茎秆基部节间特性和木质素合成与燕麦抗倒伏的关系
2022-11-23南铭李晶赵桂琴柴继宽刘彦明
南铭,李晶,赵桂琴,柴继宽,刘彦明
(1.甘肃省农业科学院作物研究所,甘肃 兰州 730070;2.甘肃省定西市农业科学研究院,甘肃 定西 743000;3.甘肃农业大学草业学院,甘肃 兰州 730070)
燕麦(Avena sativa)是禾本科一年生粮饲兼用型作物,因其抗寒、耐旱、耐盐碱、适应性强,具有生产性能突出及营养价值丰富等特点,在世界5大洲42个国家都有广泛栽培[1]。我国主要分布在东北、华北、西北中部及西南高寒地区,年播种面积保持在80万hm2左右。燕麦适口性好,消化率高,青刈后茎叶鲜嫩多汁,适宜青饲或调制干草[2],是西北干旱半干旱地区及农牧交错地带家畜的主要饲草料来源之一[3],对修复退化天然草场、保护农业生态环境和促进草牧业健康发展具有不可替代的作用[4]。
在燕麦的生长发育过程中,倒伏是很常见的现象。倒伏破坏燕麦群体结构与个体茎秆疏导系统,造成养分和水分的运输受阻[5],降低燕麦产量和品质,使之质量变弱,其已经成为目前燕麦稳产栽培的主要制约因素之一,也是燕麦生产中普遍面临的问题[6]。茎秆形态特征作为判别作物倒伏与否的重要表型性状,与作物抗倒伏有关,也是研究抗倒伏性的一个主要方向。在一定株高范围内,恰当的茎秆节间配置和合理的基部节间结构能显著提高茎秆质量,增强茎秆机械强度,从而达到高产和抗倒伏的有效平衡[7]。很多学者结合形态结构的差异性,通过对小麦(Triticum aestivum)[8]、水 稻(Oryza sativa)[9]、玉米(Zea mays)[10]、大豆(Glycine max)[11]、荞麦(Fagopyrum esculentum)[12]、青稞(Hordeum vulgare)[13]、油菜(Brassica napus)[14]等作物茎秆生理学特点的分析,认为作物茎秆木质素和纤维素含量较高,其茎秆刚度和机械强度大,植株抗倒伏能力强。木质素作为植物次生细胞壁的组分之一,填充于纤维素构架中主要用来增强茎秆细胞壁的机械强度[15],能增强作物茎秆的抗压强度和弯曲性能,其含量与作物抗倒伏能力强弱相关,增加茎秆中木质素含量是实现作物抗倒伏能力提高的有效途径[16]。近些年来,对于茎秆中木质素合成与作物抗倒伏关系的研究越来越受到关注,但是围绕茎秆木质素合成与燕麦抗倒伏关系的研究鲜见报道。本试验选用田间表现稳定且表型性状存在差异的不同倒伏类型燕麦材料,通过茎秆形态特征和生理指标及木质素合成酶活性的研究,明确茎秆中木质素合成与燕麦抗倒伏能力的相互关系,以期为燕麦抗倒伏品种选育提供依据。
1 材料与方法
1.1 试验地点
试验地位于甘肃省定西市农业科学研究院试验基地(E 104°42′,N 35°32′),海拔1920 m,无霜期105 d左右,年均气温8.3℃,降水量350 mm,土壤为黄绵土,容重1.3 g·cm-3,有机质1.61%,速效氮58.0 mg·kg-1,速效磷7.38 mg·kg-1,速效钾141.64 mg·kg-1,pH值8.21。
1.2 试验设计
抗倒伏材料为蒙燕1号、坝燕6号燕麦,易倒伏材料为定燕2号和青引2号燕麦。采用随机区组设计,小区面积为10 m2(5 m×2 m),3次重复,人工条播种植,行距25 cm,2017-2018年分别在4月上旬播种,8月上旬收获,田间管理同大田示范。
1.3 研究方法
燕麦进入抽穗期,每个品种选取株型一致、抽穗整齐的植株20株,对主茎进行挂牌标记。灌浆期每个品种随机选取10株挂牌且未发生倒伏的单株,分别剪取主茎基部第2伸长节间,5株用于测定茎秆基部第2伸长节间长度、茎粗、秆壁厚及茎秆重量等,其余置液氮中极速冷冻,存于-80℃超低温冰箱用于测定纤维素、木质素及其合成酶活性。
茎秆密度:称取基部第2节间去叶(鞘)鲜重,在105℃条件下杀青2 h并在60℃下烘干至恒重,用基部第2节间长度、内径、壁厚、茎粗计算基部第2节间体积(中空圆柱体,不含髓腔),茎秆密度=第2节间干重(g)/第2节间体积,茎秆充实度=第2节间干重/第2节间长度。茎秆穿刺强度测定利用茎秆强度测定仪(YYD-1A,浙江托普云农科技有限公司),携带穿刺探头(横断面积为1 mm2),将第2茎节除去叶鞘,置于凹槽内,两支点间距5 cm,以平稳的力垂直向下匀速插入节间中部,探头瞬间穿破进入茎秆时记录显示的最大值即为茎秆穿刺强度。茎秆抗折力测定利用茎秆强度测定仪(YYD-1A,浙江托普云农科技有限公司),将茎秆基部第2伸长节间置仪器支撑架凹槽内部,两支撑点的间距为5 cm,茎秆瞬间折断时峰值即为茎秆抗折力。
茎秆中木质素含量测定参照池宁琳[17],单位为百分比(%)。茎秆中酸性洗涤纤维(acid detergent fiber,ADF)和中性洗涤纤维(neutral detergent fiber,NDF)含量测定参照Van Soest等[18]的方法,所测值为干物质的百分比(%)。纤维素(cellulose,CLL)含量利用北京索莱宝科技有限公司生产的试剂盒检测,参照陈晓光等[9]利用可见分光光度法及酶标仪方法测定苯丙氨酸转氨酶(phenylalanine aminotransferase,PAL)、酪氨酸解氨酶(tyrosine ammonia lyase,TAL)、肉桂醇脱氢酶(cinnamyl alcohol dehydrogenase,CAD)和4-香豆酸:CoA连接酶(4-coumaric acid:coa ligase,4CL)的活性。
燕麦进入成熟期按照任长忠等[19]的方法调查燕麦田间倒伏率(lodging rate,LR),评定倒伏分级(lodging degree,LD)。田间倒伏率=实际倒伏植株数/小区总植株数×100%。倒伏级别分为4级:0级:未倒伏;1级:0~15%倒伏;2级:15%~45%倒伏;3级:45%以上倒伏。倒伏面积按照实际倒伏面积占小区总面积的百分率(%)表示。
1.4 数据处理
采用Microsoft Excel 2019进行数据整理以及作图,用SPSS 22.0软件进行统计分析。
2 结果与分析
2.1 燕麦倒伏发生情况与茎秆抗折力的关系
抗倒伏品种蒙燕1号和坝燕6号在全生育期内未发生倒伏,田间群体植株实际倒伏率均为0,倒伏级别均为0级。易倒伏品种青引2号进入灌浆期初就开始出现倒伏,成熟收获前田间实际倒伏率达到93.70%。易倒伏品种定燕2号进入乳熟期后发生倒伏现象,成熟期田间实际倒伏率略低于青引2号,倒伏级别均为3级(表1)。不同倒伏类型燕麦品种茎秆基部第2伸长节间长度存在极显著(P<0.01)差异,抗倒伏品种组茎秆基部第2伸长节间长度较易倒伏品种组平均小34.11%,而茎粗、秆壁厚、茎重、密度、充实度分别为易倒伏品种组的1.41、1.35、1.35、1.39和1.53倍,相比易倒伏燕麦品种组,抗倒伏燕麦品种组茎秆基部第2伸长节间抗折力和穿刺强度较易倒伏品种组平均高62.10%。抗倒伏品种茎秆基部第2伸长节间抗折力和穿刺强度较易倒伏品种分别高36.23%、36.60%,易倒伏品种较抗倒伏品种茎秆基部第2伸长节间茎粗小29.11%。
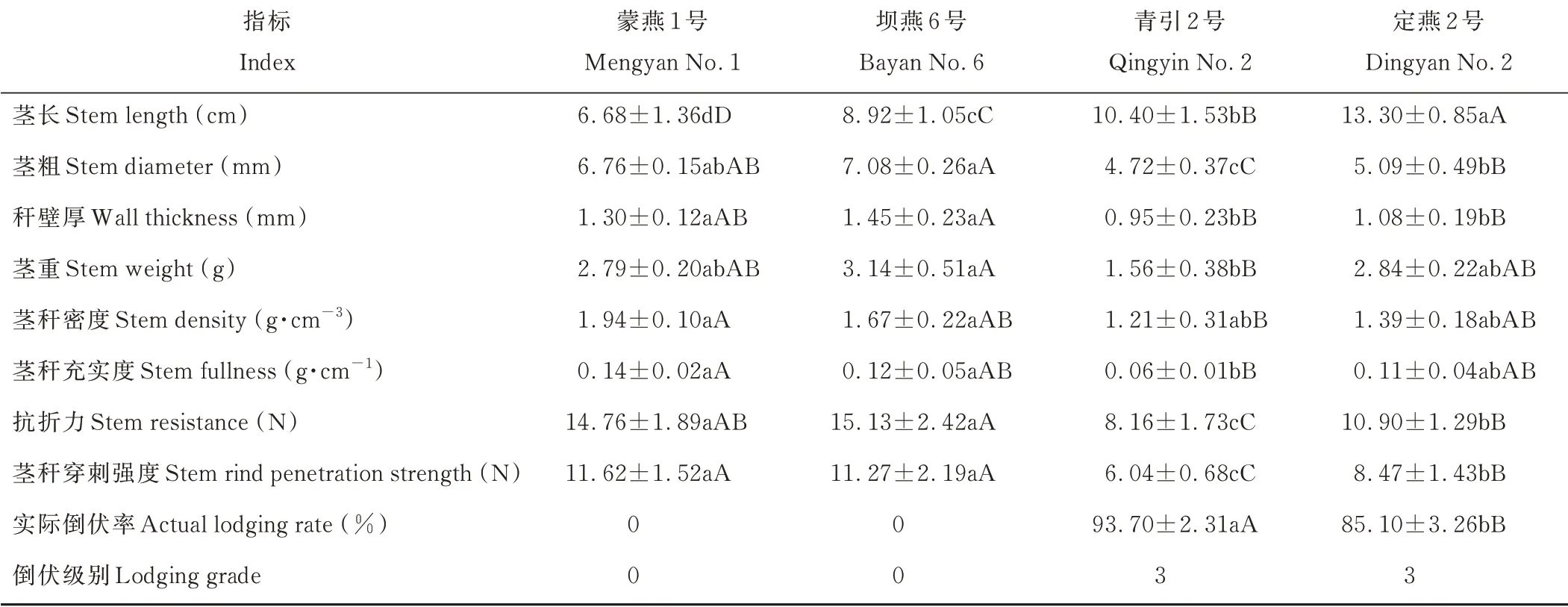
表1 燕麦茎秆指标与抗折力和实际倒伏率Table 1 Stem index and breaking resistance and actual lodging rate of oat varieties
2.2 茎秆中木质素和纤维素含量
通过比较发现,抗倒伏品种(KD-1)茎秆基部第2伸长节间纤维素含量显著(P<0.05)高于易倒伏品种组,抗倒伏品种(KD-2)茎秆基部第2伸长节间木质素含量显著(P<0.05)高于易倒伏品种组(图1)。相比易倒伏品种组,抗倒伏品种组茎秆基部第2伸长节间木质素、纤维素含量增幅分别为9.2%~27.6%、19.40%~29.19%,抗倒伏品种(KD-1)与易倒伏品种(YD-1)基部第2伸长节间木质素含量存在显著(P<0.05)差异,抗倒伏品种组与易倒伏品种(YD-1)基部第2伸长节间纤维素含量存在显著(P<0.05)差异,说明茎秆中木质素和纤维素含量与燕麦茎秆抗倒伏能力有关,但影响并不明显。
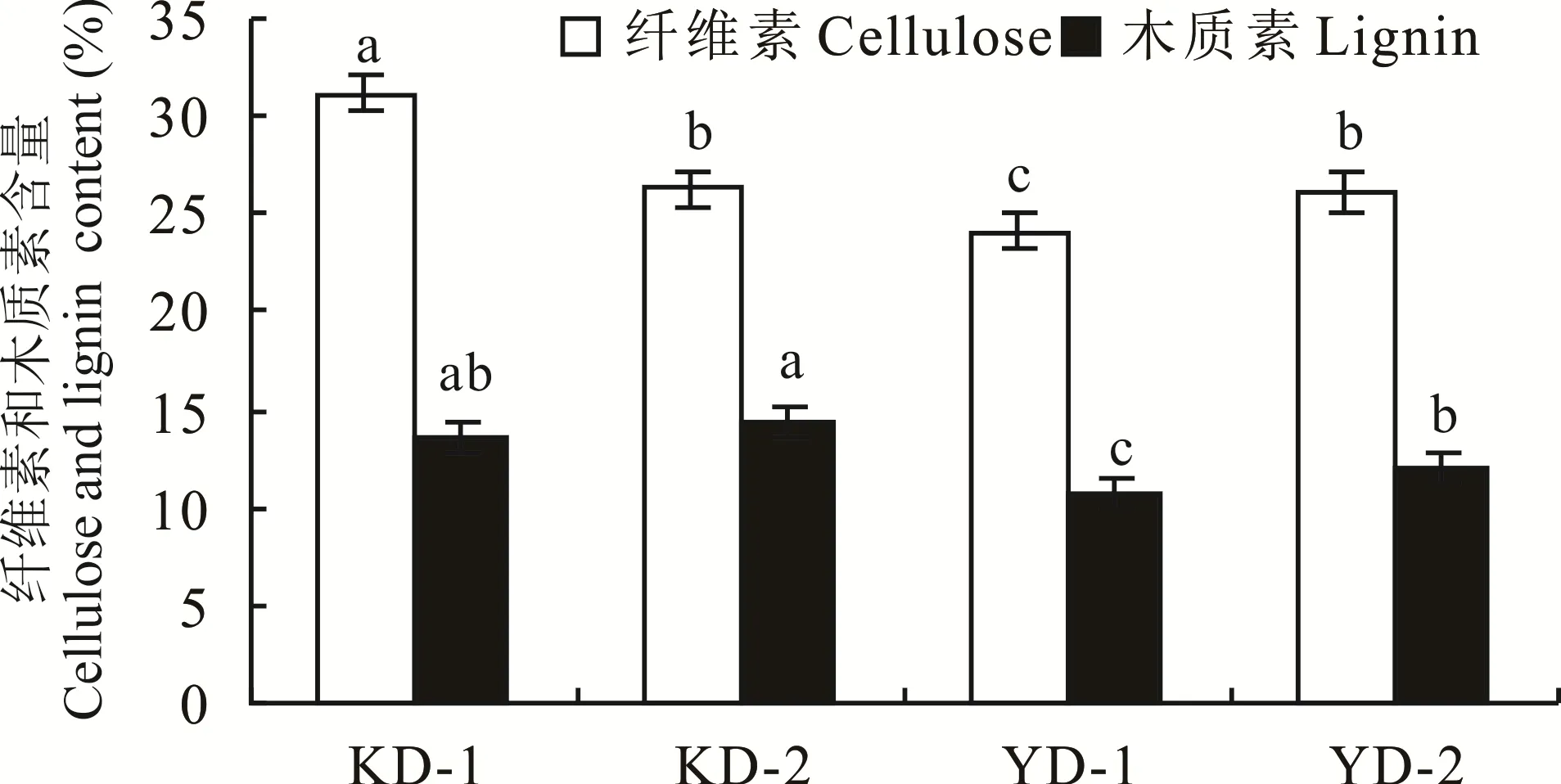
图1 不同燕麦品种茎秆中纤维素和木质素含量Fig.1 Cellulose and lignin content of stem in different oat varieties
2.3 茎秆中中性洗涤纤维和酸性洗涤纤维含量
灌浆期同一燕麦品种茎秆中中性洗涤纤维含量高于酸性洗涤纤维含量,不同倒伏类型燕麦品种之间茎秆中NDF和ADF的含量差异并不显著(图2)。说明中性洗涤纤维(NDF)和酸性洗涤纤维(ADF)的含量高低并不是诱发燕麦倒伏的关键指标,对燕麦茎秆抗倒伏能力的影响是间接的。
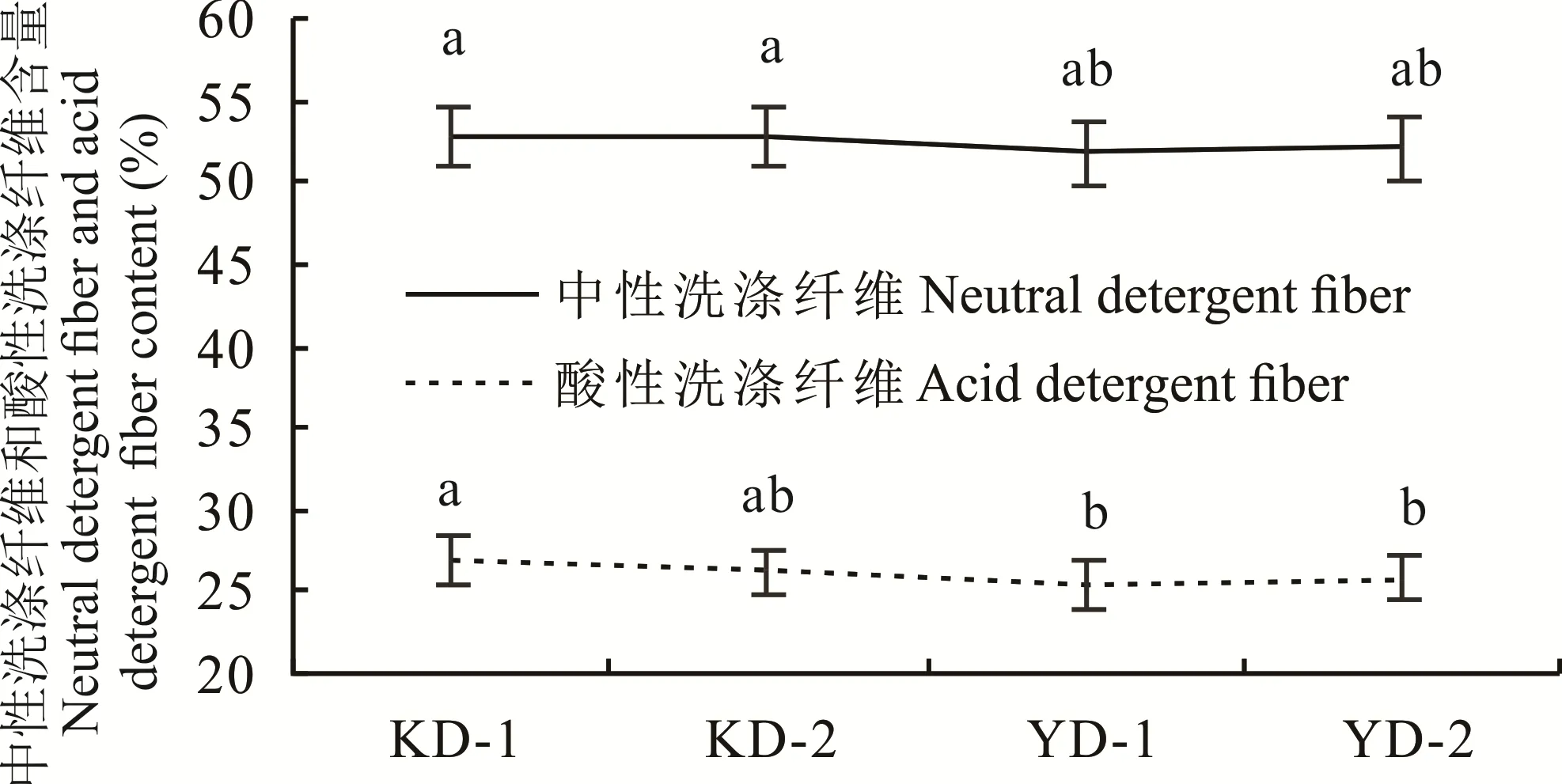
图2 燕麦茎秆中中性洗涤纤维和酸性洗涤纤维含量Fig.2 Neutral detergent fiber and acid detergent fiber content of the oat stems
2.4 茎秆中木质素合成相关酶活性
PAL、TAL、CAD和4CL是参与调控木质素生物合成全过程的4种主要酶类。抗倒伏品种茎秆中木质素合成酶活性普遍高于易倒伏品种(图3)。其中,易倒伏品种(YD-1)PAL、TAL、4CL和CAD的活性均较低,导致木质素积累量少,茎秆细胞壁硬度下降,支撑强度减弱。同一品种中PAL和CAD活性较高,TAL和4CL活性较低,不同倒伏类型品种间4种酶活性存在显著(P<0.05)差异,与倒伏敏感品种相比,抗倒伏品种茎秆中PAL、TAL、CAD和4CL的活性增幅分别为19.3%~71.9%、86.4%~204.8%、7.2%~22.0%和6.0%~46.1%,酶活性升高会增加茎秆木质素代谢通路中的代谢产物含量,进而促进茎秆基部节间合成和积累较多的木质素,增强茎秆细胞壁硬度和机械性能,增大茎秆抗折力,提高植株抗倒伏能力。
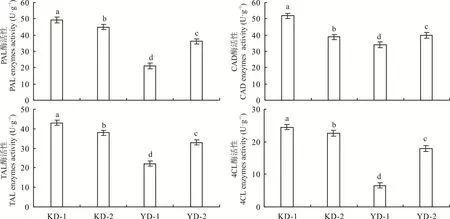
图3 不同燕麦品种茎秆中木质素合成相关酶活性Fig.3 Enzymes activity of lignin synthesis related of stem in different oat varieties
2.5 茎秆木质素及其合成酶活性与燕麦抗倒伏能力的相关性
燕麦抗倒伏能力与茎秆基部第2伸长节间木质素含量密切相关(表2),茎秆抗折力、茎秆穿刺强度与木质素含量极显著(r=0.9862,r=0.9638,P<0.01)正相关,而木质素含量与田间实际倒伏率呈显著(r=-0.9407,P<0.05)负相关关系。表明茎秆中木质素含量较高的燕麦品种,植株基部节间抗折力较大,田间倒伏程度较轻,抗倒伏能力较强。茎秆基部第2伸长节间长度与其抗折力、穿刺强度负相关但不显著(P>0.05),与田间实际倒伏率正相关也不显著(P>0.05)。茎粗和秆壁厚均与茎秆抗折力、茎秆穿刺强度极显著正相关(P<0.01),与田间实际倒伏率极显著或显著负相关(P<0.05),与木质素含量极显著正相关(P<0.01),表明茎秆中木质素含量较高的品种基部伸长节间明显较短、粗度较大、秆壁较厚、密度和充实度高,主茎基部第2伸长节间长、粗、厚及其构成显著影响茎秆抗折力和穿刺强度的大小。茎秆木质素含量与PAL、TAL、CAD和4CL活性均显著正相关(P<0.05),PAL、CAD活性与茎秆抗折力极显著(P<0.01)正相关,PAL、TAL、CAD活性和穿刺强度极显著正相关(P<0.01),而4CL活性与茎秆抗折力和穿刺强度不显著(P>0.05)正相关,PAL、TAL、CAD和4CL活性与田间实际倒伏率均呈负相关关系,表明提高PAL、TAL、CAD和4CL的活性是茎秆中木质素含量增加的重要酶学基础,也是增强燕麦茎秆抗倒伏能力的关键。
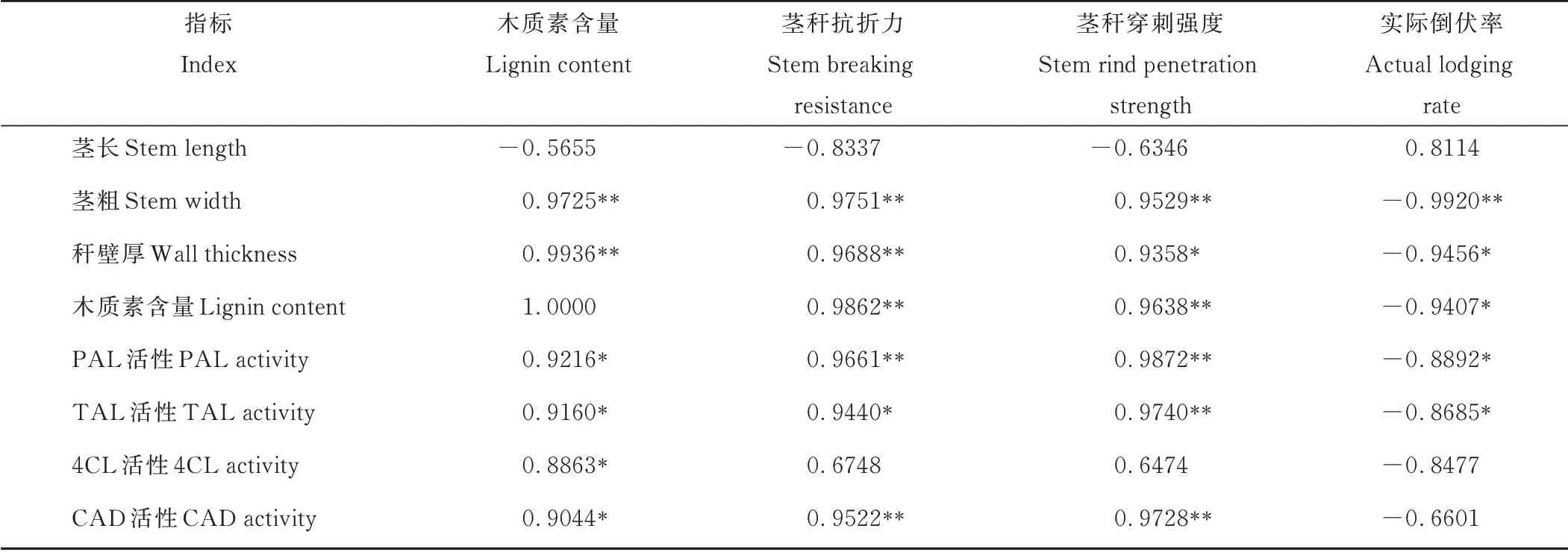
表2 茎秆木质素含量及其酶活性与抗倒伏能力的相关系数Table 2 Correlation coefficients of lignin content and related enzyme activity with lodging resistance
3 讨论
3.1 燕麦茎秆形态特征与抗倒伏的关系
作物茎秆形态特征、生理特性等因素影响其植株的抗倒伏能力[20],茎秆形态结构作为基于形态学鉴别与区分作物抗倒伏能力强弱的主要依据[21],其中茎秆质量、茎秆充实度[6,22]、茎秆重心高度,基部伸长节间长度、粗度、秆壁厚度、节间重量、抗折力及穿刺强度等与抗倒伏的关系密切[23]。研究发现燕麦茎秆矮、粗,坚韧性强,重心高度低,负荷轻,植株的抗倒性能力强[24],Pinthus[25]认为燕麦茎秆支撑的主要受力部位在基部节间,基部伸长节间短、粗的植株抗倒伏能力强。本研究表明,抗倒伏燕麦品种茎秆基部第2伸长节间短、粗、秆壁厚,茎重大,密度和充实度高,抗折力和穿刺强度大,田间实际倒伏率较低,倒伏程度轻。这可能是由于抗倒伏燕麦品种在生长发育后期,将更多的养分用于植株茎秆横向生长,使其茎秆重心高度降低,基部伸长节间变短变粗,茎秆表皮细胞壁中填充了较多的木质素等结构性化合物,从而提高了茎秆抗折力和茎秆穿刺强度,增强了茎秆机械性能。与抗倒伏品种相比,灌浆期间易倒伏品种将其更多地同化物分配到营养器官中,引发茎秆快速纵向生长,使其茎秆基部伸长节间变长,秆壁变薄,导致茎秆抗折力和穿刺强度减小,降低茎秆机械强度,田间易发生倒伏。彭佃亮[26]和Wang等[27]研究认为木质素、纤维素等结构性物质的积累,可以加固细胞壁的机械强度,能有效增强茎秆刚性,进而提高植株的抗倒伏能力。赵小红等[28]认为茎秆穿刺强度和抗折力的快速增加与木质素的积累密切相关,木质素含量提高可显著增强茎秆的机械强度,有效提高茎秆的抗倒伏能力,进而降低田间实际倒伏率[29],茎秆抗折力和穿刺强度是茎秆形态特性中决定倒伏的重要特征[30],茎秆形态结构和生理特征与抗倒伏性相关,提高茎秆基部伸长节间木质素含量是增强茎秆抗倒伏能力的关键。本研究表明,茎秆木质素含量和茎秆抗折力显著正相关(P<0.05),而与田间实际倒伏率显著负相关(P<0.05),表明茎秆中木质素含量的下降是引起燕麦茎秆抗折力和机械强度减小的重要因素。茎秆基部第2伸长节间木质素含量高的燕麦品种,茎秆抗折力大,抗倒伏能力强,田间实际倒伏率低,适当降低茎秆基部伸长节间的长度、增加秆壁厚,能有效提高燕麦茎秆的抗倒伏能力。
3.2 木质素合成相关酶活性与抗倒伏性的关系
木质素作为衡量作物抗倒伏能力强弱的一个重要生理指标,集中于木质化的机械组织和输导组织细胞壁中,能维持细胞壁稳定的物理特征和茎秆机械性能。木质素合成代谢过程比较复杂,有许多酶基因参与调控,主要包括PAL、TAL、CAD、4CL这4个酶类[31],这也是禾本科植物木质素合成过程中的重要酶类,在响应非生物胁迫中起关键作用[32]。PAL属莽草酸代谢途径中的一个限速酶,能催化苯丙氨酸脱氨产生反式肉桂酸,罗自生[33]认为PAL酶活性下降时,植株茎秆中木质素含量显著降低;当PAL活性过量表达时,植株茎秆木质素含量会明显增加。本研究表明,PAL活性与燕麦茎秆基部伸长节间木质素含量和茎秆抗折力显著相关,PAL活性升高,木质素含量会增加,与易倒伏品种相比,抗倒伏品种茎秆基部第2伸长节间PAL活性较高,PAL活性的升高对增强燕麦抗倒伏能力至关重要。TAL作为仅存在于禾本科植物木质素合成代谢过程中的关键酶,用于催化酪氨酸生成香豆酸[34],本研究中抗倒伏品种茎秆基部第2伸长节间存在一定量的TAL,TAL与PAL活性在不同倒伏类型燕麦品种间的变化趋势基本保持一致,TAL活性略低于PAL。刘晓燕等[35]认为CAD在木质素合成代谢过程中参与了最后一步的还原反应,本研究中CAD活性和木质素含量极显著(P<0.01)正相关,表明CAD活性升高会促进燕麦茎秆基部第2伸长节间木质素含量的增加,灌浆期对增强燕麦茎秆抗倒伏能力的正向调控作用相对较大。Boerjanl等[36]认为4CL参与了木质素生物合成过程的链式反应,能催化肉桂酸生成相应的CoA酯类,相关分析表明,4CL活性与燕麦茎秆基部第2伸长节间抗折力、穿刺强度正相关但不显著,这可能与4CL参与了催化羟基肉桂酸及其木质素合成过程中其他中间衍生产物的代谢活动,与辅酶A酯化合物的形成有关[37]。本研究中同一燕麦品种茎秆基部第2伸长节间PAL活性最高,4CL活性较低,CAD活性与PAL活性程度相当,抗倒伏品种茎秆基部第2伸长节间PAL、TAL、CAD、4CL活性显著高于易倒伏品种,4种酶活性在蒙燕1号表达量均较高,而在青引2号中表达量最低,这与陈晓光等[9]和汪灿等[13]的相关研究结果保持一致,田间实际倒伏率低的品种,茎秆中木质素含量及其合成酶活性、茎秆抗折力均大于易倒伏品种。茎秆基部伸长节间木质素含量及合成酶活性的高低显著影响燕麦茎秆抗折力和抗倒伏性能,推测PAL和CAD活性的增强更有利于提高燕麦茎秆基部第2伸长节间木质素含量,提高燕麦茎秆的抗倒伏能力。
4 结论
燕麦茎秆基部第2伸长节间形态和其生理特性与抗倒伏能力有关,抗倒伏品种相比易倒伏品种表现为主茎基部第2伸长节间短,茎秆粗,秆壁厚,内腔体积小,茎秆密度和充实度高,茎秆抗折力和穿刺强度大,茎秆质量优,倒伏程度轻,田间实际倒伏率低。茎秆木质素含量及其合成酶(TAL、PAL、CAD和4CL)活性大,能有效促进茎秆基部伸长节间木质素的积累,增大茎秆抗折力,提高茎秆机械强度和支撑弹性,从而增强燕麦植株的抗倒伏能力。
