香蕉种质资源叶片表型性状多样性分析
2022-11-25赵丽娟只佳增张建春周劲松刘学敏张荣琴
赵丽娟,只佳增,张建春,杜 浩,周劲松,刘学敏,张荣琴
(云南省红河热带农业科学研究所,云南河口 661300)
0 引言
香蕉(Musaspp.)为芭蕉科(Musaccac)芭蕉属(MusaL.)多年生大型草本植物,是仅次于水稻、小麦、玉米的第四大粮食作物,其出口贸易量居世界第二,是全球重要的水果作物[1-2]。香蕉起源于东南亚热带亚热带地区和太平洋西部,经过人类数千年的驯化传播,目前,已被广泛种植,其种植面积超过百万hm2[2-3]。其中,中国香蕉种植面积为40万hm2,是全球第二大香蕉生产国[4-5],香蕉产业的发展对中国经济发展影响重大,一直以来香蕉产业发展受叶斑病、枯萎病、黄斑弄蝶、花蓟马等病虫害的制约[6-7],其中香蕉枯萎病的影响最大,然而现有的化学防治药剂在大田实验中效果不明显;生物菌防治虽能抑制病原菌的生长蔓延,但仍未解决如何在不降低拮抗菌的数量和活力的同时延长生物菌肥保质期的关键难题[8]。因此选育高抗品种成为发展香蕉产业的重要研究方向[9]。目前的香蕉栽培品种是由尖叶蕉(Musa acuminata)和长梗蕉(Musa balbisiam)种内或者种间杂交后代进化而成,多为三倍体[10]。三倍体香蕉存在高度不育,品种退化,以及面对逆境、病虫害等不利因素时抗性较弱等问题[11-12]。因此,开展香蕉种质资源遗传多样性分析,对寻找抗性材料和香蕉品质改良具有重要意义。目前有关香蕉种质资源的研究主要通过形态学标记、细胞学标记、生化标记、分子标记等方法[1,13-14]。形态学标记较方便直观是最早使用且沿用至今的方法。1962年Simmonds以25个核心性状为依据首次制定了香蕉形态学分类法,该方法成为了当时芭蕉属野生种质分类的主体方法;1996年,国际香(大)蕉改良网络组织出版了香蕉种质描述规范《Descriptors for Banana(Musaspp.)》一书,使形态分类法成为了香蕉种质资源分类的主体方法;2009年黄秉智编著《香蕉种质资源描述规范和数据标准》,该标准沿用至今,对香蕉种质资源研究的意义重大[12]。根据形态学分类法,姜燕[15]对中国主要蕉类进行种质的鉴定分类,明确其分类地位。赖钟雄等[16]对福州野生蕉进行植物学性状调查,并将其确定为“AA类群”。龙兴等[17]对广西野生蕉类种质资源进行鉴定,结果表明,供试野生蕉种质均为二倍体,均属芭蕉属真蕉组(Eumusa),其中,2份野生蕉种质为M.acuminatasubsp.malaccensis种,其余12份归属于M.itinerans种。余智城等[18]对福建漳州香蕉种质资源进行表型性状研究得出香牙蕉组根据假茎高度的不同可被细分成3小类。冯慧敏等[19]对12份野生蕉进行聚类分析,结果表明,根据染色体数,供试材料聚为2类,第一类为观赏蕉组和真蕉组,第二类为红花蕉组,根据叶柄基部、叶鞘光滑程度、果柄长短等性状可将第一类分为2个亚类。形态特征是基因的表现型,表型多样性是物种多样性的重要表现形式,对表型多样性进行评价是研究种质资源的基础[20]。叶是植物进行光合作用制造有机养料的主要器官[21],直接影响植物健康生长和果实产量。对香蕉种质资源叶片表型性状进行分析,较为全面的了解香蕉叶片表型性状的多样性,对香蕉品种选育意义重大。本研究对云南省红河热带作物研究所香蕉资源圃保存的香蕉种质资源叶型性状进行观察和测量;明确其叶片表型多样性、变异系数,并将试验所用的香蕉种质资源进行相关性分析和聚类分析,以期为云南香蕉品种改良和新品种选育提供参考。
1 材料与方法
1.1 试验材料
本试验所用香蕉种质资源均保存于云南省红河热带农业科学研究所香蕉资源圃,资源圃位于22°57′N,103°88′E,海拔93 m;用于调查的植株均在抽蕾期;观察、测定叶片均为第三片叶;且所有植株均栽植于同一地块,生长环境条件一致。供试香蕉种质材料共49份,详细信息见表1。
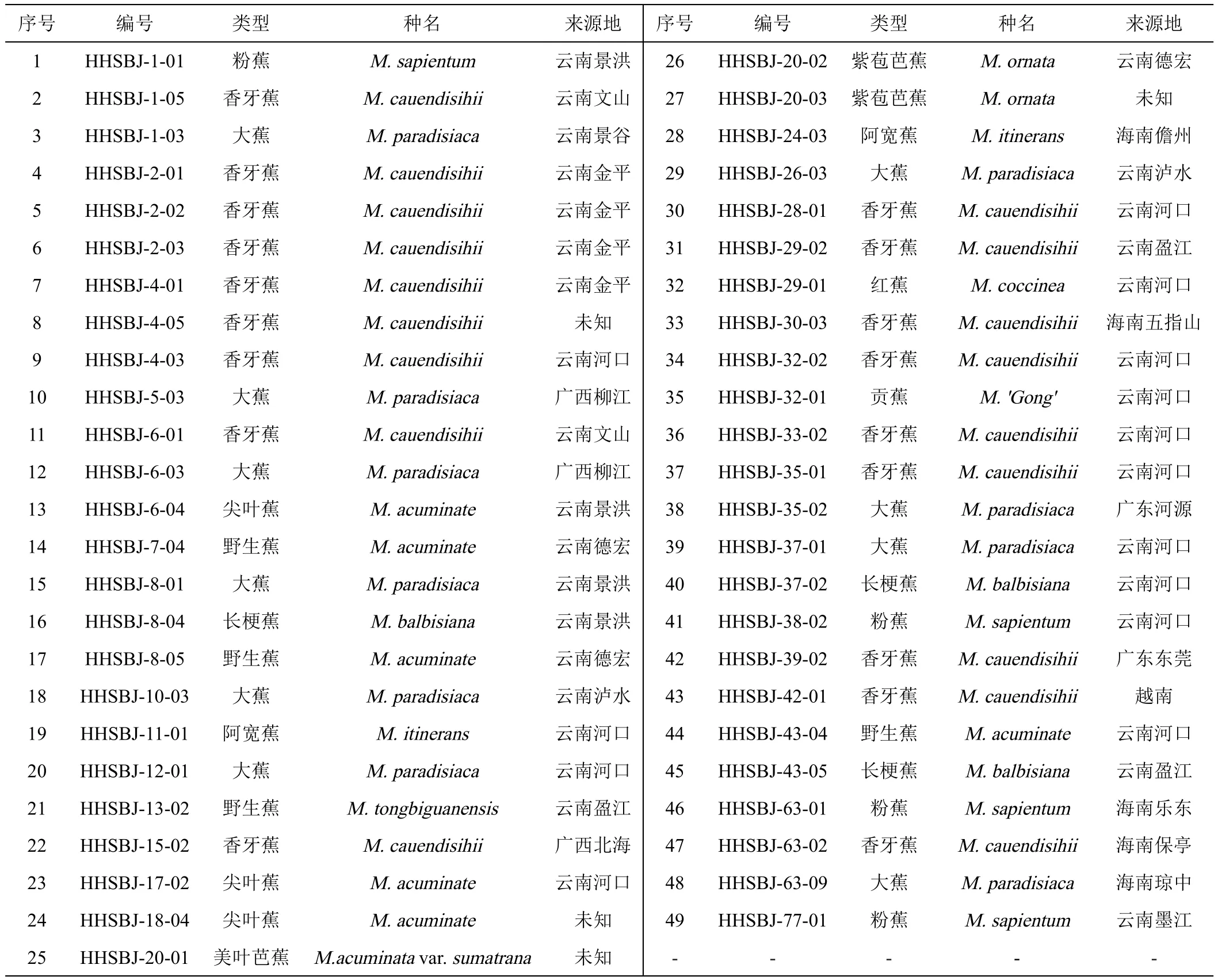
表1 供试材料名称及来源地
1.2 调查方法
试验于2020—2021年在云南省红河热带农业科学研究所香蕉资源圃内进行,18个性状调查标准参考《香蕉种质资源描述规范》[22];4个数量性状(叶柄长、叶片长度、叶片宽度、叶形比)用卷尺进行测量并计算,14个质量性状通过赋值法进行分级(表2)。
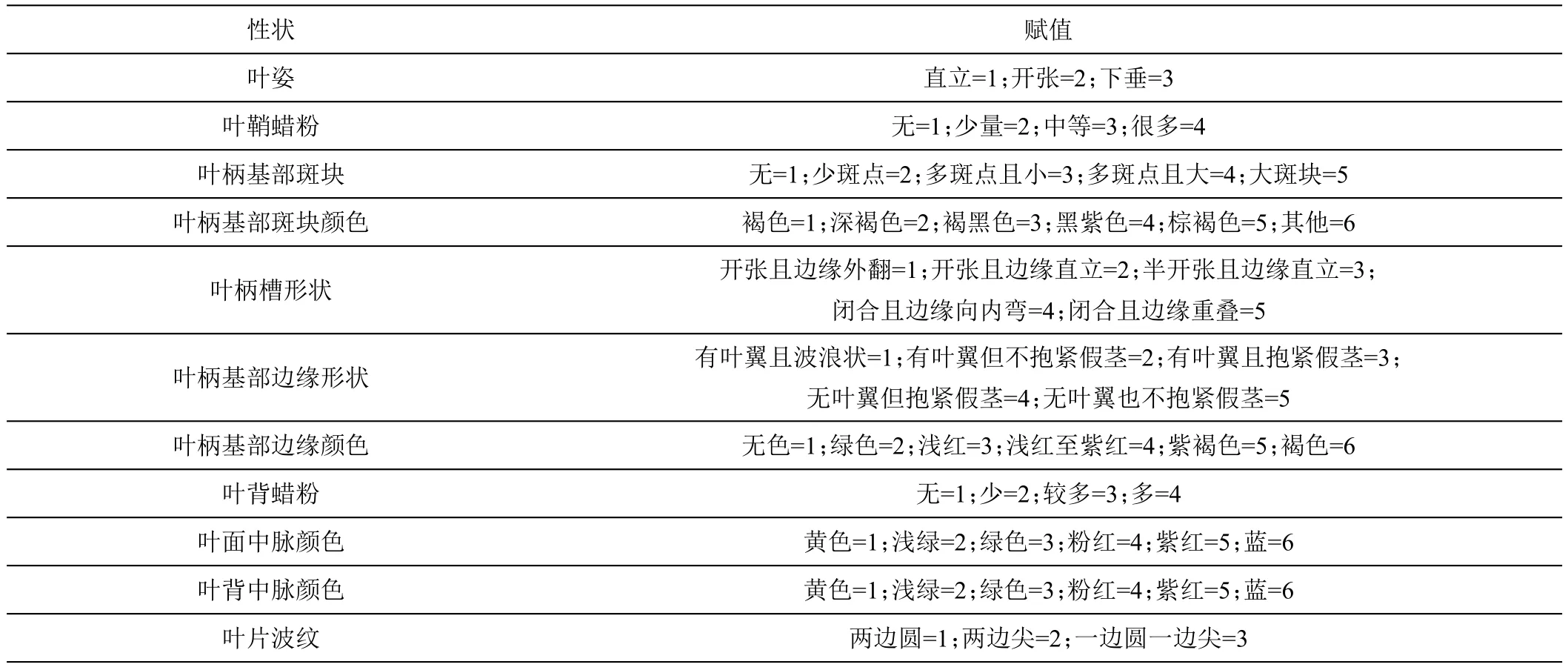
表2 香蕉种质资源叶片表型质量性状的赋值描述

续表2
1.3 数据处理
用Excel软件对调查数据进行汇总及整理,计算质量性状的频率分布和遗传多样性指数(H'),以及数量性状的最大值(Max)、最小值(Min)、平均值(X)、标准差(S)、变异系数(CV)、频率分布和遗传多样性指数(H')。变异系数计算见公式(1)[23]。

遗传多样性指数采用Shannon-Weaver信息指数,计算见公式(2)。

其中Pi为某一性状第i级出现的频率[24]。计算数量性状遗传多样性指数时,首先对数量性状进行分级,根据平均值、标准差将测定数据分为10级,分级区间在[X-2s,X+2s],相连两级间相差0.5 s,根据种质数量在区间内的频次分布,计算出频率,并进一步计算多样性指数[25]。用SPSS21.0软件完成数据的相关性分析、主成分分析及聚类分析。
2 结果与分析
2.1 香蕉种质资源叶型性状多样性分析
2.1.1 质量性状多样性分析 由表3可看出,14个质量性状的H'为0.269~1.455,均值为0.97。其中,叶柄基部边缘形状多样性指数最高(1.455),其次是叶柄基部边缘颜色(1.454),二者的频率分布均表现出较高的离散程度,表明该批香蕉种质叶柄基部边缘形状类型和叶柄边缘颜色丰富。叶背蜡粉的多样性指数为1.314,种质性状类型分布相对均匀。叶柄槽性状多样性指数为1.309,表型性状频次分布主要为开张且边缘外翻。叶鞘蜡粉(1.183)、叶柄基部斑块颜色(1.175)、叶柄基部斑块(1.055)3个性状多样性指数接近,说明它们的多样性和丰富度接近。叶片基部形状(0.906)、叶片基部对称性(0.834)、叶背中脉颜色(0.722)、叶翼干枯类型(0.683)、叶片波纹(0.616)、叶姿(0.572)、叶面中脉颜色(0.269)7个性状的多样性指数均小于1,说明所评价的香蕉种质在这些性状上的多样性程度相对较低。表型性状频率分布显示类型比较集中的几个性状中,叶面中脉颜色以绿色为主,占比93.88%,叶姿以开张为主,占比79.53%,叶背中脉颜色以浅绿色为主,占比73.47%。
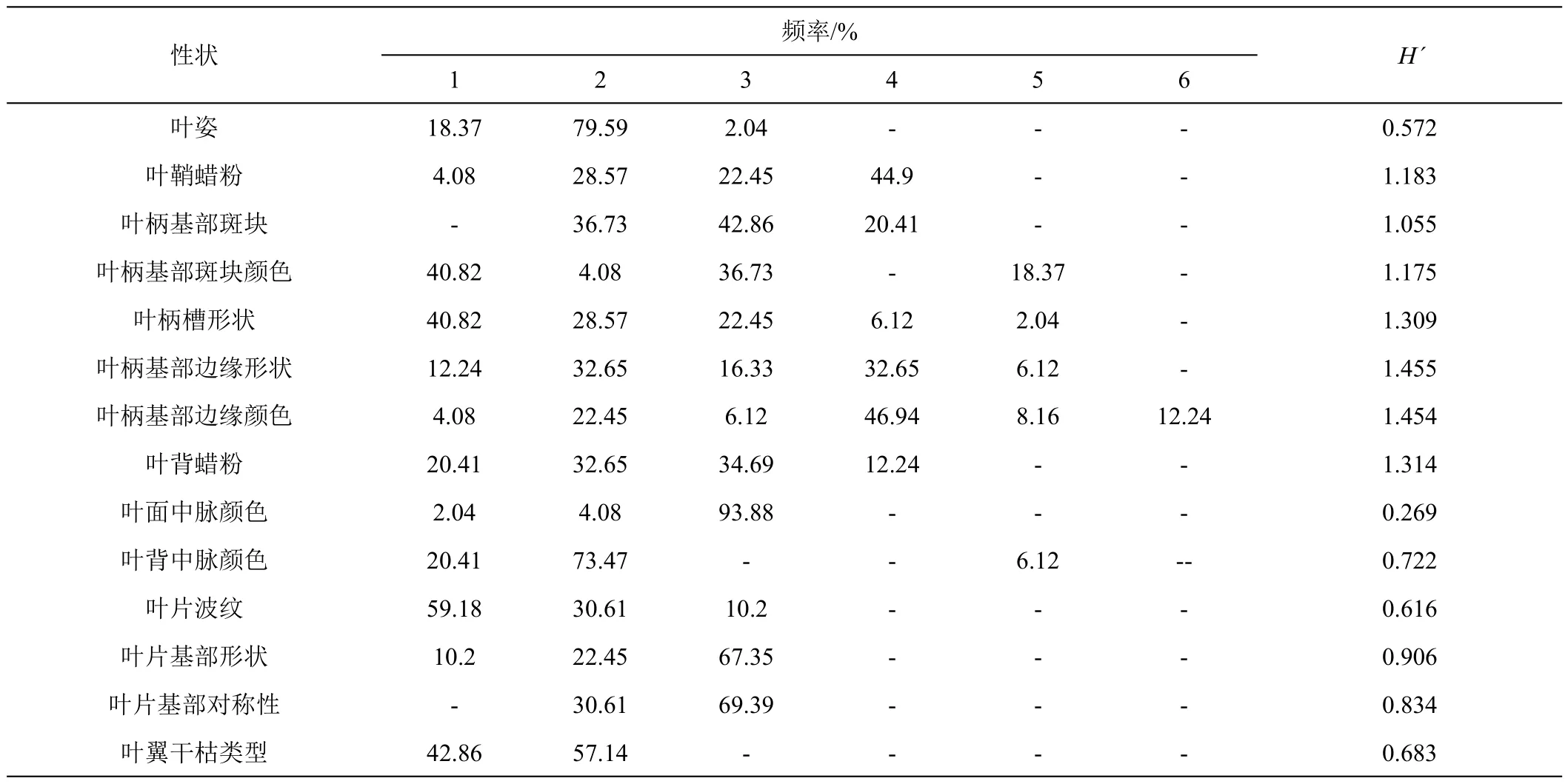
表3 香蕉种质资源叶片表型质量性状的分布频率
2.1.2 数量性状多样性分析 由表4可知,4个数量性状的H'为1.88~2.24,均值为2.11,说明4个数量性状均具有丰富的遗传多样性。4个数量性状变异系数范围在19.29%~33.46%,变异系数由大到小依次为叶柄长(33.46%)、叶形比(20.62%)、叶片宽(20.47%)、叶片长(19.29%)表明所评价的香蕉种质在4个数量性状上的变异幅度较大,多样性程度较高。
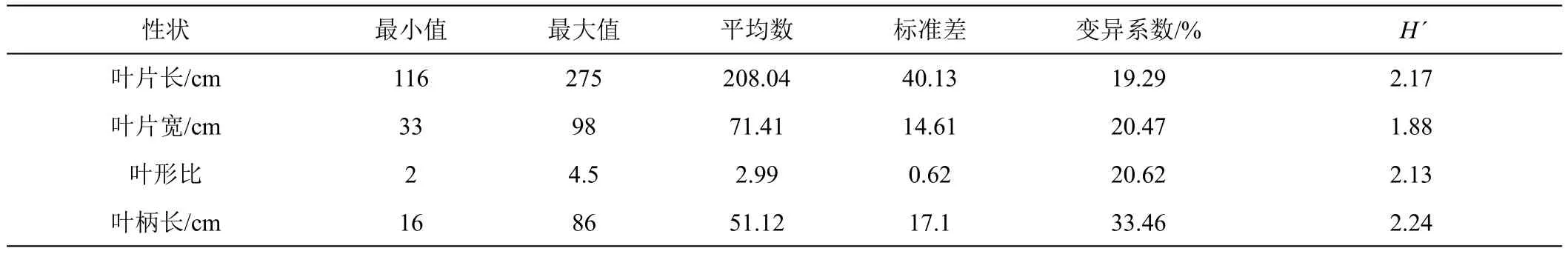
表4 香蕉种质资源叶片表型性状遗传多样性分析
2.2 香蕉种质资源表型性状的相关性分析
对49份香蕉种质的18个性状进行双变量相关性分析(表5),结果显示,31对性状相关性达到极显著水平(P<0.01),18对性状相关性达到显著水平(P<0.05)。其中,叶翼干枯类型、叶形比、叶柄长两两呈极显著正相关;叶鞘蜡粉、叶背蜡粉、叶面中脉颜色两两呈极显著或显著正相关;叶柄基部边缘形状和叶柄槽形状分别与叶翼干枯类型、叶片长、叶形比、叶柄长极显著正相关,且二者互为极显著正相关;叶姿和叶背蜡粉分别与叶片波纹、叶片宽极显著正相关,且二者互为显著正相关;叶片基部形状仅与叶片基部对称性极显著正相关,与叶片波纹极显著负相关;叶柄边缘颜色仅与叶柄基部斑块颜色显著负相关,叶片基部对称性仅与叶片基部形状极显著相关;叶背中脉颜色与叶片宽极显著负相关,叶片宽与叶形比极显著负相关,叶背中脉颜色与与叶形比极显著正相关。18个性状间存在不同程度的相关性,彼此之间互相影响关系密切。
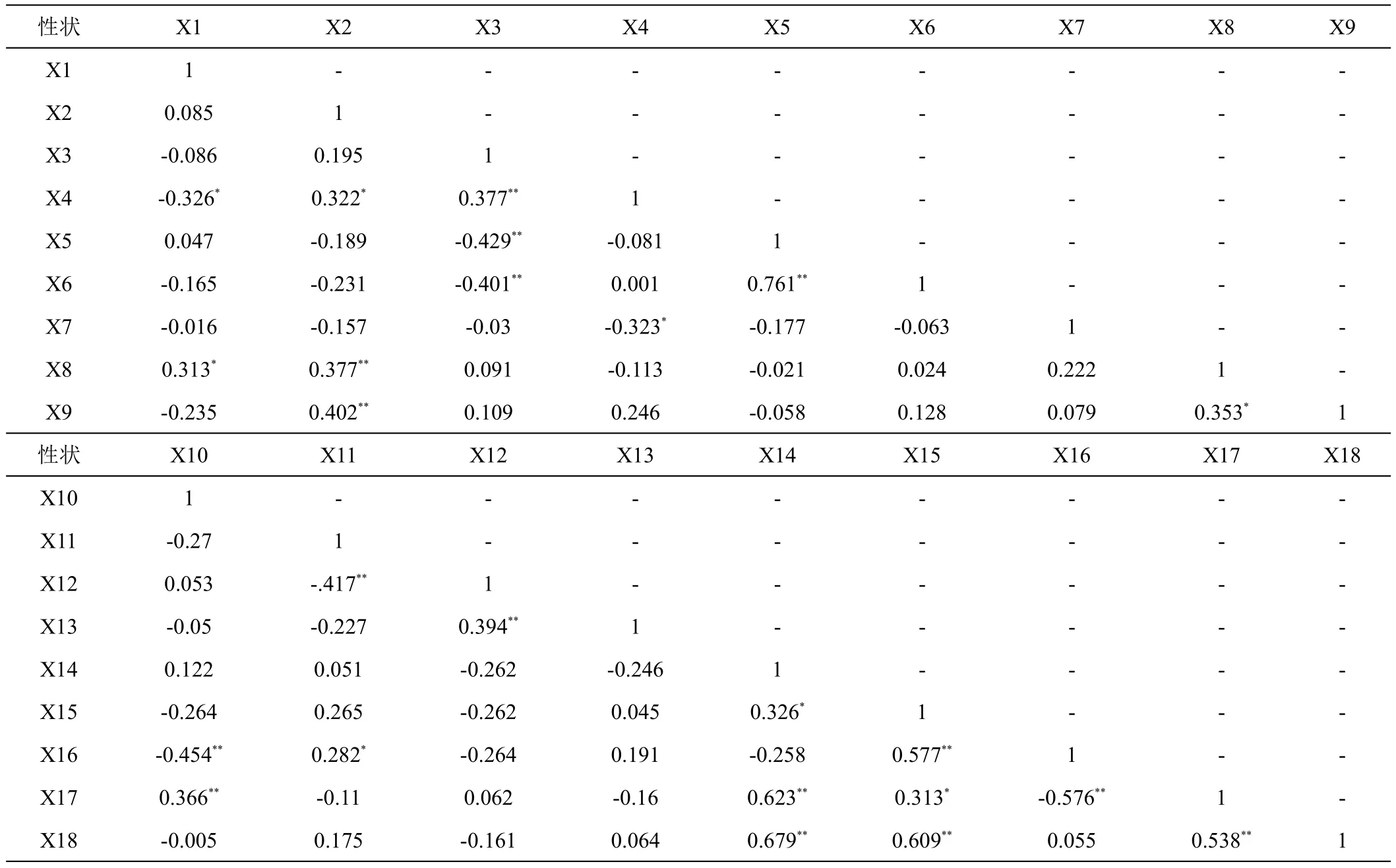
表5 49份香蕉种质叶片表型性状相关性
2.3 香蕉种质资源叶片表型性状的主成分分析
对49份香蕉种质资源的18个叶型性状进行主成分分析(表6),结果表明,前6个主成分贡献率分别为25.31%、19.31%、11.40%、10.40%、6.73%、5.64%,累计贡献率达78.80%,表明这6个主成分代表了18个叶片表型性状的大多数信息。第1主成分主要由叶柄槽形状、叶柄基部边缘形状、叶翼干枯类型、叶柄长4个性状决定,贡献率最大。第2主成分中叶姿、叶背蜡粉、叶片宽的特征向量较高,是该主成分的主要性状。第3主成分中叶鞘蜡粉、叶面中脉颜色的特征向量较大,说明该主成分由这3个性状决定。第4、5、6主成分中分别是叶片基部对称性、叶鞘蜡粉、叶柄边缘颜色的特征向量最高,为各主成分的主要贡献指标。

表6 香蕉种质资源叶片表型性状的主成分分析
2.4 香蕉种质资源叶片表型性状的聚类分析
采用离差平方和法(Ward法)对49份香蕉种质资源进行基于欧式遗传距离的聚类分析(图1)。结果表明:在欧氏遗传距离为10处可将供试材料分为4个类群,第I类群为野生蕉和观赏蕉,共4份,占供试材料的8.16%,该类群主要表现为叶姿直立、叶柄基部斑块呈棕褐色、叶柄基部有叶翼且抱紧、叶片基部明显不对称、叶形比最大,其中,叶姿、叶柄基部斑块颜色、叶柄基部边缘形状为该类群明显区别于其他类群的主要性状;第II类群多为大蕉和粉蕉,共14份,占供试材料的28.57%,主要特征为叶姿开张、叶翼不干枯、叶片最长、最宽、叶柄最长;第III类群共13份,占供试材料的26.53%,其叶形比和叶柄长处于居中水平,为除第I类群外的所有野生蕉,其中包括1份‘红蕉’,1份‘美叶芭蕉’;第IV类群为香牙蕉,共18份,占供试材料的36.73%,主要表现为叶姿开张、叶柄槽开张且边缘外翻、叶柄边缘颜色多为浅红至紫、叶翼干枯、叶柄最短、叶形比最小,该类群最显著的特征为叶柄槽开张且边缘外翻(表7)。
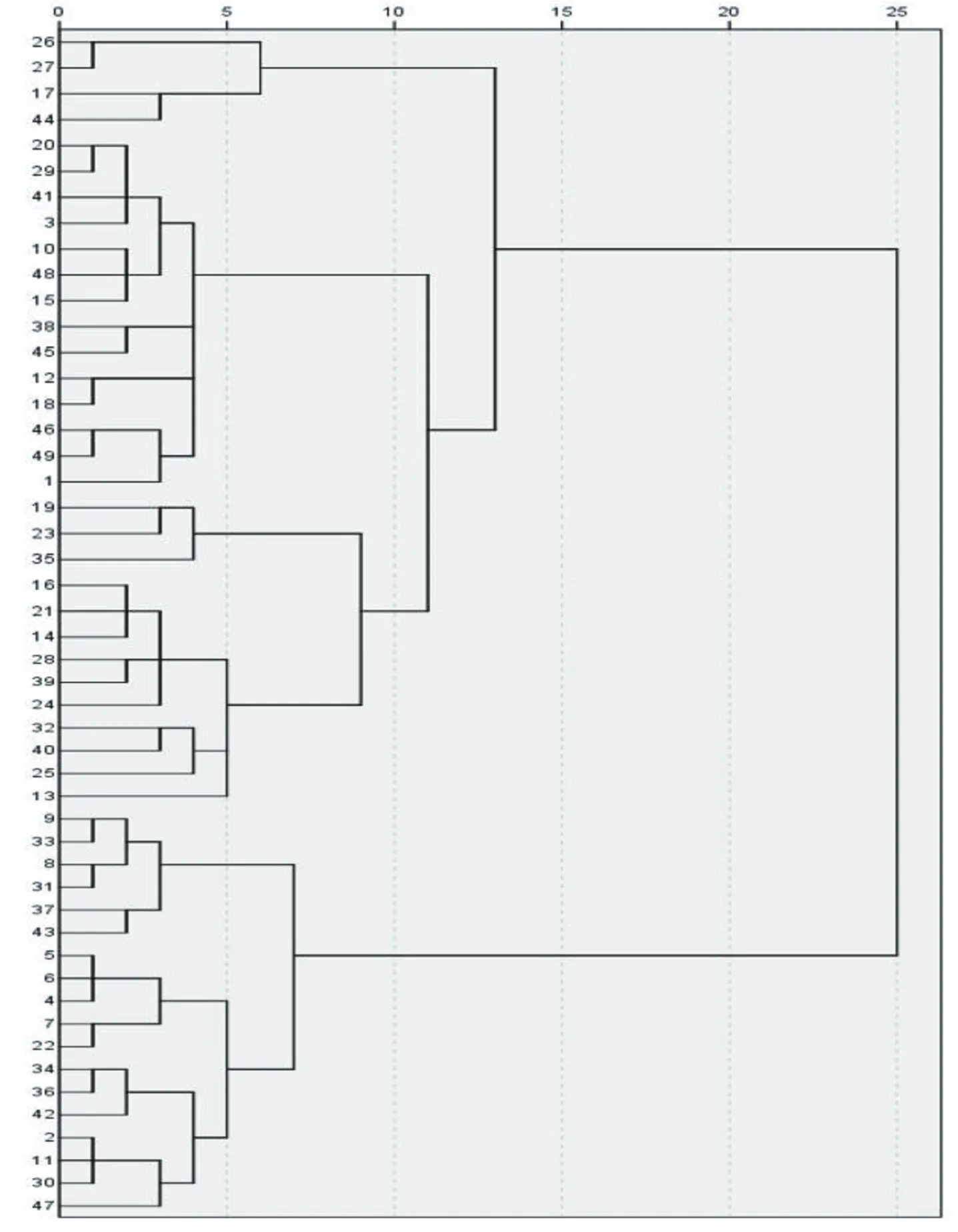
图1 49份香蕉种质聚类分析图

表7 香蕉种质资源各类群叶型性状特征和平均数
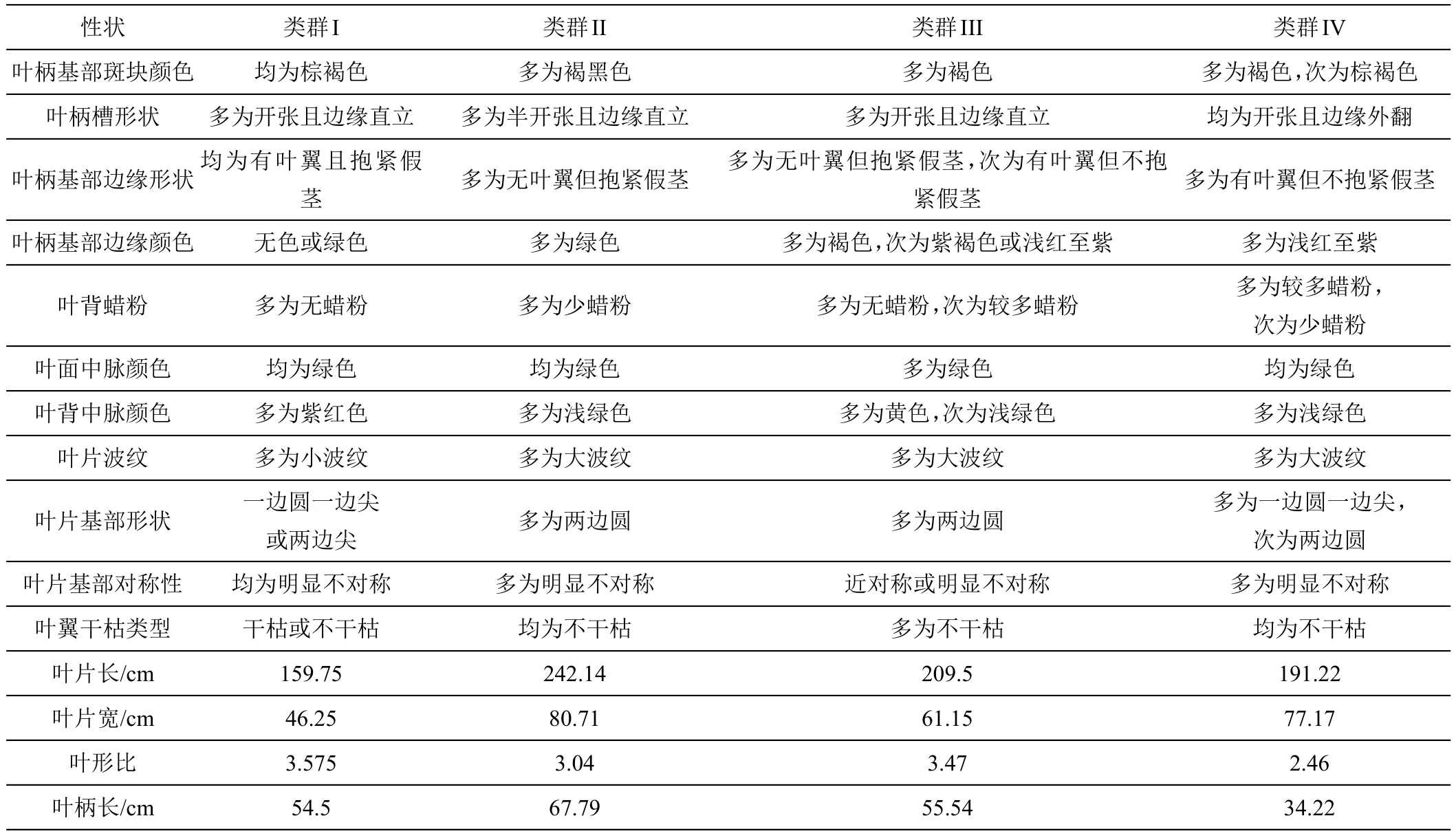
续表7
3 讨论与结论
表型性状是基因与环境共同影响的结果,表型性状的变异分析、相关性分析、主成分分析和聚类分析可以较为全面地了解种质资源的多样性[26]。Shannon-Wiener多样性指数是评价种质资源多样性的一个重要指标,现已广泛应用于种质资源多样性分析[27]。本研究利用其对香蕉种质叶片表型性状进行多样性分析发现数量性状的平均多样性指数远高于质量性状的平均多样性指数,且单个数量性状的多样性指数均高于各质量性状的多样性指数。这与李峰等[28]对67份慈姑种质资源进行表型多样性分许,发现叶片形状、球茎形状、球茎皮色的多样性指数均低于其他数量性状多样性指数的结果相似;与王治会等[29]通过对35份茶树种质资源的21个芽叶表型性状进行多样性研究,发现数量型性状的平均多样性指数高于描述性性状平均多样性指数的结果一致。究其原因可能是数量性状易受环境因素影响,与质量性状相比不够稳定,从而导致数量性状变异更丰富,多样性指数较高。变异系数反映了各性状数据的离散程度,有研究证明,变异系数大于10%时,样本间差异明显[30]。本研究中数量性状变异系数分别为叶柄长(33.46%)、叶形比(20.62%)、叶片宽(20.47%)、叶片长(19.29%),均大于10%,说明香蕉种质资源叶片表型多样性丰富,对后续的开发利用有积极意义。进一步进行相关性分析后发现各性状间均存在不同程度的相关关系,这与郭琪等[25]对山西刺槐种质资源的叶片表型多样性分析结果相似。各性状间相互影响,在进行性状改良及选育种时,需将各性状间密切的相关关系考虑在内。
主成分分析可以将多个性状指标简化为几个能反映较多信息的综合评价指标[23]。本研究中18个性状简化为6个主成分,各主成分的贡献指标为叶姿、叶鞘蜡粉、叶柄槽形状、叶柄基部边缘形状、叶柄边缘颜色、叶面中脉颜色、叶背蜡粉、叶片基部对称性、叶翼干枯类型、叶柄长、叶片宽。6个主成分累计贡献率为78.80%,11个叶片表型性状反映了香蕉种质的大多数信息,为香蕉叶片表型多样性评价的关键指标,可为香蕉选育种提供参考[20]。聚类分析明确了香蕉种质的类型,本研究中香芽蕉聚为一个类群,大蕉、粉蕉聚为一个类群的结果与谭卫萍等[31]应用AFLP技术对60个栽培蕉和野生蕉种进行聚类分析得到的结果一致。唐敏慧[32]基于SSR标记对66个香蕉品种的聚类分析,发现AA、AAB、ABB组香蕉聚为一类,得出了部分栽培种粉蕉与野生二倍体香蕉亲缘关系较近。陈红俊[33]通过对福州和泉州5个类型野生蕉的7个自然居群进行ISSR遗传多样性分析,发现居群3、4、5与居群2、7聚在一起后,得出宦溪野生蕉与福州北峰野生蕉、永春野生蕉亲缘关系比较近。本研究聚类分析结果中‘红蕉’、‘美叶芭蕉’和‘长梗蕉’被聚为同一类群,推测这可能是三者间的亲缘关系比较近,也可能是与本研究仅从叶片表型性状进行探讨有关。为进一步确定香蕉种质的间关系,接下来将从花、果实等具有传统分类价值的性状进行研究。
