泥蚶快速生长季生化代谢物的变化特征
2022-11-25梅丽敏吴延华孙爱丽徐建峰周成旭
梅丽敏, 吴延华, 杨 玲, 孙爱丽, 徐建峰, 周成旭
泥蚶快速生长季生化代谢物的变化特征
梅丽敏1, 吴延华1, 杨 玲1, 孙爱丽2, 徐建峰3, 周成旭1
(1. 宁波大学 食品与药学学院, 浙江 宁波 315211; 2. 宁波大学 海洋学院, 浙江 宁波 315211; 3. 福建省连江县水产技术推广站, 福建 连江 350500)
研究了象山港泥蚶()快速生长季(2020年5—11月)糖原、游离氨基酸和脂肪酸等代谢物的组成、含量及其变化, 并对其营养特征变化进行了分析。结果显示, 7—9月为泥蚶快速生长期, 此期前后泥蚶生长缓慢。泥蚶肌肉糖原含量约5.6~11.1 mg/g; 牛磺酸、丙氨酸、谷氨酸和天冬氨酸是其主要的游离氨基酸; 不饱和脂肪酸含量丰富, 且主要为多不饱和脂肪酸。生长过程中几种生化代谢物含量变化显著: 5—7月间, 糖原含量显著降低、氨基酸和脂肪酸组成发生显著变化。7—9月间, 糖原含量、生糖和呈味氨基酸含量显著升高、饱和及单不饱和脂肪酸含量显著降低, 但脂肪酸组成结构无显著差异。11月, 氨基酸及多不饱和脂肪酸含量均显著升高。11月泥蚶具有高的营养品质。本研究为泥蚶养殖管理及水产品营养研究提供参考。
泥蚶(); 快速生长期; 糖原; 游离氨基酸; 脂肪酸
泥蚶()是我国重要的滩涂养殖贝类, 营养丰富, 且富含生物活性物质, 具有极高的食用、药用及经济价值[1-3]。糖原、脂肪酸和氨基酸等, 既是生物自身生理活动重要的调节物质, 也是水产食品品质和营养的指标物质。糖原在能量代谢中具有重要作用[4], 其含量高低反映了贝类生理状态、抗逆能力[5]。游离氨基酸在贝类中含量丰富, 参与维持细胞渗透压、内平衡等关键生理调节活动[6]。脂肪酸是脂类代谢产物, 在低温适应、渗透调节、繁殖发育等[7-8]方面起着关键作用。而水产食品营养、质量品质等, 与其此类代谢物的组成和含量均密切相关。糖原和游离氨基酸是水产品新鲜度、风味和营养等特征的重要评价指标[9]。脂肪酸组成和比例是脂质营养品质的评价标准。研究显示,-3与-6 多不饱和脂肪酸对人体有着重要的生理功能[10]。基于脂肪酸组成特征的脂质质量指数, 如PUFA/SFA (polyunsaturated fatty acid/ saturated fatty acids)、(-6)/(-3)、低/高胆固醇血症脂肪酸比例(hypocholesterolaemic/hypercholesterolaemic fatty acid ratio, H/H)、致动脉粥样硬化指数(index of atherogenicity, AI)和血栓形成指数(index of thrombogenicity, TI)等, 已被广泛用于评估人类食品的营养价值。
养殖生物生化代谢过程受到生物自身内因以及环境条件外因的影响, 如生长、繁殖、季节、温度、水质以及食物的可获得性等[11-13]。双壳贝类的生长和繁殖具有阶段性。自然滩涂养殖的泥蚶, 不仅生殖周期具有显著的节律性, 其个体生长速率也有差异, 表现为第一年生长相对快速、第二年及以后则生长速率下降。快速生长期也是养殖管理的重要时期。对泥蚶研究涵盖了多个领域, 例如, 关于不同区域营养成分的比较[14-15]、生物学特性和生态习性[16-18]、贝藻饵料[19-20]等众多方面。Nguyen等[21]研究了韩国南部海岸泥蚶在不同季节的营养组成特征。不同养殖物种在不同季节的营养组成变化对养殖管理和水产品营养研究及加工具有重要的参考价值。
本研究于2020年5—11月间, 跟踪采集了象山港滩涂区养殖的泥蚶由幼贝进入成贝快速生长前后的样品, 定量分析了糖原、游离氨基酸、脂肪酸等代谢物的组成、含量及变化特征, 并基于营养品质指标, 解析了泥蚶在显著生长季中营养品质变化特征, 拟为泥蚶养殖管理、水产品营养和加工研究提供参考。
1 材料与方法
1.1 泥蚶样品采集、处理和环境因子调查
2019年9月, 在象山港塔头旺高潮蓄水塘中, 投泥蚶苗(规格: 壳长9.1 mm、壳宽6.8 mm、壳高5.4 mm)。2020年5—11月间泥蚶显著生长阶段, 于5、7、9、11月采集泥蚶。及时运回实验室, 置冰上解剖。随机取30个泥蚶样品, 游标卡尺测量壳长、壳宽、壳高, 解剖全部软组织用于组织干重分析。取足量消化腺和肌肉样品用于糖原含量分析; 取足量肌肉样品分别用于游离氨基酸(除色氨酸)、脂肪酸含量分析。肌肉样品指除去消化腺和生殖腺的其他软组织。解剖样于−80 ℃冰箱保存备测。现场采集蓄水塘中3个位点水样, 水质分析仪(YSI, Professional Plus, USA)检测水温、盐度及叶绿素(Chl)含量。
1.2 糖原分析方法
以蒽酮法分析糖原含量[22]。称取肌肉和消化腺鲜样各100.0 mg, 加入 8 mL 5%三氯乙酸, 匀浆 1 min, 3 000 r/min 离心匀浆液 15 min, 取 1 mL上清液于10 mL离心管中, 加入 4 mL 95%乙醇, 置于37~40 ℃ 水浴3 h。3 000 r/min 离心 15 min, 弃上清液, 加入1 mL 蒸馏水溶解沉淀的糖原; 配制空白管与标准管溶液, 各加入5 mL蒽酮硫酸试剂, 混匀后沸水浴中反应15 min(反应液呈蓝绿色), 取出后迅速于流水下冷却以停止反应。酶标仪检测吸光度(620), 根据标准曲线计算糖原含量。
1.3 游离氨基酸分析
准确称量1.0 g泥蚶肌肉样品, 加入10 mL纯水, 充分均质后, 超声破碎10~15 min, 4 ℃冷冻离心(12 000 r/min) 10 min, 取上清液超滤(截留分子量 3 kDa) (4 000r/min, 30 min, 4 ℃), 收集滤液冷冻干燥。取冷冻干燥后粉末, 以1 mL盐酸(0.02 moL/L)复溶后, 以微量全自动氨基酸分析仪(LA8080, 日本日立公司)分析测定。采用铝离子交换树脂柱(4.6 mm× 60.0 mm)进行样品分离; 氨基酸标准品为日本日立公司标准氨基酸混合液(B型和AN-Ⅱ型); 反应柱为热传导膜不锈钢柱4.6 mm ×(42.5 mm+42.5 mm); 交换柱柱温50 ℃, 反应柱柱温135 ℃; 缓冲液流速0.4 mL/min; 茚三酮流速0.35 mL/min。检测波长570 nm, 检测时间150 min。
1.4 脂肪酸分析
将贝类肌肉样品经冷冻干燥后, 研钵碾碎成粉末。准确称取100.0 mg肌肉干粉, 各5个平行。总脂提取在Blight-Dyer法上稍作改进[23]。提取总脂后加入100 μL浓度为1 μg/μL的C19: 0脂肪酸内标、2 mL 5%~6% 氢氧化钾甲醇水溶液(V/V=4︰1), 60 ℃水浴皂化2 h。冷却后以6 mL氯仿正己烷混合液(V/V=1︰4)分3次进行萃取, 3 000 r/min离心, 取上清液。旋转蒸发干燥后, 加入0.5 mL三氟化硼甲醇溶液, 60 ℃水浴1 h。冷却后用6 mL分析纯级正己烷分3次萃取, 3 000 r/min离心, 取上清液。向上清液中加入3 g无水硫酸钠, 静置3 h后取上清液, 旋转蒸发干燥后, 用1 mL色谱纯级正己烷定容, 用于气相色谱-质谱联用(gas chromatography-mass spectrometry, GC-MS)上机分析。
GC条件: 不分流模式进样。柱温箱起始温度140 ℃保持5 min, 以4 ℃/min, 升温至240 ℃保持30 min。进样口温度250 ℃。载气用高纯氦气, 流速为2.25 mL/min。MS条件: 采用EI 离子源分析, 电子能量为70 eV。离子源温度为230 ℃, 灯丝发射电流为200 μA。扫描质量范围为40~600 m/z。扫描模式采用Scan模式。
基于各组分质谱, 初步确定分子质量, 结合离子碎片质量谱图, 通过NIST库和WILEY库进行检索[24], 保留相似度大于80%的组分, 参考文献进行鉴定。同时用37种脂肪酸甲酯标准品的保留时间来进行辅助鉴定。用面积归一法分别计算出各种脂肪酸的百分含量, 结果保留两位小数。基于内标C19: 0脂肪酸的初始浓度、峰面积和各待测组分的峰面积, 计算测脂肪酸组分的绝对含量, 单位为μg/mg, 响应因子按1计算。
1.5 数据统计分析
数据计算作图采样EXCEL 2019, 结果以平均值±标准差(X±SD) 表示。采用IBM-SPSS26软件进行单因素方差分析(One-way ANOVA), 统计学显著性设置0.05。采用Simca-P软件进行主成分分析(PCA)。
第一,控制生物制品,加大打击力度。在实际生活中,有关部门应该加大抽检兽药质量力度和监管力度,执行查处兽药生产使用、经营等违法案例,加强动物防疫疫苗和试剂的订购,确保有效供应,质量符合标准,并且需要和生物安全的环节,相关部门领导应加大管理力度,有效督促检查实验室活动,及时排除隐患,确保安全,对于突出问题应进行整改。各级部门应执行控制生物制品生产和使用,防止疫苗上市出售,有违反规定者进行处罚。
2 结果
2.1 泥蚶生长过程及水质、叶绿素a变化
泥蚶生长过程如图1a所示。总体而言, 5—7月为快速生长前期、7—9月为快速生长期、9—11月为快速生长后期。软组织干重在快速生长前后期无显著变化, 但7—9月相对增长了400%(由0.01 g增至0.05 g)。个体大小呈现持续增长的特征, 但5—7月或9—11月间无显著差异(0.05)。
水温变化范围22.35~34.32 ℃, 7月最高, 11月最低(图1b)。盐度变化范围24.03~27.92, 9月相对较高。Chl平均为1.61 µg/L, 9月相对最高(2.15 µg/L), 11月下降至最低(1.24 µg/L), 5—9月间无显著差异(0.05)。
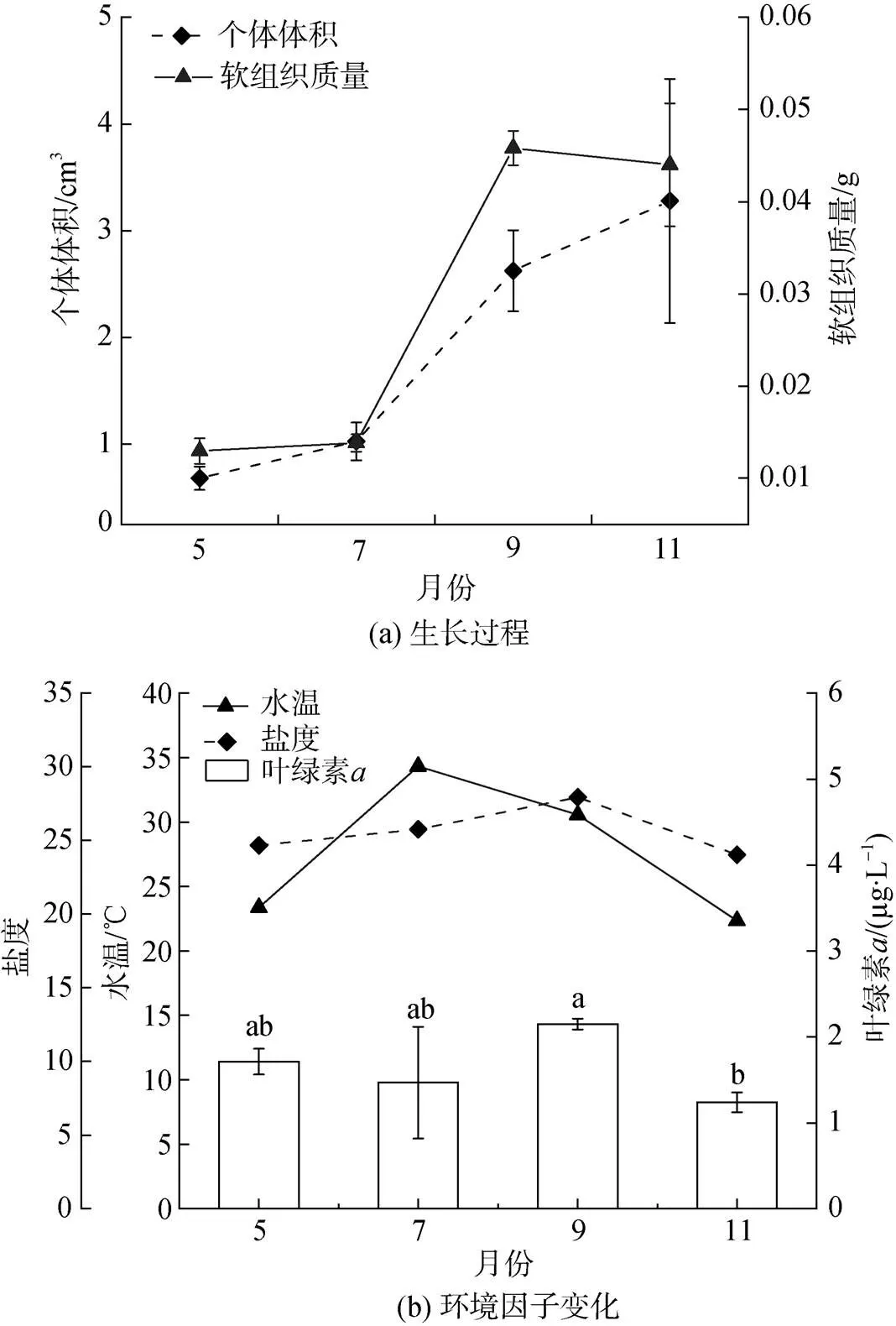
图1 泥蚶生长过程(a)及水温、盐度和叶绿素a(b)变化
2.2 糖原含量的变化
泥蚶消化腺和肌肉的糖原含量不同, 且在生长过程中发生显著变化(图2)。快速生长前(5月)后(9—11月), 肌肉的糖原含量显著大于消化腺(0.05)。5月, 肌肉糖原含量最高[(11.14±0.24) mg/g], 而进入7月时, 其含量显著降低(0.05), 且7月的肌肉糖原含量为整个生长季最低[(5.56±0.29) mg/g]。9月, 糖原含量显著升高, 至11月无显著变化。5—7月间, 消化腺糖原含量亦显著降低(由5.98±0.27降低至5.13±0.26 mg/g), 但7—11月间则无显著变化(0.05)。

图2 不同生长月泥蚶消化腺及肌肉糖原含量变化
2.3 游离氨基酸组成及含量变化
在所有样本中, 共检测了19种游离氨基酸(表1)。相对于人类食用水产品而言, 对氨基酸进行了必需氨基酸、呈味氨基酸分类解析。
其中7种必需氨基酸, 分别是赖氨酸、亮氨酸、苯丙氨酸、缬氨酸、苏氨酸、异亮氨酸、蛋氨酸, 其含量为223.91~322.06 µg/g, 占12.30%~15.27%。赖氨酸含量最高, 5—11月含量为52.73~83.30 µg/g, 11月最高。蛋氨酸含量最低, 5—11月其含量为15.34~ 26.15 µg/g。7—9月, 必需氨基酸含量均呈升高趋势, 但仅蛋氨酸含量具有统计学显著性(0.05); 11月, 总必需氨基酸达最大含量(0.05)(图3a; 表1)。12种生糖氨基酸中(表1), 天冬氨酸含量相对最高, 平均为[(199.82±16.56) µg/g]。精氨酸在11月样品中未检出, 半胱氨酸在9月未检出。
总生糖氨基酸含量为609.64~864.868 µg/g, 占36.88%~41.60%。其含量在7—9月显著升高(0.05), 但相对丰度无显著变化(0.05)。9—11月, 总生糖氨基酸亦呈升高趋势, 但无统计学显著性(0.05) (图3a; 表1)。

图3 不同生长月泥蚶肌肉各类别游离氨基酸含量变化(a)及氨基酸组成的PCA分析(b)
Fig. 3 Variations of different group of free amino acids in muscle of Tegillarca granosa (a) and PCA analysis of the composition of the free amino acids (b) during the growth period
5种呈味氨基酸含量总计382.18~642.67 µg/g, 占24.84%~30.05%。呈味氨基酸含量在7—9月显著升高(0.05), 11月显著升高(0.05)至最大值(图3a; 表1)。其相对丰度在7—9月无显著变化(0.05), 但进入11月, 亦显著升高(0.05)。
其他游离氨基酸中, 牛磺酸在所有游离氨基酸中含量最高, 占41.71%~45.52%, 达(665.55±53.86)~ (1 030.67±43.17) µg/g。其变化特征显示, 5—7月间, 含量显著降低, 7—9月间含量显著升高, 至11月, 显著升高至最大含量(0.05)。但相对丰度呈现与含量显著相反的变化趋势。鸟氨酸含量相对最低, 为(12.86±1.27)~(23.13±0.17) µg/g, 占0.88%~1.28%。7—9月间, β-丙氨酸显著增加(0.05), 酪氨酸和鸟氨酸无显著变化(0.05)。
2.4 脂肪酸组成特征、含量及脂质营养指数变化
在所有样本中, 共检出18种脂肪酸(表2), 包括5种饱和脂肪酸(SFA), 6种单不饱和脂肪酸(MUFA)以及7种多不饱和脂肪酸(PUFA)。PUFA占比最高(42.78%~50.12%), SFA次之(32.96%~35.74%), MUFA占比最低(16.91%~22.14%)。
SFA含量为3.72~4.46 mg/g, 由C14: 0、C16: 0、C18: 0和C20: 0组成。占比较高的主要是C16: 0 (15.37%~17.21%)、C18: 0 (12.23%~15.06%)。SFA含量在7—9月间显著下降(<0.05), 11月恢复至5—7月间水平(图4a)。SFA的相对丰度在5—11月间均无显著变化(0.05)。
MUFA含量为2.19~2.8 mg/g, 占16.91%~22.14%。C20: 1(-13)相对丰度最高, 占7.04%~11.53%。MUFA含量在7—9月间显著降低(0.05)(图4a), 但相对丰度无显著差异(0.05)。
PUFA主要由-3和-6组成, 含量为5.27~ 6.51 mg/g, 占42.78%~50.12%。PUFA含量在5—9月间无显著差异, 但11月显著高于其他各月(图4a)。其相对丰度在7月时显著最低(0.05), 11月显著最高(0.05)。C22: 2(-6)是含量最高的PUFA(平均2.34±0.33 mg/g), 5—11月间无显著变化。ARA、EPA、DHA的含量和相对丰度均在11月达到最大(0.05)。
对脂肪酸组成的PCA分析显示(图4b), 7—9月脂肪酸聚为一组, 5月和11月分别单独聚类。显示7月和9月脂肪酸的组成具有相似的组成结构, 与5月和11月差异较大。5月与11月脂肪酸组成结构差异大。
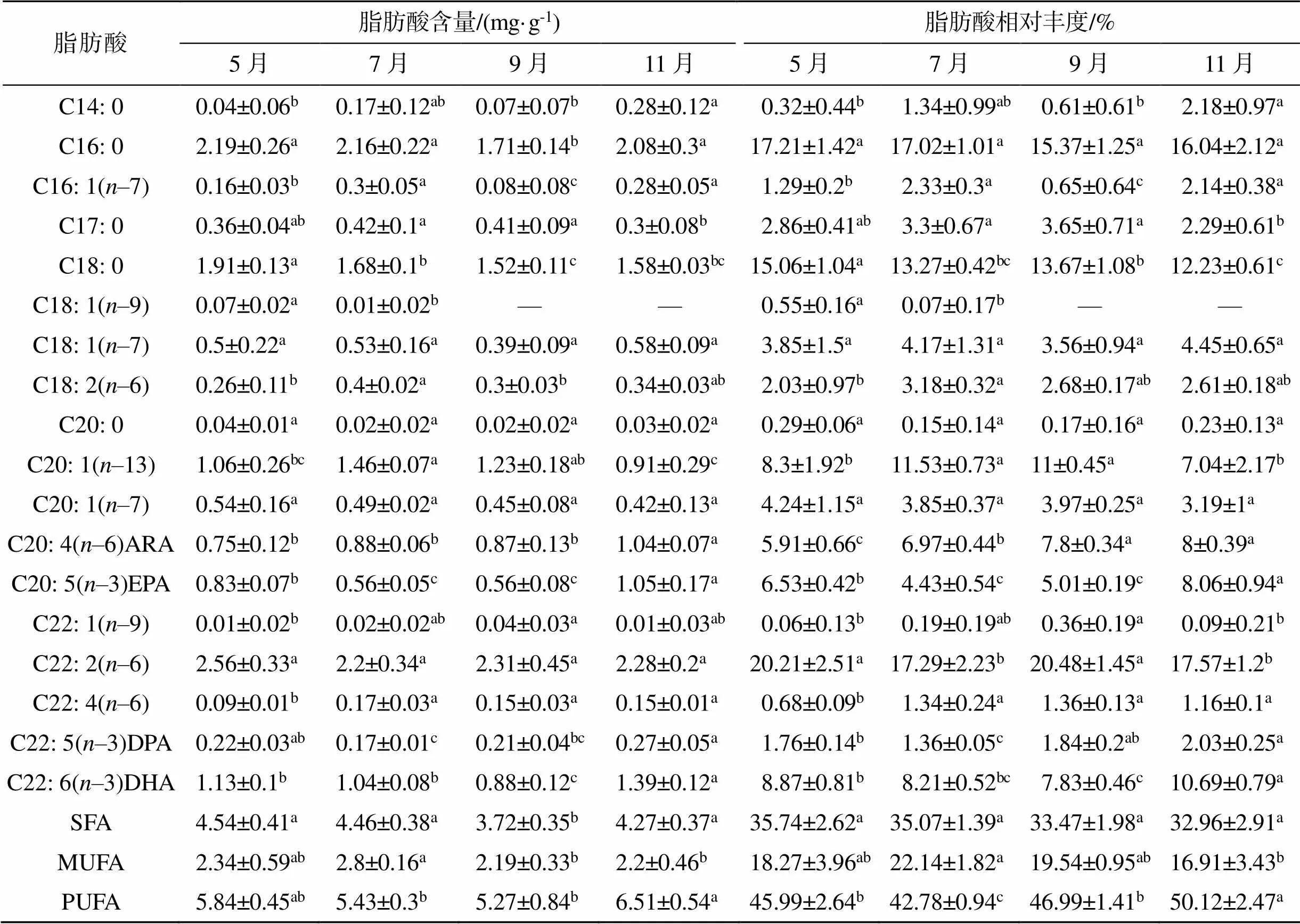
表2 不同生长月中泥蚶肌肉脂肪酸的组成和含量变化
注: 表中数据以平均值±标准差表示, 每组5个平行; 不同字母代表数据之间具有显著性差异(0.05); —表示未检测到; SFA: 饱和脂肪酸; MUFA: 单不饱和脂肪酸; PUFA: 多不饱和脂肪酸。
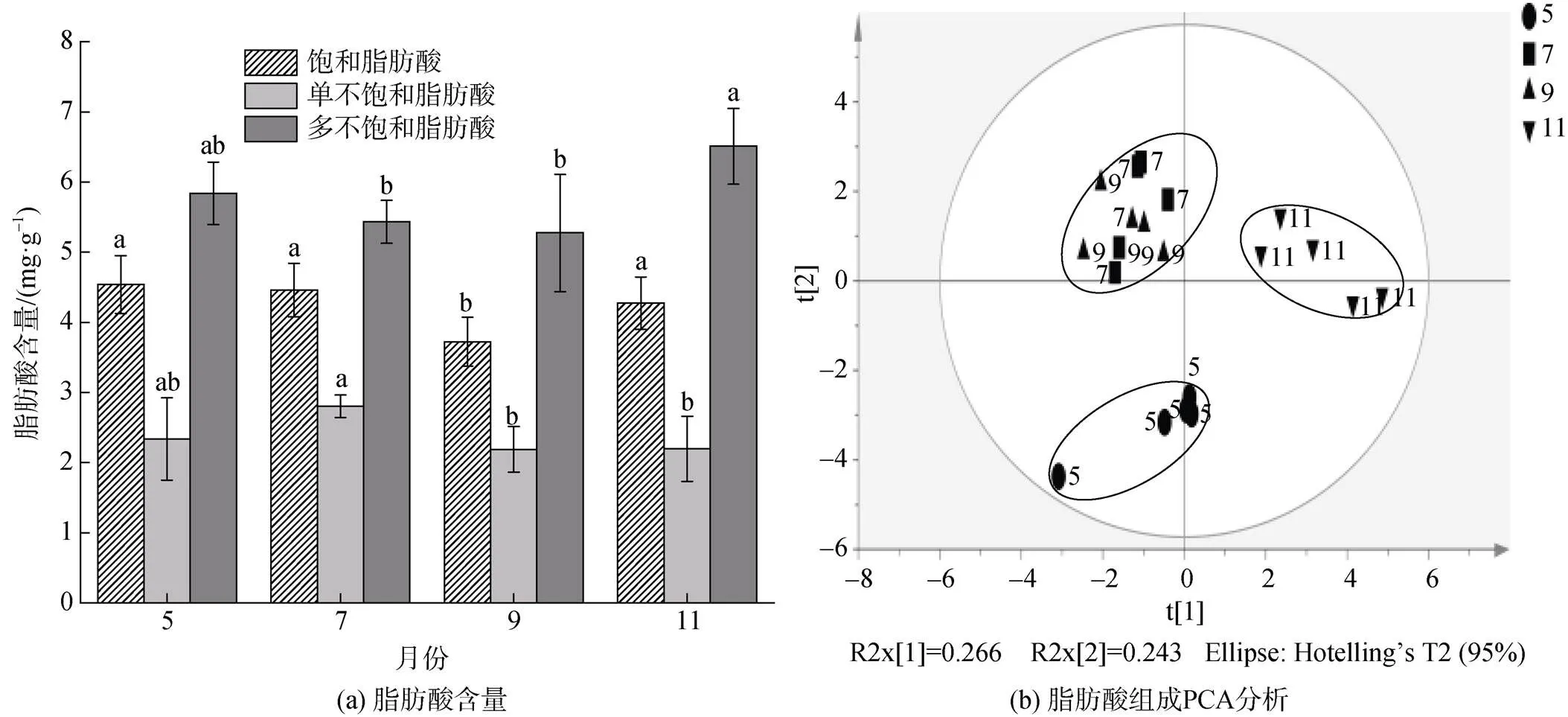
图4 不同生长月泥蚶肌肉各类别脂肪酸含量变化(a)及脂肪酸组成PCA分析(b)
脂肪酸特征指标及脂质营养指数特征显示(表3), 总脂肪酸(TFA)7—9月间显著降低(0.05), 11月恢复至与5月相当的水平。11月, ∑(-3)较前期各月具有显著最高值(20.78%,0.05)。EPA/DHA在5—7月间显著降低(0.05), 7—11月间呈逐渐升高特点(0.05)。∑(-6)/∑(-3)在5月和11月分别为1.69±0.26和1.42± 0.1, 较7月和9月(2.07±0.25, 2.2±0.12)显著低(0.05), 7—9月间无显著差异(0.05), 11月显著最低(0.05)。PUFA/SFA在9—11月间较5—7月间有显著增加(0.05), 11月达最大值。
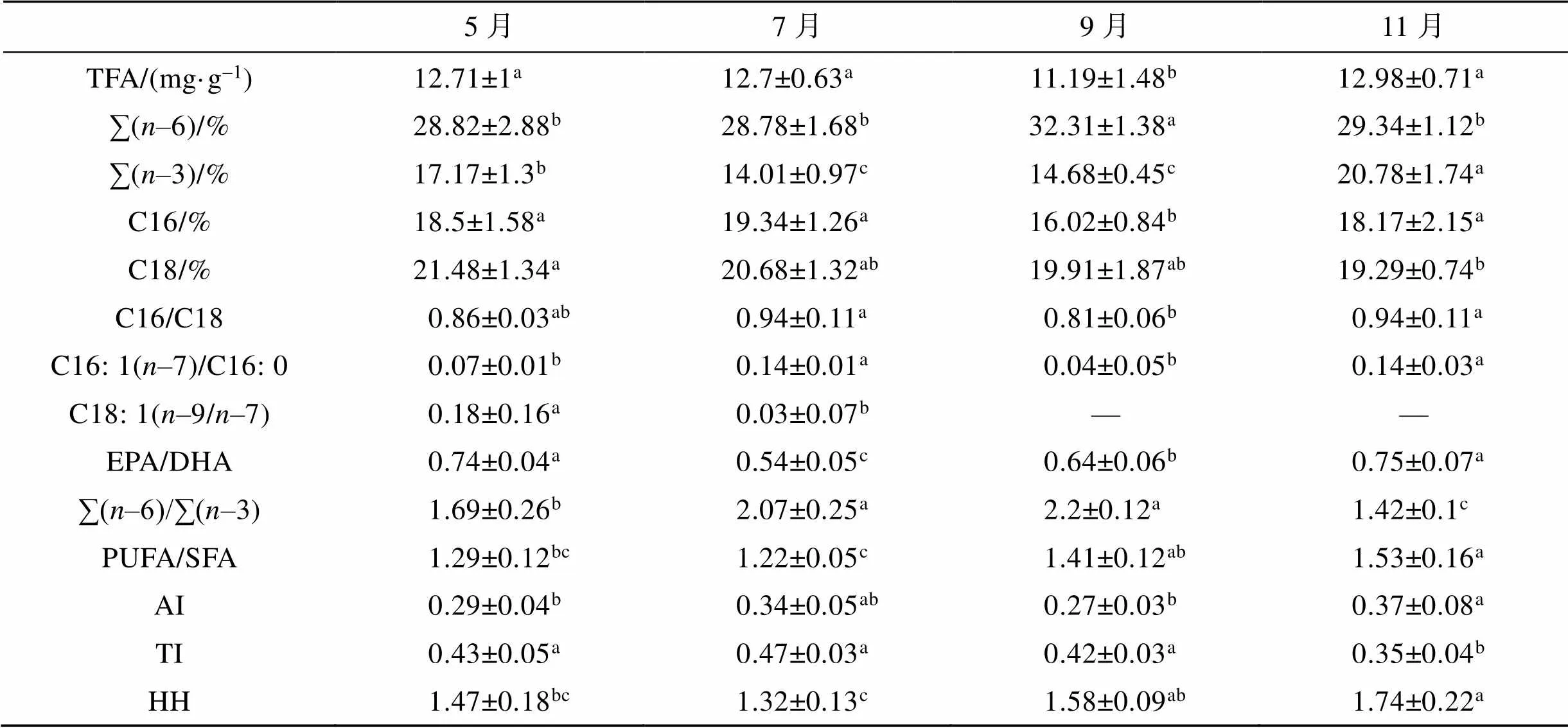
表3 不同生长月中泥蚶肌肉脂肪酸特征指标及脂质营养指数
5—11月间, AI平均值0.32±0.05, 变化范围在(0.27±0.03)~(0.37±0.08); TI平均值0.42±0.04, 变化范围在(0.35±0.04)~(0.47±0.03)。AI在11月显著升高(0.05), TI在11月显著最低(0.05); HH平均值1.53±0.16, 变化范围在(1.32±0.13)~(1.74±0.22)。HH在7—11月显著升高, 11月达最大值。
3 讨论
糖原在能量代谢中起到关键的作用。高温使代谢加快, 能量消耗增加, 糖原含量降低; 低温时, 新陈代谢速度降低, 能量消耗减少[25-26]。5月环境温度较低, 生长速度较为缓慢, 无需消耗大量的能量, 肌肉和消化腺糖原含量均较高。7月, 水温升高, 代谢加快, 能量消耗增加, 糖原含量显著下降。5—7月, 泥蚶进入生殖成熟期, 生殖细胞成熟需要消耗大量储存的糖原, 也可导致糖原显著降低[21, 25]。但过低的糖原是导致贝类死亡的重要因素。Patrick等对长牡蛎()的研究显示[5], 进入夏季时, 生产糖原的代谢不足, 牡蛎糖原降低, 可能导致死亡率高发。因此糖原显著降低的时期也是贝类生理脆弱期。泥蚶消化腺糖原含量从5月降低以后, 7—11月间, 维持相对稳定。双壳类消化腺类似于脊椎动物的肝脏, 起到脱毒解毒的重要作用[27-28], 其糖原含量的相对稳定可能保障了贝类维持正常生理活动需求。7—9月快速生长期间, 生糖氨基酸含量显著升高。生糖氨基酸可通过糖异生作用转化为葡萄糖[27], 可能与此期糖原含量升高相关。
海洋软体动物可以利用高浓度的氨基酸来调节细胞渗透压[29]。丙氨酸、甘氨酸、精氨酸、天冬氨酸、谷氨酸和牛磺酸等游离氨基酸, 在贝类渗透调节中尤为重要[30]。本研究显示, 此类氨基酸均是相对高含量的, 且月际变化相对显著。从几类氨基酸总体含量及相对丰度的月际变化来看, 氨基酸代谢在泥蚶生长过程中是活跃调控的。
游离氨基酸在11月显著增加, 可能显示了蛋白合成降低或者蛋白降解增加。11月水温和Chl的含量显著下降, 低温和食物相对减少, 可能此期泥蚶生理代谢更多地适应于低温和食物限制。相对于其他显著变化的氨基酸而言, 苯丙氨酸在各生长月间均无显著差异。苯丙氨酸是能量代谢关键酶丙酮酸激酶的抑制剂, 丙酮酸激酶对苯丙氨酸的浓度变化敏感[31]。苯丙氨酸维持相对稳定的含量, 可能说明该氨基酸没有参与泥蚶的能量代谢调控。
贝类体内脂肪酸含量和组成与物种、饵料、温度、盐度等因素有关。虽然在同一地点采集的同一种贝类基本具有相似的脂肪酸组成[32], 但本研究显示, 泥蚶生长过程中, 脂肪酸组成和含量发生了变化。不同于氨基酸在不同月际间均有明显变化, 脂肪酸组成在7—9月间相似, 显示脂肪酸的变化与季节关系更为密切。11月温度显著降低时, 多不饱和脂肪酸含量显著增加。低温时细胞膜流动性减弱, 多不饱和脂肪酸的上升可提高细胞膜的流动性, 减少泥蚶组织损伤的程度。
泥蚶营养品质随着几种生化代谢物的月际变化而差异显著。11月必需氨基酸含量最高, 且呈味氨基酸和糖原含量也升高, 显示此期泥蚶不仅营养价值相对较高, 且较鲜美以及口味甘甜。Cordain等人研究认为[33], ∑(-6)/∑(-3)值应当低于4, 可降低一些疾病的发病率。泥蚶在各生长月中, 该值均低于4。在关于脂质营养指标的各研究中提出[34-37], 食物中PUFA/SFA的值应当高于0.4, 更有利于降低人体患某些慢性疾病的风险; AI和TI低于1时, 对于人体防治相关疾病是有重要意义的; 而H/H越高则越有利于人体降低血液中的胆固醇含量。本研究发现, 泥蚶在各生长月中, PUFA/SFA值均超过0.4, 且在11月比值最高, 说明11月的泥蚶更有利于人体的健康; 泥蚶的AI和TI指数在各生长月中均低于1; HH在7—11月显著升高, 在11月含量最高。由此可见, 泥蚶在各生长月中均符合利于人类健康的营养品质, 11月时, 其营养品质最高, 此期以后收获为最佳状态。
4 结论
泥蚶快速生长季中, 生化代谢物的组成和含量具有显著的变化。游离氨基酸处于活跃代谢调控状态, 脂肪酸组成与季节变化关系更为密切。泥蚶进入夏季快速生长时, 糖原和多种氨基酸呈现显著降低的特征, 为泥蚶较不稳定的生理状态期。不同生长阶段的泥蚶均有较高的营养指数, 快速生长后, 糖原、游离氨基酸和多不饱和脂肪酸含量均显著升高, 并在11月呈现最高值, 说明此期泥蚶具有最高的营养品质。
[1] LI G Y, LIU J Z, CHEN S G, et al.extract Haishengsu inhibits the expression of-glycoprotein and induces apoptosis in drug-resistant K562/ ADM cells[J]. Pharmaceutical Biology, 2010, 48(5): 529-533.
[2] LV S, GAO J, LIU T, et al. Purification and partial characterization of a new antitumor protein from[J]. Marine Drugs, 2015, 13(3): 1466-1480.
[3] YANG S, DONG Y, AWEYA J J, et al. Antimicrobial activity and acting mechanism ofhemoglobin-derived peptide (TGH1) against[J]. Microbial Pathogenesis, 2020, 147: 104302.
[4] BAYNE B L. Metabolic expenditure[M]. Amsterdam: elsevier, 2017, 41: 331-415.
[5] PATRICK S, FAURY N, GOULLETQUER P. Seasonal changes in carbohydrate metabolism and its relationship with summer mortality of Pacific oyste(Thunberg) in Marennes-Oléron bay (France)[J]. Aquaculture, 2006, 252(2/4): 328-338.
[6] TANIMOTO S, KAWAKAMI K, MORIMOTO S. Changes in the free amino acid content of the shucked oysterstored in salt water at 3℃[J]. Fisheries and Aquatic Science, 2013, 16(2): 63-69.
[7] FARKAS T, NEVENZEL J C. Temperature acclimation in the crayfish: Effects on phospholipid fatty acids[J]. Lipids, 1981, 16(5): 341-346.
[8] FOULADI-NASHTA A A, GUTIERREZ C G, GONGJ G, et al. Impact of dietary fatty acids on oocyte quality and development in lactating dairy cows[J]. Biology of Reproduction, 2007, 77(1): 9-17.
[9] 张苏平, 邱伟强, 卢祺, 等. 全自动氨基酸分析仪法测定4种贝类肌肉中谷胱甘肽和游离氨基酸含量[J]. 食品科学, 2017, 38(4): 170-176.
ZHANG Supin, QIU Weiqiang, LU Qi, et al. Determination of glutathione and free amino acids in muscles of four shellfish species by automatic amino acid analyzer[J].Food Science, 2017, 38(4): 170-176.
[10] RYAN A S, ASTWOOD J D, GAUTIER S, et al. Effects of long-chain polyunsaturated fatty acid supplementation on neurodevelopment in childhood: a review of human studies[J]. Prostaglandins, Leukotrienes and Essential Fatty Acids, 2010, 82(4/6): 305-314.
[11] DRIDI S, ROMDHANE M S, ELCAFSI M. Seasonal variation in weight and biochemical composition of the Pacific oyster,in relation to the gametogenic cycle and environmental conditions of the Bizert lagoon, Tunisia[J]. Aquaculture, 2007, 263(1/4): 238-248.
[12] MARIN M G, MOSCHINO V, DEPPIERI M, et al. Variations in gross biochemical composition, energy value and condition index offrom the Lagoon of Venice[J]. Aquaculture, 2003, 219(1/4): 859- 871.
[13] 喻国洪, 王扬才, 金中文. 两次卵巢发育成熟拟穴青蟹卵巢与肝胰腺营养成分比较分析[J]. 海洋科学, 2021, 45(9): 12-20.
YU Guohong, WANG Yangcai, JIN Zhongwen.Comparative analysis of nutrient composition of the ovaries and hepatopancreas in the mud crabat two stages of ovary maturation[J]. Marine Sciences, 2021, 45(9): 12-20.
[14] 张永普, 贾守菊, 应雪萍. 不同种群泥蚶肉营养成分的比较研究[J]. 海洋湖沼通报, 2002(2): 33-38.
ZHANG Yongpu, JIA Shouju, YING Xueping.A comparative study on the nutritive composition and contents in the meat ofin varied populations[J].Transactions of Oceanology and Limnology, 2002(2): 33-38.
[15] 詹堃, 李太武, 苏秀榕. 阳江乐清两地泥蚶肌肉组织营养成分研究[J]. 水产科学, 2007, 26(12): 665-667.
ZHAN Kun, LI Taiwu, SU Xiurong.Fatty acids and amino acids in muscular tissue of mud clamlinnaeus from two different regions[J].Fisheries Science, 2007, 26(12): 665-667.
[16] 焦海峰, 严巧娜, 郑丹, 等. 温度和盐度对埋栖性双壳类泥蚶()呼吸、排泄的影响[J]. 海洋与湖沼, 2015, 46(6): 1333-1338.
JIAO Haifeng, YAN Qiaona, ZHENG Dan, et al.Effects of temperature and salinity on oxygen consumption and ammonia excretion of muddy shellfish[J]. Oceanologia et Limnologia Sinica, 2015, 46(6): 1333-1338.
[17] 吴洪喜, 柴雪良. 浙江乐清湾泥蚶的繁殖习性和生长特性[J]. 动物学杂志, 2004, 39(3): 47-50.
WU Hongxi, CHAI Xueliang. The reproductive habit and growth character of bloody clamin Yueqing Bay[J]. Chinese Journal of Zoology, 2004, 39(3): 47-50.
[18] 李敏, 郑伊诺, 许凯伦, 等. 盐度胁迫对泥蚶存活率及3种酶活力的影响[J]. 浙江农业科学, 2018, 59(4): 650-653, 657.
LI Min, ZHENG Yinuo, XU Kailun, et al.Effects of salinity stress on survival rate and three enzyme activities of[J]. Journal of Zhejiang Agricultural Sciences, 2018, 59(4): 650-653, 657.
[19] GENG S, ZHOU C, CHEN W, et al. Fatty acid and sterol composition reveal food selectivity of juvenile ark shellLinnaeus after feeding with mix microalgae[J]. Aquaculture, 2016, 455: 109-117.
[20] XU J, ZHOU H, YAN H, et al. Effect of unialgal diets on the composition of fatty acids and sterols in juvenile ark shellLinnaeus[J]. Journal of Agricultural and Food Chemistry, 2012, 60(15): 3973-3980.
[21] NGUYEN T T, CHOI Y J, ROHMAH Z, et al. Seasonal variations of nutritional components in cockles () processed from the Southern Coast of Korea[J]. Cogent Food and Agriculture, 2017, 3(1): 1360102.
[22] CARROLL N V, LONGLEY R W, ROE J H. The determination of glycogen in Liver and muscle by use of anthrone reagent[J]. The Journal of Biological Chemistry, 1956, 220(2): 583-593.
[23] BLIGH E G, DYER W J. A rapid method lipid extraction and purification[J]. Canadian Journal of Biochemistry and Physiology, 1959, 37(8): 911-917.
[24] 徐继林, 严小军, 周成旭, 等. 甾醇在海洋微藻中的分布研究[J]. 海洋学报, 2007, 29(6): 93-100.
XU Jilin, YAN Xiaojun, ZHOU Chengxu, et al.Studies on sterol distribution in marine microalgae[J]. Acta Oceanologica Sinica, 2007, 29(6): 93-100.
[25] ANDRADE J, CORDEIRO N, MONTRESOR L C, et al. Effect of temperature on behavior, glycogen content, and mortality in(Dunker, 1857) (Bivalvia: Mytilidae)[J]. Journal of Limnology, 2017, 77(2): 189-198.
[26] CAREY C S, JONES J W, HALLERMAN E M, et al. Determining optimum temperature for growth and survival of laboratory-propagated juvenile freshwater mussels[J]. North American Journal of Aquaculture, 2013, 75(4): 532-542.
[27] FAGGIO C, TSARPAL V, DAILIANIS S. Mussel digestive gland as a model tissue for assessing xenobiotics: An overview[J]. Science of the Total Environment, 2018, 636: 220-229.
[28] LOUIS F, ROCHER B, BARJHOUX I, et al. Seasonal monitoring of cellular energy metabolism in a sentinel species,(bivalve): Effect of global change?[J]. Science of the Total Environment, 2020, 725: 138450.
[29] JIE L, SHI Y, WANG S, et al. NMR-based metabolomic analysis ofexposed to thermal and hypoxic stresses[J]. Science of the Total Environment, 2016, 545: 280-288.
[30] KUBE S, SOKOLOWSKI A, JANSEN J M, et al. Seasonal variability of free amino acids in two marine bivalves,andspp., in relation to environmental and physiological factors[J]. Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology, 2007, 147(4): 1015-1027.
[31] MUSTAFA T, HOCHACHKA P W. Catalytic and Regulatory Properties of Pyruvate Kinases in Tissues of a Marine Bivalve[J]. Journal of Biological Chemistry, 1971, 246(10): 3196-3203.
[32] 李海英, 徐继林, 严小军. 16种贝类脂肪酸和甾醇组成分析[J]. 宁波大学学报(理工版), 2009, 22(1): 48-54.
LI Haiying, XU Jilin, YAN Xiaojun.Analysis of Fatty Acid and Sterol Composition in 16 Shellfishes[J].Journal of Ningbo University (Natural Science and Engineering Edition), 2009, 22(1): 48-54.
[33] CORDAIN L, EATON S B, SEBASTIAN A, et al. Origins and evolution of the Western diet: health implications for the 21st century[J]. The American Journal of Clinical Nutrition, 2005, 81(2): 341-354.
[34] SIMOPOULOS A P. The importance of the ratio of omega-6/omega-3 essential fatty acids[J]. Biomedicine and Pharmacother, 2002, 56(8): 365-379.
[35] 陈蝶玲, 黄巍峰, 郑晓辉, 等. N3系多不饱和脂肪酸膳食参考摄入量的研究进展[J].食品工业科技, 2015, 36(11): 378-388.
CHEN Dieling, HUANG Weifeng, ZHENG Xiaohui, et al. Research status of the dietary reference intakes of N3 polyunsaturated fatty acids[J].Science and Technology of Food Industry, 2015, 36(11): 378-388.
[36] SANTOS-SILVA J, BESSA R J B, SANTOS-SILVA F. Effect of genotype, feeding system and slaughter weight on the quality of light lambs: II. Fatty acid composition of meat[J]. Livestock Production Science, 2002, 77(2/3): 187-194.
[37] PLEADIN J, LEŠIĆ T, KREŠIĆ G, et al. Nutritional quality of different fish species farmed in the Adriatic Sea[J]. Italian Journal of Food Sciences, 2017, 29(3): 537-549.
Variations of biochemical metabolites induring rapid growth period
MEI Li-min1, WU Yan-hua1, YANG Ling1, SUN Ai-li2, XU Jian-feng3, ZHOU Cheng-xu1
(1. School of Food and Pharmaceutical Sciences, Ningbo University, Ningbo 315211, China; 2. School of Marine Sciences, Ningbo University, Ningbo 315211, China; 3. Aquaculture Technique Popularization Station of Lianjiang, Lianjiang 350500, China)
In the present study, blood clamgrown in the intertidal of Xiangshan Bay were sampled during a rapid growth period (May-November, 2020), and the variations of the content and composition of the metabolites, such as glycogen, free amino acids and fatty acids, and consequently the nutritional qualities, were studied. The results showed that, the rapid growth period was from July to September, before or after that, the growth rate was low. Glycogen in muscle was 5.6-11.1 mg/g. Taurine, alanine, glutamic acid, aspartic acid were the most abundant amino acids. Unsaturated fatty acids were abundant and PUFA was the dominant. The biochemical metabolites variation significantly during the growth period. In July, the content of glycogen decreased significantly compared to that in May. Free amino acids and fatty acids significantly changed as well during this period. From July to September, the contents of glycogen, amino acids that correlated to glycogen and seafood taste increased significantly, while SFA and MUFA decreased significantly. However, the composition of fatty acids was not significantly different between the two months. In November, the contents of amino acids and PUFA increased significantly. The content of total sterol did not change significantly. Relatively the highest nutritional benefits were achieved in November. This study applied the references in aquaculture management and food nutrition researches of blood clam.
; fast growing period; glycogen; free amino acids; fatty acids
Jan. 18, 2022
S968.3
A
1000-3096(2022)10-0094-10
10.11759/hykx20220118004
2022-01-18;
2022-08-03
国家重点研发计划 (2018YFD0900702); 科技基础资源调查专项 (2018FY100206)
[National Key Research and Development Plan, No. 2018YFD0900702; Special Investigation on Basic Resources of Science and Technology, No. 2018FY100206]
梅丽敏(1995—), 女, 安徽合肥人, 硕士研究生, 主要从事水产品品质构建的环境条件研究, E-mail: mlm096460@163.com; 周成旭(1968—),通信作者, E-mail: zhouchengxu@nbu.edu.cn
(本文编辑: 杨 悦)
