 152-159.
XIAN BO, GAO Jiancao, XU Gangchun, et al. Influence of salinity on the growth, digestive ability, antioxidant ability and osmoregulation of[J]. Transactions of Oceanology and Limnology, 2020(2): 152-159.
[8] 丁红兵, 李浩宇, 陈义华, 等. 高盐对缢蛏生长存活、Na~(+)/K~(+)-ATPase活性及能量代谢相关指标的影响[J]. 上海海洋大学学报, 2022, 31(4): 831-838.
DING Hongbing, LI Haoyu, CHEN Yihua, et al. Effects of high salinity on growth and survival, Na+/K+- ATPase activity and energy metabolism related indexes of razor clam[J]. Journal of Shanghai Ocean University, 2022, 31(4): 831-838.
[9] 赵乃乾, 肖述, 刘浩翔, 等. 盐度对牡蛎“华南1号”早期发育与生长的影响[J]. 中国海洋大学学报(自然科学版), 2020, 50(8): 53-64.
ZHAO Naiqian, XIAO Shu, LIU Haoxiang, et al. Effect of salinity on early development and growth of oyster “South China No.1”[J]. Periodical of Ocean University of China, 2020, 50(8): 53-64.
[10] 蔡清海. 香螺的资源保护措施及加工方法[J]. 中国水产, 2001(10): 73.
CAI Qinghai. The recourse protection and processing method of[J]. China Fisheries, 2001(10): 73.
[11] 郭栋, 刘修泽, 王爱勇, 等. 辽东湾香螺资源的分布研究[J]. 水产科学, 2015, 34(11): 718-721.
GUO Dong, LIU Xiuze, WANG Aiyong, et al. Stock distribution ofin Liaodong Bay[J]. Fisheries Science, 2015, 34(11): 718-721.
[12] 何晋浙. 香螺生物体内的微量元素分析及食用安全性[J]. 食品科学, 2010, 31(8): 181-184.
HE Jinzhe. Analysis of trace elements and edibleness oftissue[J]. Food Science, 2010, 31(8): 181-184.
[13] 郝振林, 王煜, 于洋洋, 等. 香螺肌肉营养成分分析及评价[J]. 大连大学学报, 2016, 37(6): 66-70.
HAO Zhenlin, WANG Yu, YU Yangyang, et al. Analysis and evaluation of nutritive composition in the muscle of(gastropoda: bccinidae)[J]. Journal of Dalian University, 2016, 37(6): 66-70.
[14] 邵林, 王中华, 郭庆梅, 等. 贝壳类药材海螺壳和香螺壳的性状显微鉴别[J]. 中国海洋药物, 2017, 36(4): 18-24.
SHAO Lin, WANG Zhonghua, GUO Qingmei, et al. Comparative study on morphology and microscopy of Rapanae Concha and Neotuneae Concha[J]. Chinese Journal of Marine Drugs, 2017, 36(4): 18-24.
[15] 赵杰, 丁晓非, 高山, 等. 香螺壳体的结构特征分析[J]. 材料科学与工程学报, 2004(5): 644-646.
ZHAO Jie, DING Xiaofei, GAO Shan, et al. Structure characteristics of the Sshell of the conch[J]. Journal of Materials Science and Engineering, 2004(5): 644-646.
[16] 于洋洋, 李靖, 吴薇, 等. 香螺繁殖生物学及人工繁育技术研究进展[J]. 河北渔业, 2019(4): 54-56.
YU Yangyang, LI Jing, WU Wei, et al. An introduction to current research of reproductivebiology and artificialpropagation of[J]. Hebei Fisheries, 2019(4): 54-56.
[17] 王煜. 黄、渤海海域香螺遗传多样性研究[D]. 大连: 大连海洋大学, 2018.
WANG Yu. Study of genetic diversity of[D]. Dalian: Dalian Ocean University, 2018.
[18] 张旦旦, 王煜, 李卓, 等. 香螺线粒体COXⅠ和CYTB基因遗传多样性研究[J]. 大连海洋大学学报, 2021, 36(4): 612-619.
ZHANG Dandan, WANG Yu, LI Zhuo, et al. Genetic diversity of mitochondrial COXⅠ and CYTB genes in neptune whelk[J]. Journal of Dalian Ocean University, 2021, 36(4): 612-619.
[19] KATSUAKI F, SHIGERU N. Migration pattern of, with special reference to relations with reproductive and feeding activities[J]. Japanese Journal of Ecology, 1994, 44: 331-338.
[20] AZUMA N. Polymorphic microsatellite markers isolated from the neptune whelk[J]. Molecular Ecology Resources, 2009, 9(1): 120-121.
[21] HANS U R, MULOT M, MERINO L, et al. Effects of salinity-changing rates on filtration activity of mussels from two sites within the Baltic Mytilus hybrid zone: the brackish great belt (Denmark) and the low saline central Baltic sea[J]. Open Journal of Marine Science, 2014, 4(2): 101-109.
[22] 尤仲杰, 陆彤霞, 马斌, 等. 盐度对墨西哥湾扇贝幼虫和稚贝生长与存活的影响[J]. 动物学杂志, 2003(3): 58-60.
YOU Zhongjie, LU Tongxia, MA Bin, et al. Effect of seawater salinity on the growth and survival of Argopecten irradians concentricus Larvae and Juveniles[J]. Chinese Journal of Zoology, 2003(3): 58-60.
[23] 彭建华, 陈文祥, 栾建国, 等. 温度、pH对二种淡水贝类滤水率的影响[J]. 动物学杂志, 2004(6): 2-6.
PENG Jianhua, CHEN Wenxiang, LUAN Jianguo, et al. The effects of temperature and pH on the filtration rates of two species of freshwater mollusks[J]. Chinese Journal of Zoology, 2004(6): 2-6.
[24] 许巧情, 刘俊, 冯抗抗. 温度对橄榄蛏蚌滤水率的影响[J]. 中国水产科学, 2005(2): 207-210.
XU Qiaoqian, LIU Jun, FENG Kangkang. Influence of temperature on clearance rate of[J]. Journal of Fishery Sciences of China, 2005(2): 207- 210.
[25] 栗志民, 刘志刚, 徐法军, 等. 温度、盐度、pH和饵料密度对皱肋文蛤清滤率的影响[J]. 渔业科学进展, 2011, 32(4): 55-61.
LI Zhimin, LIU Zhigang, XU Fajun, et al. Effects of temperature, salinity, pH, and microalgae density on clearance rates of[J]. Progress in Fishery Sciences,2011, 32(4): 55-61.
[26] 栗志民, 刘志刚, 邓海东. 温度和盐度对企鹅珍珠贝清滤率、滤食率、吸收率的影响[J]. 水产学报, 2011, 35(1): 96-103.
LI Zhimin, LIU Zhigang, DENG Haidong. Effects of temperature and salinity on clearance rate, filtration rate and absorption efficiency of Pteria penguin[J]. Journal of Fisheries of China, 2011, 35(1): 96-103.
[27] 周学家, 张玉玺, 刘信艺, 等. 山东近海香螺资源的分布研究[J]. 齐鲁渔业, 1995(1): 8-10.
ZHOU Xuejia, ZHANG Yuxi, LIU Xinyi, et al. On distributing characteristics ofin Shandong coastal waters[J]. Shandong Fisheries, 1995(1): 8-10.
[28] 罗杰, 曹伏君, 刘楚吾, 等. 管角螺稚贝在不同盐度下的生长和存活研究[J]. 热带海洋学报, 2011, 30(6): 90-96.
LUO Jie, CAO Fujun, LIU Chuwu, et al. Study on the growth and survival of(Gmelin) Juvenile at different salinity[J]. Journal of Tropical Oceanography, 2011, 30(6): 90-96.
[29] 温久福, 蓝军南, 周慧, 等. 盐度对花鲈幼鱼消化酶和抗氧化系统的影响[J]. 动物学杂志, 2019, 54(5): 719-726.
WEN Jiufu, LAN Junnan, ZHOU Hui, et al. Effects of salinity on digestive enzymes and antioxidant system of juvenile[J]. Chinese Journal of Zoology, 2019, 54(5): 719-726.
[30] 王怡, 胡婉彬, 李家祥, 等. 急性盐度胁迫对紫石房蛤()鳃组织结构及4种酶活性的影响[J]. 中国农业科技导报, 2016, 18(5): 178-186.
WANG Yi, HU Wanbin, LI Jiaxiang, et al. Effects of acute salinity stress on gill structure and four enzyme activities in[J]. Journal of Agricultural Science and Technology,2016, 18(5): 178-186.
[31] 亢玉静, 郎明远, 赵文. 水生生物体内抗氧化酶及其影响因素研究进展[J]. 微生物学杂志, 2013, 33(3): 75-80.
KANG Yujing, LANG Mingyuan, ZHAO Wen. Advance in antioxidant enzymes and its effect factors in aquatic organisms[J]. Journal of Microbiology, 2013, 33(3): 75-80.
[32] BERGER V J, KHARAZOVA A D. Mechanisms of salinity adaptations in marine molluscs[J]. Hydrobiologia, 1997, 355(1/3): 115-126.
[33] 林志华, 柴雪良, 方军, 等. 硬壳蛤对环境因子适应性试验[J]. 宁波大学学报(理工版), 2002(1): 19-22.
LIN Zhihua, CHAI Xueliang, FANG Jun, et al. An aptitude test on the environmental habits of Mercenaria mercenaria[J]. Journal of Ningbo University (Natural Science & Engineering Edition), 2002(1): 19-22.
[34] 阎希柱, 王桂忠, 李少菁. 盐度对九孔鲍能量收支的影响[J]. 生态学杂志, 2009, 28(8): 1520-1524.
YAN Xizhu, WANG Guizhong, LI Shaojing. Effects of water salinity on energy budget ofaquatilis[J]. Chinese Journal of Ecology, 2009, 28(8): 1520-1524.
[35] 龙晓文, 吴仁福, 侯文杰, 等. 水体盐度对雌性三疣梭子蟹生长、卵巢发育、渗透压调节、代谢和抗氧化能力的影响[J]. 水产学报, 2019, 43(8): 1768-1780.
LONG Xiaowen, WU Renfu, HOU Wenjie, et al. Effects of water salinity on the growth, ovarian development, osmoregulation, metabolism and antioxidant capacity of adult female swimming crab ()[J]. Journal of Fisheries of China, 2019, 43(8): 1768-1780.
[36] 张晨捷, 张艳亮, 高权新, 等. 低盐胁迫对黄姑鱼幼鱼肝脏抗氧化功能的影响[J]. 南方水产科学, 2015, 11(4): 59-64.
ZHANG Chenjie, ZHANG Yanliang, GAO Quanxin, et al. Effect of low salinity stress on antioxidant function in liver of juvenile[J]. South China Fisheries Science, 2015, 11(4): 59-64.
[37] 谭春明, 赵旺, 吴开畅, 等. 盐度胁迫对方斑东风螺免疫酶活性的影响[J]. 水产科学, 2020, 39(5): 677- 684.
TAN Chunming, ZHAO Wang, WU Kaichang, et al. Effects of salinity stress on activities of enzymes Rrelated to immune of snail[J]. Fisheries Science, 2020, 39(5): 677-684.
[38] 张雨, 陈爱华, 曹奕, 等. 盐度胁迫对大竹蛏成活率与抗氧化酶活性的影响[J]. 海洋渔业, 2018, 40(3): 351-359.
ZHANG Yu, CHEN Aihua, CAO Yi, et al. Effects of salinity stress on survival and antioxidant enzymes activities of[J]. Marine Fisheries, 2018, 40(3): 351-359.
[39] 陈宇锋, 艾春香, 林琼武, 等. 盐度胁迫对锯缘青蟹血清及组织、器官中PO和SOD活性的影响[J]. 台湾海峡, 2007(4): 569-575.
CHEN Yufeng, AI Chunxiang, LIN Qiongwu, et al. Effect of salinity stress on the activities of phenoloxidase and superoxide dismutase of the serum, tissue and organ of mud crab,[J]. Journal of Oceanography in Taiwan Strait, 2007(4): 569-575.
[40] 徐立红, 张甬元, 陈宜瑜. 分子生态毒理学研究进展及其在水环境保护中的意义[J]. 水生生物学报, 1995, 19(2): 171-185.
XU Lihong, ZHANG Yongyuan, CHEN Yiyu. The advances of molecular ecotoxicology and its significance in water environment protection[J]. Acta Hydrobiologica Sinica, 1995, 19(2): 171-185.
[41] Carmona R, García-Gallego M, Sanz A, et al. Chloride cells and pavement cells in gill epithelia of Acipenser naccarii: ultrastructural modifications in seawater- acclimated specimens[J]. Journal of Fish Biology, 2004, 64(2): 553-566.
[42] CHITRAMVONG Y P, KRUATRACHUE M, UPATHAM E S, et al. The pallial organs of(Gastropoda: Haliotidae)[J]. Science Asia, 2002, 28: 17-23.
[43] COPELAND M. The olfactory reactions and organs of the marine snails(say) and(Linn.)[J]. Journal of Experimental Zoology, 2005, 25(1): 177-227.
[44] KRAEMER L R. Supra branchial and branchial shelves of bivalved molluscs: structural functional context of visceral ganglion, osphradium and branchialnerves[J]. American Zoologist, 1979, 19(3): 959.
[45] 甘洋. 方斑东风螺嗅觉及其对海洋酸化的响应[D]. 厦门: 厦门大学, 2017.
GAN Yang. Olfaction and its response under ocean acidification of[D]. Xiamen: Xiamen University, 2017.
[46] 刘龙龙, 罗鸣, 陈傅晓, 等. 盐度对珍珠龙胆石斑鱼幼鱼生长及鳃肾组织学结构的影响[J]. 大连海洋大学学报, 2019, 34(4): 505-510.
LIU Longlong, LUO Ming, CHEN Fuxiao, et al. Effects of salinity on growth, and gill and kidney histological structures of juvenile pearl gentian grouper[J]. Journal of Dalian Ocean University, 2019, 34(4): 505-510.
[47] 孙梦蕾, 蒋洁兰, 王莉苹, 等. 低盐胁迫下红鳍东方鲀幼鱼鳃、肾和肠组织结构的变化[J]. 广东海洋大学学报, 2016, 36(6): 38-43.
SUN Menglei, JIANG Jielan, WANG Liping, et al. Structural changes in gill, kidney and intestine of juvenileunder low salinity treatment[J]. Journal of Guangdong Ocean University,2016, 36(6): 38-43.
[48] 孙虎山, 王宜艳, 王平, 等. 栉孔扇贝外套膜和鳃黏液细胞的类型与分布[J]. 中国水产科学, 2002(4): 315-317, 390.
SUN Hushan, WANG Yiyan, WANG Ping, et al. Types and distribution of mucous cells in mantle and gill of[J]. Journal of Fishery Sciences of China,2002(4): 315-317, 390.
[49] 孙虎山, 李光友. 双壳贝类参与免疫防御的体液因子[J].海洋科学, 2001, 25(4): 34-36.
SUN Hushan, LI Guangyou. Humoral factors of Bivalves participating in immunity defence[J]. Marine Sciences, 2001, 25(4): 34-36.
[50] HARRIA J E, WATSON A, HUNT S. Histochemical analysis of mucous cells in the epidermis of brown trout[J]. Journal of Fish Biology, 1973, 5(3): 345-351.
[51] 方展强, 邱玫, 王春凤. 剑尾鱼鳃结构的光镜、扫描和透射电镜观察[J]. 电子显微学报, 2004(5): 553-559.
FANG Zhanqiang, QIU Mei, WANG Chunfeng. Light, scanning and transmission electron microscopical observation of the gill structure of[J]. Journal of Chinese Electron Microscopy Society, 2004(5): 553-559.
[52] 李艳菲, 狄桂兰, 王宁, 等. 鱼类黏液细胞研究进展[J]. 水产科学, 2020, 39(1): 143-150.
LI Yanfei, DI Guilan, WANG Ning, et al. Research advances on fish mucous cells[J]. Fisheries Science,2020, 39(1): 143-150.
Effect of salinity onbehavior and antioxidant enzyme activities and tissue structure of the gill and kidney
ZHANG Si-yan1, WANG Shao-jun2, YANG Jia-cheng1, ZHU Jian-ye1, TIAN Ying1, WANG Luo1, MAO Jun-xia1, WANG Xu-bo1, CHANG Ya-qing1, LU Zhi-chuang3, HAO Zhen-lin1
(1. Key Laboratory of Mariculture & Stock Enhancement in North China’s Sea, Ministry of Agriculture and Rural Affairs, Dalian Ocean University, Dalian 116000, China; 2. Shandong Fisheries Development and Resource Conservation Center, Jinan 250000, China; 3. Liaoning Ocean and Fisheries Science Research Institute, Dalian 116000, China)
The behavior, pathological damage, and antioxidant enzyme activities ofunder different salinity conditions (16, 20, 24, 28, 32, 36, 40, 44, and 48) were studied using different methods to clarify the physiological and ecological response and adaptive mechanism ofunder different salinities. The results showed thatexhibited a stress response, decreased feeding rate, and increased mortality under high and low salt stress. The total antioxidant capacity offirst increased and then decreased as salinity increased from 16 to 32. The activities of catalase (CAT) and superoxide dismutase (SOD) in gill first decreased and then increased. Kidney CAT and SOD activities first increased and then decreased. The tissue section results showed that the morphology of theolfactory apparatus and gill filaments changed significantly under high salt and low salt stress, and the number of mucus cells increased. These results show that salinity significantly affected the physiological and ecological responses of.adapted to the external salinity changes by changing behavior, regulating antioxidant enzyme activities, and changing tissue morphological structure.
; salinity; behavior; antioxidant enzyme activity; pathological damage
Oct. 20, 2021
S966.28
A
1000-3096(2022)10-0129-11
10.11759/hykx20211020004
2021-10-20;
2022-02-06
国家自然科学基金(42076101); 辽宁省自然资源厅海洋经济发展专项; 辽宁省教育厅面上项目(LJKZZZ0700); 大连市重点研发计划项目(2021YF16SN015)
[National Natural Science Foundation of China, No. 42076101; Program of Marine Economic Development of Liaoning Provincial Department of Natural Resources; General Program of Department of Education of Liaoning Province, No. LJKZZZ0700; Program of Dalian key R & D plan, No. 2021YF16SN015]
张思研(1998—), 女, 辽宁沈阳人, 硕士研究生, 研究方向: 水产, E-mail: 1173695993@qq.com; 郝振林(1980—),通信作者, 教授, E-mail: haozhenlin@dlou.edu.cn; 鹿志创(1982—), 通信作者, 硕士生导师, 研究员, E-mail: luzhichuang@hotmail.com
(本文编辑: 杨 悦)
