长穗偃麦草(Thinopyrum ponticum)幼苗对盐旱胁迫的生理响应*
2022-11-16封晓辉吴玉洁李劲松刘小京
张 睿,封晓辉,吴玉洁,孙 琦,李 静,李劲松,刘小京**
(1.河北省土壤生态学重点实验室/中国科学院盐碱地资源高效利用工程实验室/中国科学院遗传与发育生物学研究所农业资源研究中心 石家庄 050022;2.中国科学院大学 北京 100049;3.唐山曹妃甸滨海公路开发建设有限公司 曹妃甸 063200)
干旱和土壤盐渍化是制约农业发展的主要因素[1]。尤其在盐渍区域,由于蒸发量大、降水量少导致土壤积盐,植物在盐渍区不仅面对盐胁迫,也受到干旱胁迫,两者往往同时发生,会产生盐与干旱的双重胁迫。盐胁迫和干旱胁迫均会使土壤溶液水势下降,造成植物细胞失水而导致植株吸水困难,最终对植物生长产生不利影响[2-3]。滨海盐渍区水资源缺乏且含盐量高,春季干旱少雨,地表蒸发快,土壤表层返盐重[4],生长在这一地区的作物易遭受盐和干旱的双重胁迫,因此,在盐碱地中的植物需具备较强的耐盐和耐旱能力。
目前,关于植物对干旱、盐胁迫响应方面的研究主要集中在小麦(Triticum aestivum)[5]、沙芦草(Agropyron mongolicum)[6]、蓝稷(Panicum antidotale)[7]、柽柳(Tamarix chinensis)[8]、燕麦(Avena sativa)[9]等植物。植物对盐、旱交互胁迫的响应主要体现在渗透调节、离子平衡以及植物体内的抗氧化酶活性变化[10]。盐旱交互胁迫会破坏植物体内的氧化还原平衡,导致活性氧(reactive oxygen species,ROS)的过度生成,从而引起二次氧化胁迫[11]。吕廷良等[12]对紫荆幼苗(Cercis chinensis)在盐、旱及其交互胁迫下净光合速率及其叶绿素含量变化的研究表明,低浓度的盐分胁迫、低强度的干旱胁迫以及低强度的交互胁迫在短时间内均可增强紫荆的光合能力,增加植株对胁迫的抵抗能力。于振群等[13]以皂角(Gleditsia sinensis)为材料,研究了盐旱交互胁迫对幼苗膜脂过氧化及保护酶活性的影响,结果表明适度的水分胁迫能够缓解盐胁迫,但在严重的干旱条件下,盐、旱交互胁迫的适应性降低,超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)和过氧化氢酶(catalase,CAT)活性明显下降,不足以清除体内的自由基,导致脂质过氧化和膜系统受到损伤。翁亚伟等[5]对小麦幼苗(Triticum aestivum)进行盐旱复合胁迫的试验,结果表明小麦幼苗在复合胁迫下表现出一定的交互胁迫适应性,即通过降低Na+/K+缓解离子毒害,从而保持相对高效的光合速率。
长穗偃麦草(Thinopyrum ponticum)是小麦族禾本科偃麦草属(Thinopyrum)的多年生草本植物。其根茎发达,多花多实,侵占能力强,生长速度快,草质柔软,茎叶蛋白含量高,是我国重要的牧草物种,也是防风固沙、盐碱地改良、天然草地改良的理想植物[14-15]。种植偃麦草具有改良盐碱土、抑制杂草、为野生动物提供栖息地点等效益[16-17],同时偃麦草属植物也是小麦遗传改良的野生种质资源库,具有较高的经济价值和种质资源价值。长穗偃麦草是优良的牧草品种,已在国外大面积种植。李振声通过小麦和长穗偃麦草远缘杂交的方法,育成了小偃系列小麦品种;同时,依据农艺性状,选出了远缘杂交的第一代7 个可作为牧草品种的株系[18]。前人的研究表明,在耐盐性植物中,长穗偃麦草比大多数单子叶植物[如水稻(Oryza sativa)、小麦和大麦(Hordeum vulgare)]和一些双子叶植物[如苜蓿(Medicago Sativa)]具有更强的耐盐性[19]。2020年,李振声提出在环渤海地区的66.7 万 hm2盐碱地上种植耐盐牧草的“滨海草带”。有学者通过对长穗偃麦草在不同盐分条件下的产量、农艺性状和生理生化指标研究发现,长穗偃麦草在滨海盐渍化土上具有较高的适应能力,且对这一地区的盐渍化土壤具有较好的控制和改良作用[20]。
目前,有关长穗偃麦草对干旱和盐分响应的研究多集中在干旱或盐的单一因素胁迫[21-22],而以盐旱交互胁迫对长穗偃麦草生长的影响尚少见报道。因此,本研究以长穗偃麦草为材料,利用土壤盆栽试验,研究干旱、盐及其互作对长穗偃麦草的生长及生理特性的影响,明确其抗旱耐盐特征,以期为长穗偃麦草在盐碱地中的栽培提供理论依据。
1 材料与方法
1.1 材料培养与胁迫处理
本试验在中国科学院南皮生态农业试验站防雨棚中进行。供试材料为长穗偃麦草,由中国科学院遗传与发育生物学研究所李振声课题组提供。种子经消毒,然后播种于育苗钵中,待生长到15 cm 高时,选取生长良好,长势基本一致的实生苗,移栽至盛有2.3 kg 风干菜园土的塑料花盆中(长×宽×高为17 cm×16 cm×13 cm),每盆移栽2 株,然后灌水,缓苗15 d后进行试验处理。花盆底部用托盘承接,收集渗漏水及时返还花盆中,以确保盆内盐总量。盐分处理按施入土壤NaCl 的量设0 g·kg—1、4 g·kg—1、8 g·kg—1和12 g·kg—14 个水平,干旱处理按占土壤田间持水量的百分比设75%~85%(正常供水)、55%~65%(中度干旱)、35%~45%(重度干旱)3 个水平,共12 个处理,以不施用NaCl 和正常供水的处理为对照(CK),每个处理重复8 次。试验期间白天温度25~30 ℃,晚上15~20 ℃,空气湿度40%~60%。土壤含盐量的控制是通过灌入不同梯度的NaCl 溶液来实现,为防止短时间内盐对植株生长造成的冲击,将施入土壤的NaCl 溶液从4 g·kg—1起递增,每天递增4 g·kg—1,直至达预定含盐量,对照处理浇灌相同体积的清水。达预定的土壤含盐量后开始控制水分,采用称重法控制水分梯度,根据不同水分梯度对应的土壤含水量控制水分条件,5 d 达预定水分条件,试验期间定时称盆补水,称重间隔为2 d,为了减少蒸发,达预定土壤水分和盐分时用塑料薄膜覆盖整个花盆盆面。每周进行一次随机区组排列,避免边行效应。处理28 d后,开始对植株进行各项指标的测定。
1.2 测定指标
生物量测定:将地上部与根系分开收获,分别装入信封中,在烘箱105 ℃杀青30 min,85 ℃下烘至恒重,用高精度天平分别称得干重。
叶片SPAD 值和气体交换参数测定:采用SPAD-502 型手持式叶绿素仪(日本 Minolta 公司)测定叶片的叶绿素SPAD 值。测量时,在每个叶片的主脉两侧中部测3 个点,取平均值作为该叶片的SPAD 值。取每个重复中3 个叶片的SPAD 值的平均值作为该重复叶片的SPAD 值。采用LI-6400 光合仪(LI-COR,Inc.USA)于晴天8:30—11:00 测定长穗偃麦草主茎倒二叶的净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rate,Tr)、气孔导度(stomatal conductance,Gs)和胞间二氧化碳浓度(intercellular carbon dioxide concentration,Ci),设定的环境条件为光强500 μmol·m—2·s—1、温度20 ℃,相对湿度30%~40%。
丙二醛(MDA)含量和保护酶活性测定:试验结束时,分别取各处理新鲜的叶片,每个处理取0.8 g,液氮速冻置于—80 ℃保存,测定其MDA 含量和保护酶活性。采用硫代巴比妥酸法测定丙二醛的含量,采用分光光度计测定在532 nm、600 nm 以及450 nm下的吸光度值,计算MDA 含量。超氧化物歧化酶(SOD)活性以氮蓝四唑(NBT)法[23]测定,以抑制氮蓝四唑光氧化还原的50%酶量为1 个活性单位,用分光光度计测定560 nm 处的吸光值,计算SOD 的活性。过氧化物酶(POD)活性以愈创木酚法测定[23],按每分钟A470变化0.01 为1 个酶活性单位,采用分光光度计测定其在470 nm 处的吸光值变化,计算POD 活性。过氧化氢酶(CAT)活性的测定采用可见光法[23],以吸光度每分钟变化0.01 为1 个酶活性单位,用分光光度计测定其在240 nm 处的吸光值变化,计算CAT 活性。
无机离子含量的测定:取植株地上部和地下部分用自来水和去离子水冲干净后于105 ℃杀青30 min后85 ℃烘干至恒重,磨碎、称重后置于1 mmol·L—1HCl 溶液中震荡浸提2 h,用蒸馏水定容后再使用原子吸收分光光度仪(WYX-420C,JASCO Corporation,Japan)测定Na+、K+含量,并计算K+/Na+。
1.3 数据处理
数据采用Excel 和SPSS 26.0 软件进行处理分析,并使用邓肯法对数据进行多重比较(P<0.05),用SPSS 对各项指标进行Pearson 相关性分析,得到相关系数矩阵,Origin 9.1 软件进行制图。
2 结果与分析
2.1 盐旱互作对长穗偃麦草幼苗生长的影响
表1 结果表明,干旱或盐胁迫极显著影响长穗偃麦草地上部干重和根干重(P<0.01),干旱胁迫对根/冠比无显著影响,而盐分显著影响根/冠比(P<0.05);干旱和盐分互作对地上部干重无显著影响,但极显著影响根干重和根/冠比(P<0.01)。长穗偃麦草地上部干重和根干重随土壤含水量降低或土壤含盐量增加而下降。单一干旱胁迫降低根/冠比,同一盐分水平在不同干旱胁迫下的根/冠比差异不显著,但同一干旱处理下根/冠比随土壤含盐量增加而增加。在盐旱互作下,地上部干重随土壤含盐量增加和土壤含水量降低进一步下降:4 g·kg—1、8 g·kg—1和12 g·kg—1NaCl 处理的地上部干重与同一干旱处理下的无NaCl 处理相比,在常规灌水下分别降低24.3%、38.3%和56.0%,在中度干旱处理下分别降低24.1%、48.2%和55.9%,在重度干旱处理下分别降低27.9%、56.6%和60.2%;根干重则随土壤含水量的降低,盐分胁迫导致的根干重下降趋势减缓,4 g·kg—1、8 g·kg—1和12 g·kg—1NaCl 处理的根干重与同一干旱处理下的无NaCl 处理相比,在常规灌水下分别降低44.8%、49.8%和59.3%,而在中度干旱处理下分别降低13.7%、36.7%和33.8%,在重度干旱处理下分别降低14.4%、38.1%和50.8%;根/冠比则随土壤含水量的降低,盐分胁迫导致的根/冠比增加效应更强,4 g·kg—1、8 g·kg—1和12 g·kg—1NaCl 处理的根/冠比与同一干旱处理下的无NaCl 处理相比,常规灌水下分别降低29.3%、22.0%和9.8%,而在中度干旱处理下分别增加了10.3%、20.7%和44.8%,在重度干旱处理下分别增加16.1%、38.7%和19.4%。如果按地上部生物量比对照降低一半时的处理为临界处理,不同旱盐处理的临界值不同,在正常供水(75%~85%)条件下,当土壤含盐量达12 g·kg—1时,地上部干重是对照干重的44.0%;在中度干旱(55%~65%)胁迫条件下,当土壤含盐量达8 g·kg—1时,地上部干重是对照干重的46.7%;在重度干旱(35%~45%)胁迫条件下,当土壤含盐量达4 g·kg—1时,地上部干重是对照干重的51.6%;即使在重度干旱胁迫下,当土壤含盐量达12 g·kg—1时仍能存活,这可能与其具有发达的根系和较高的根冠比有关。
2.2 盐旱互作对长穗偃麦草幼苗SPAD 值及光合参数的影响
双因素方差分析结果表明(表2),盐分、干旱及其互作均极显著影响叶片叶绿素含量、气孔导度、胞间CO2浓度和蒸腾速率(P<0.01)。盐分极显著影响叶片的净光合速率,但干旱及其与盐分互作则影响不显著。

表2 盐旱互作对长穗偃麦草叶片SPAD 值和光合参数影响的双因素方差分析Table 2 Two-way ANOVA of effects of salt,drought and their interactions on leaf SPAD value and photosynthetic parameters of Thinopyrum ponticum
由图1 可知,单独干旱胁迫显著降低了长穗偃麦草幼苗叶片叶绿素含量(P<0.05),中度(55%~65%)和重度(35%~45%)干旱胁迫的叶片SPAD 值与对照相比分别降低5.7%和11.9%。正常供水条件(75%~85%)下,长穗偃麦草的叶绿素SPAD 值随土壤含盐量的升高呈显著下降趋势,4 g·kg—1、8 g·kg—1和12 g·kg—1盐处理的叶片SPAD 值与对照相比分别降低7.1%、13.1%和19.5%。在中度和重度干旱胁迫下,轻度盐分(4 g·kg—1)胁迫能够提高叶绿素含量,但过高的盐分会降低叶绿素的含量。同正常供水处理相比,盐分胁迫能使长穗偃麦草叶片叶绿素含量在中度干旱胁迫下保持较高的水平。
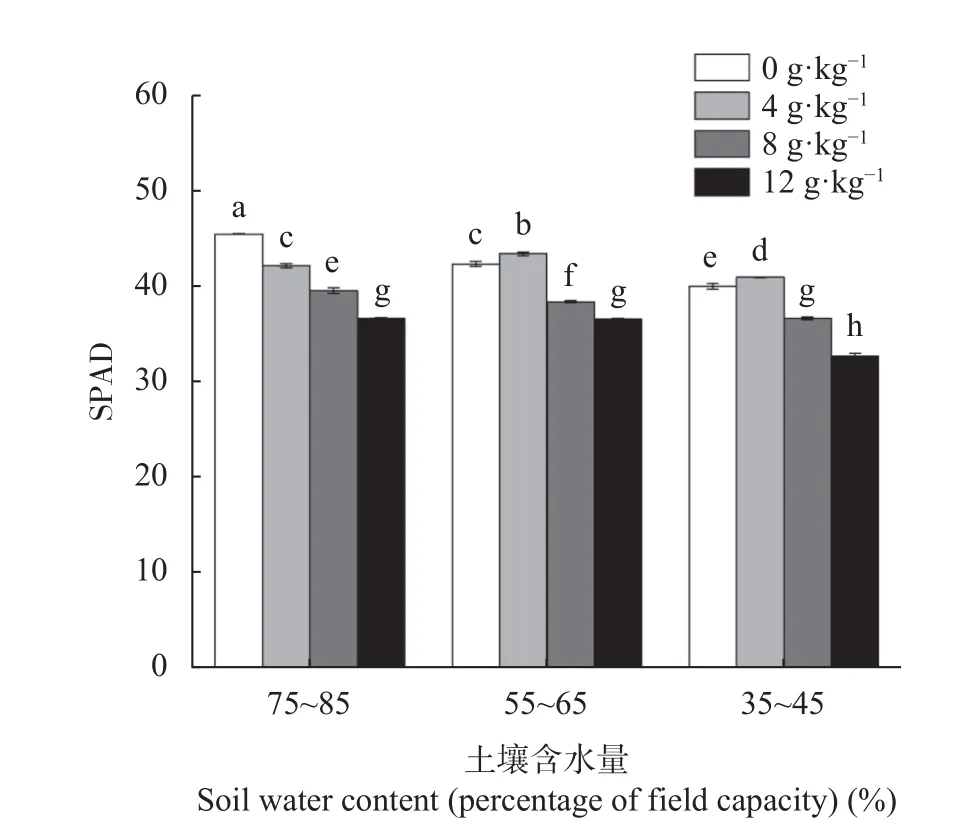
图1 盐旱互作对长穗偃麦草叶片叶绿素SPAD 值的影响Fig.1 Interactive effect of salt and drought on the leaf chlorophyll SPAD value of Thinopyrum ponticum
由图2 可知,单独干旱胁迫使长穗偃麦草叶片的净光合速率、蒸腾速率、气孔导度和胞间CO2浓度均低于对照,重度干旱胁迫下降幅最大,与对照相比分别下降23.5%、38.0%、50.2%和58.4%。正常供水条件下,盐胁迫显著降低了长穗偃麦草叶片净光合速率、蒸腾速率和气孔导度,且在土壤含盐量为12 g·kg—1时降幅达最大,与对照相比分别降低72.3%、74.2%、81.5%;而胞间CO2浓度呈先降低后升高的趋势,但均低于对照。中度干旱条件下,4 g·kg—1NaCl 处理与无NaCl 处理相比,叶片的净光合速率、蒸腾速率、气孔导度、胞间CO2浓度均有增加趋势,但差异不显著,过高的盐分显著降低了叶片净光合速率、蒸腾速率、气孔导度,增加了叶片胞间CO2浓度。在重度干旱胁迫条件下,叶片净光合速率、蒸腾速率、气孔导度随土壤含盐量的增加而显著下降,胞间CO2浓度呈显著增加趋势。
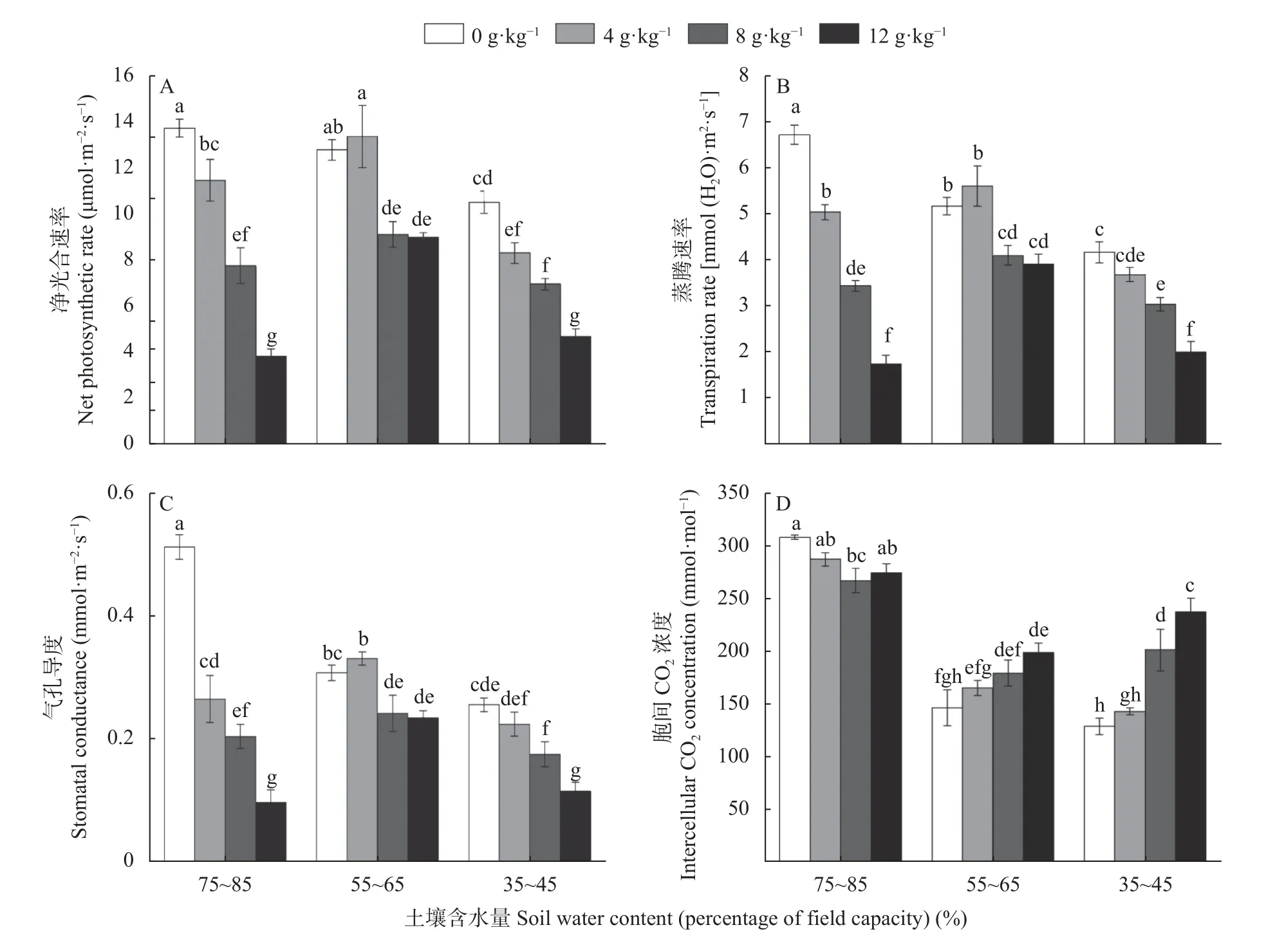
图2 盐旱互作对长穗偃麦草叶片光合参数的影响Fig.2 Interactive effects of salt and drought on leaf photosynthetic parameters of Thinopyrum ponticum
2.3 盐旱互作对长穗偃麦草幼苗叶片保护酶活性的影响
双因素方差分析结果表明(表3),盐分显著影响叶片SOD、POD、CAT 的活性和MDA 含量;干旱和旱盐互作显著影响叶片POD、CAT 的活性和MDA 含量,而对SOD 活性影响不显著。
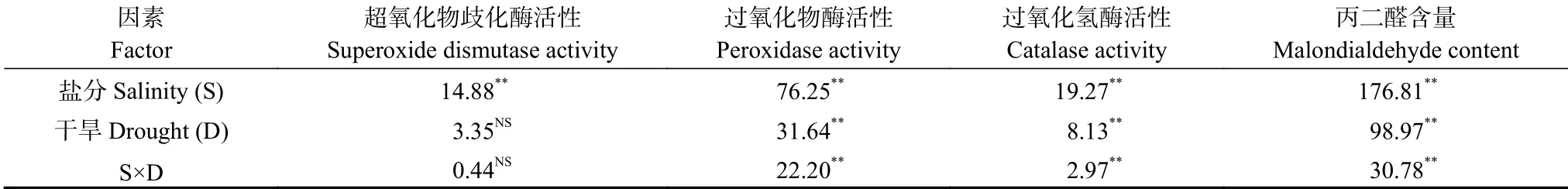
表3 盐旱互作对长穗偃麦草叶片保护酶活性和丙二醛含量影响的双因素方差分析Table 3 Two-way ANOVA of effects of salt,drought and their interactions on leaf protective enzymes activities and malondialdehyde content of Thinopyrum ponticum
植物在面对逆境时,体内会产生大量活性氧,而SOD、POD 和CAT 是植物体内重要的活性氧清除酶[24]。干旱处理有提高叶片SOD 活性的趋势,但差异不显著;同一干旱处理下,4 g·kg—1和8 g·kg—1NaCl处理提高了叶片SOD 活性,过高的盐分(12 g·kg—1)降低了SOD 活性(图3A)。盐旱互作下,叶片POD活性随干旱和盐胁迫加重显著提高,但过高的盐分(12 g·kg—1)降低了叶片POD 活性(图3B)。说明适度盐旱互作下可诱导或激活SOD 和POD 的活性,使长穗偃麦草体内自由基的产生和清除处于平衡状态,但当胁迫超过一定程度时就会打破这一平衡,从而抑制了长穗偃麦草的生长。
CAT 是植物体内重要的过氧化氢清除酶。单独干旱胁迫提高了长穗偃麦草叶片CAT 活性,与对照相比,中度和重度干旱胁迫下叶片CAT 活性分别增加16.3%和22.3%,但差异不显著;在正常供水条件下,长穗偃麦草叶片CAT 活性随土壤含盐量的增加呈现先升高后降低的趋势,在土壤含盐量8 g·kg—1处理下叶片CAT 活性比对照增加16.7%,而在土壤含盐量12 g·kg—1处理叶片CAT 活性比对照降低。在中度干旱胁迫与不同盐分互作下,CAT 活性随含盐量增加呈先增加后降低趋势,在4 g·kg—1土壤含盐量下达峰值,增幅为62.7%;在重度干旱胁迫和不同盐分互作下,与对照相比,随胁迫程度的增加,CAT 活性同样呈先升高后降低的趋势,在土壤含盐量为4 g·kg—1和8 g·kg—1时增幅分别为22.7%和26.4%,但在12 g·kg—1土壤含盐量则下降12.5%(图3C)。以上结果表明,干旱和低浓度盐分可使CAT 被激活,使得长穗偃麦草渗透调节能力增强,能够耐受一定程度的干旱和盐胁迫。
MDA 是植物细胞中膜脂过氧化的重要产物,可与细胞膜上的蛋白质、酶等生物大分子结合,交联使之失活,从而使得生物膜结构和功能遭到破坏。由图3D 可知,在无NaCl 处理条件下,与对照相比,长穗偃麦草叶片MDA 含量随土壤含水量的降低呈先降低后升高的趋势,差异显著;在正常供水条件下,只有12 g·kg—1土壤含盐量处理导致叶片MDA 含量显著升高;在中度干旱胁迫下,4 g·kg—1NaCl 处理显著降低了叶片MDA 的含量,之后随土壤含盐量升高叶片MDA 含量显著升高;但在重度干旱胁迫下,叶片MDA 含量随土壤含盐量的升高而升高。说明在盐旱互作下,随胁迫程度加深,长穗偃麦草叶片保护酶活性下降,细胞内自由基的平衡被打破,从而引发了细胞的膜脂过氧化作用,植株生长受到抑制。

图3 盐旱互作下长穗偃麦草叶片超氧化物歧化酶(SOD,a)、过氧化物酶(POD,b)、过氧化氢酶(CAT,c)活性和丙二醛含量(MDA,d)的变化Fig.3 Leaf superoxide dismutase(a),peroxidase(b)and catalase(c)activities and the content of malondialdehyde(d)of Thinopyrum ponticum under the interaction of salt and drought
2.4 盐旱互作对长穗偃麦草幼苗离子含量的影响
双因素方差分析结果表明(表4),盐分或干旱极显著(P<0.01)影响长穗偃麦草地上部和根系中Na+、K+含量和K+/Na+比。盐旱互作极显著(P<0.01)影响根部Na+含量和地上部K+/Na+比,但对地上部Na+、K+含量和根部K+含量和K+/Na+比影响不显著。长穗偃麦草地上部Na+、K+含量和K+/Na+比高于根部,平均分别是根系的3.6 倍、6.9 倍和1.9 倍。在单独干旱胁迫条件下,根系和地上部Na+含量随土壤含水量的降低而显著升高,K+含量有下降趋势且在重度干旱胁迫下显著下降,K+/Na+比显著下降但依然维持在较高的水平。在充分供水条件下,地上部Na+含量随土壤含盐量的升高而显著升高,添加盐分显著降低地上部K+含量且随土壤盐分增加有降低趋势但差异不显著,K+/Na+比显著下降;土壤中添加盐分显著增加根系Na+含量且随土壤含盐量增加有增加趋势但差异不显著,根部K+含量相对稳定,但过高的土壤盐分显著降低根部K+含量,盐分添加显著降低根部K+/Na+比但随土壤含盐量增加保持稳定。在盐旱互作时,各指标变化同充分供水条件下趋势一致,但根部Na+含量随土壤含水量降低和土壤盐分含量增加进一步显著增加,地上部K+/Na+比则进一步显著下降但依然维持在较高水平。通过离子变化分析表明,长穗偃麦草植株在盐旱胁迫下主要通过积累Na+维持细胞膨压,缓解渗透胁迫对植株产生的不利影响。
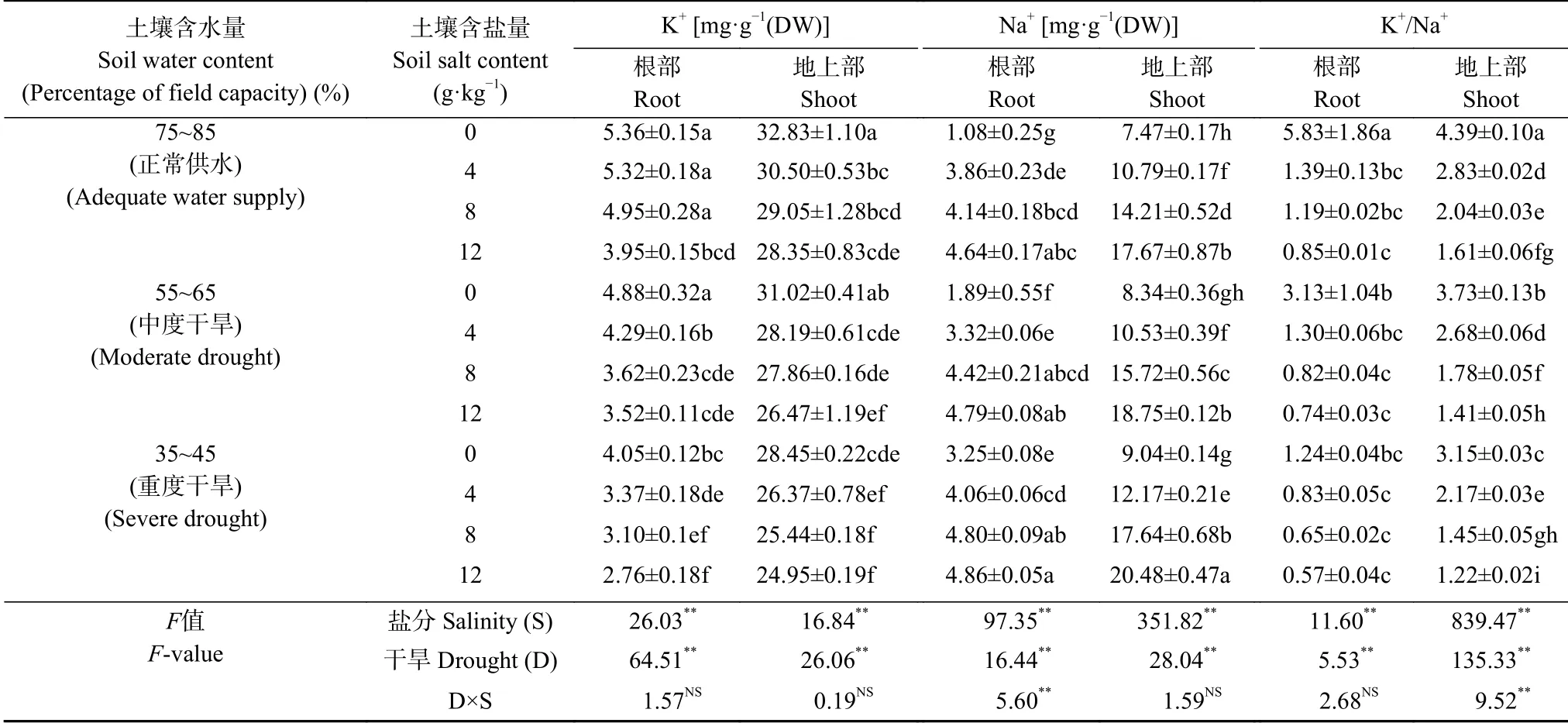
表4 盐旱互作对长穗偃麦草幼苗叶片和根系Na+、K+含量和K+/Na+的影响Table 4 Effect of salt and drought interaction on Na+,K+ contents and K+/Na+ ratio in leaves and roots of the seedlings of Thinopyrum ponticum
2.5 盐旱胁迫下长穗偃麦草各指标的相关性分析
相关分析结果表明(表5),净光合速率与蒸腾速率、气孔导度和SPAD 值呈极显著正相关,与地上部和根部K+含量呈显著正相关,与地上部和根部Na+含量、MDA 呈极显著负相关;蒸腾速率和气孔导度与SPAD、地上部和根部K+含量呈极显著或显著正相关,与地上部和根部Na+含量、MDA 呈极显著负相关;SPAD 值与地上部和根部Na+含量、MDA 呈极显著负相关,与地上部和根部K+含量呈极显著正相关;SOD 与POD、CAT 呈极显著正相关;POD 与CAT 呈显著正相关;CAT 与MDA 呈极显著负相关;MDA 与地上部和根部Na+含量呈极显著或显著正相关,与地上部和根部K+含量呈显著负相关;地上部Na+含量与地上部和根部K+含量呈极显著负相关,与根部Na+含量呈极显著正相关;地上部K+含量与根部K+含量呈极显著正相关,与根部Na+含量呈极显著负相关;根部Na+含量与根部K+含量呈极显著负相关。

表5 盐旱胁迫下长穗偃麦草生理指标之间的相关性Table 5 Correlation of physiological indexes of Thinopyrum ponticum under salt and drought stress
3 讨论与结论
生物量是植物对胁迫响应的综合体现,也是目前评价植物耐盐、抗旱能力最直接的指标[25]。在干旱和盐分胁迫条件下,植物体内发生一系列复杂的生理生化反应以应对逆境胁迫。陈成升等[26]对小麦盐旱交互胁迫的研究发现,在单一干旱或盐分胁迫下随着干旱和盐胁迫程度的升高,小麦幼苗的干物质积累量明显下降;干旱胁迫下,适量盐分的添加可使小麦幼苗干物质含量增加。而本研究中,干旱胁迫和盐胁迫均可使长穗偃麦草的生长受到抑制,与上述研究结果相一致;但盐旱互作下,随着土壤含水量的降低,盐分导致长穗偃麦草的根/冠比增加效应更强。
光合作用是植物生长发育和面对逆境的一个基本代谢过程,光合作用的许多代谢产物参与植物体内的渗透调节和细胞内环境稳态的维持。叶绿素相对含量(SPAD)是衡量叶绿素含量的一个重要指标[27]。本研究发现干旱和盐胁迫均可使长穗偃麦草叶绿素含量显著下降,且随胁迫程度的加深,下降幅度增加。盐旱互作下,适度的盐分可使干旱胁迫得到一定程度的缓解,过量盐分与不同土壤水分条件互作下,缓解作用消失,说明盐胁迫对长穗偃麦草的影响大于干旱胁迫,因为植物面临盐胁迫时不仅会产生渗透胁迫,还会产生离子胁迫,而在面临干旱胁迫时主要产生渗透胁迫[28]。刘建新等[9]的研究表明,适量的土壤盐分可缓解水分胁迫对燕麦生长的影响,与本研究结果相一致。
引起光合作用下降的主要原因一是气孔导度的下降,CO2的供应受到限制,即气孔限制因素;二是植物叶肉细胞光合活性下降,如Rubisco 羧化活性下降,引起的叶肉细胞对CO2同化能力降低的非气孔限制性因素[29]。本研究发现,在单独盐胁迫以及盐旱交互胁迫下,长穗偃麦草的光合速率、蒸腾速率、气孔导度均呈不断下降趋势,胞间CO2浓度在盐胁迫下呈先降低后升高趋势,在交互胁迫下呈不断增加趋势,说明高盐和盐旱复合胁迫下光合作用受到抑制是由于叶肉细胞对CO2利用能力下降的非气孔限制性因素引起的,可能是由于高盐胁迫和盐旱复合胁迫所诱导的渗透胁迫和离子毒害使PSⅡ氧化侧的放氧复合物的功能得到损害,造成了其向PSⅡ反应中心的电子供应数量减少。单独干旱胁迫下,随胁迫程度增加,长穗偃麦草的各光合参数均呈下降趋势,且单独盐胁迫下各光合参数与对照相比的降幅大于干旱胁迫,说明盐胁迫对长穗偃麦草生长的不利影响更大。已有的研究表明盐胁迫比等渗干旱胁迫对植物生长的危害更大[30],与本试验的研究结果一致。相关性分析表明净光合速率与蒸腾速率、气孔导度、SPAD 呈极显著正相关,与胞间CO2浓度呈负相关关系,说明光合参数蒸腾速率、气孔导度的降低是长穗偃麦草对盐旱及双重胁迫保持适应性的重要机制,另外叶绿素SPAD 值的降低也是长穗偃麦草净光合速率下降的影响因素之一。郭书奎等[31]对玉米(Zea mays)叶片的研究发现盐胁迫时间较短时,光合作用受到抑制以气孔限制因素为主,时间较长时则以非气孔限制性因素为主;秦景等[32]对沙棘(Hippophae rhamnoides)幼苗叶片的研究认为,限制光合作用的气孔因素和非气孔因素是随胁迫浓度动态变化的,盐浓度越高,由气孔限制转为非气孔限制的时间越早,这与我们的研究相一致。
MDA 是膜脂质过氧化的重要产物,也是评估细胞膜受到伤害程度的重要指标。本研究结果表明:在单独盐胁迫或干旱胁迫下,MDA 含量随胁迫程度的加深均呈先降低后增加的趋势。在适度盐旱互作下,MDA 含量呈先降低后升高的趋势;在重度盐旱互作下,MDA 含量却持续增加。在适度的盐旱互作下,与MDA 含量变化对应的过氧化物酶呈先升高后降低趋势,说明此时抗氧化酶活性增加参与体内自由基的清除,减轻细胞膜脂过氧化程度,增强长穗偃麦草抵抗逆境的能力。而余淑艳等[6]通过对沙芦草的研究发现,盐旱互作下,MDA 含量显著增加。但朱金方等[8]对柽柳幼苗的研究表明,适度的盐旱互作能够降低植株MDA 含量,增强植株对逆境的抵抗能力,与本研究结果相一致。
植物体内抗氧化酶活性的应激性变化是反映植物受逆境胁迫程度的重要指标之一。SOD、POD 和CAT 是植物中的主要抗氧化酶,主要作用为清除自由基,抗脂质过氧化,减少膜损伤等。SOD 可催化O2—生成H2O2,而POD 和CAT 又可将H2O2降解为H[33]。逆境胁迫下,植物体内抗氧化酶活性发生变化,自由基产生和清除的平衡状态被打破,抗氧化酶活性的变化因植物种类、胁迫种类及胁迫时间的不同而呈现不同的变化[34]。本试验的结果显示,长穗偃麦草幼苗叶片的SOD、POD、CAT 活性在盐胁迫和盐旱互作时呈先升高后降低的趋势,在干旱胁迫下均呈现升高趋势。说明长穗偃麦草在面临胁迫时的耐受能力是有一定的范围的,盐旱互作下,植物体内活性氧增加,为清除自由基对细胞膜系统的伤害,长穗偃麦草叶片的抗氧化酶活性逐渐升高,但抗氧化酶系统的调节能力也是有限的,当胁迫超出一定范围后,体内的自由基过多,超出抗氧化酶系统的清除能力,此时,抗氧化酶活性下降,植物生长受到了抑制。石新建等[24]通过对花花柴(Karelinia capsica)幼苗的盐旱胁迫的研究表明,在盐旱胁迫下,随着胁迫程度的增加植株体内抗氧化物酶(SOD、POD、CAT)活性呈先上升后下降的趋势。王利界等[35]通过对灰胡杨(Populus pruinosa)在盐旱交互胁迫下的研究发现,盐胁迫、盐旱交互胁迫下,随着胁迫程度的加重,SOD、POD、CAT 活性均呈先上升后下降趋势,单一干旱胁迫下SOD、POD、CAT 活性显著增加。刘建新等[36]对燕麦的研究表明,随盐分、干旱胁迫程度的提高,SOD、POD 活性呈先升高后降低趋势,CAT 活性增加,与本试验的研究结果一致。
相关性分析可知,SOD、POD 和CAT 活性与MDA 含量呈显著负相关关系,且3 种抗氧化酶活性之间则表现为显著正相关关系,可推测,盐旱互作下长穗偃麦草抗氧化酶系统活性的变化是引起长穗偃麦草膜脂过氧化程度变化的重要影响因素;在面临逆境胁迫时长穗偃麦草体内的3 种抗氧化物酶可能产生协同作用,形成较为完整的抗氧化物酶系统,进而抵御外界逆境对植物体内的伤害。
随着胁迫的加剧,植物体内离子平衡调节会被打破,产生离子毒害,从而影响了植物的正常生长。当植物体内Na+过高时,K+向外渗透,Na+/K+值增大。当Na+/K+比值增大到超出植物体正常生长所能承受的临界值时,植物受到伤害[37]。胁迫发生时,植物一般通过Na+的外排和Na+的区隔化来降低Na+的浓度。一般通过质膜Na+/H+逆向转运蛋白将Na+排出细胞来实现Na+的外排,而通过NHX1 蛋白将Na+区隔化入液泡中以消除Na+的毒害来实现Na+的区隔化。K+是参与蛋白质合成、糖酵解酶和光合作用的重要离子,在胁迫条件下可维持细胞膨压,当Na+含量过高时,与K+竞争,影响K+发挥正常的生理作用[38]。翁亚伟等[5]的研究表明,盐旱互作下的Na+、K+低于单独的盐胁迫,缓解了离子毒害,体现了植物在交互胁迫下的适应性。本研究中,在干旱或盐分胁迫下,根部和地上部Na+含量升高,K+含量降低,但依然维持在较高水平。适度盐分(4 g·kg—1NaCl)与中度(55%~65%田间持水量)干旱程度的互作下,与单一的4 g·kg—1NaCl胁迫相比,地上部和根系的K+、Na+含量均降低,但其与重度干旱(35%~45%田间持水量)胁迫互作时,地上部和根系的Na+含量增加、K+含量降低。说明长穗偃麦草在干旱或盐分胁迫下具有吸收积累Na+的特征,通过吸收Na+来维持细胞膨压抵抗逆境,同时地上部高的K+积累有利于调控离子平衡。K+含量的增加可参与提高植物细胞的渗透调节能力,稳定的K+吸收和分布可平衡Na+积累的毒性作用并控制胁迫期间植物体内的离子稳态。相关性分析表明,盐胁迫下长穗偃麦草的净光合速率与地上地下部K+具有显著的正相关性,与地上地下部Na+呈极显著负相关关系,说明盐旱胁迫下所产生的渗透胁迫对长穗偃麦草叶片的光合作用造成了影响。
综上所述,长穗偃麦草能够在适度的盐胁迫(4 g·kg—1NaCl、8 g·kg—1NaCl)、中度干旱胁迫(55%~65%田间持水量)以及适度的盐旱互作下(4 g·kg—1NaCl,55%~65%田间持水量)生长,且在重度干旱(35%~45%田间持水量)和高盐(12 g·kg—1NaCl)条件下能够存活,这与其在胁迫下Na+的吸收和其本身高K+特征以及抗氧化酶活性上升有关。虽然长穗偃麦草能够在重度干旱和盐胁迫下存活,但其生长随着干旱或盐分胁迫程度增加而受到显著抑制,盐旱互作下胁迫进一步加重,在8~12 g·kg—1NaCl 和重度干旱互作下,长穗偃麦草生长受到显著影响。长穗偃麦草在干旱胁迫下Na+的积累及其在抗逆中的调控机制有待进一步加强。长穗偃麦草幼苗在生长过程中表现出较高的抗盐耐旱性能,可用于盐渍区推广种植,但由于旱或盐显著抑制其生长,要获得较高的生物量,需选择适宜的低盐和适度干旱土壤。
